
Borneol Depresses P-Glycoprotein Function by a NF-κB Signaling Mediated Mechanism in a Blood Brain Barrier in Vitro Model

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction

2. Results
2.1. Cell Characterization and Establishment of an in Vitro BBB Model

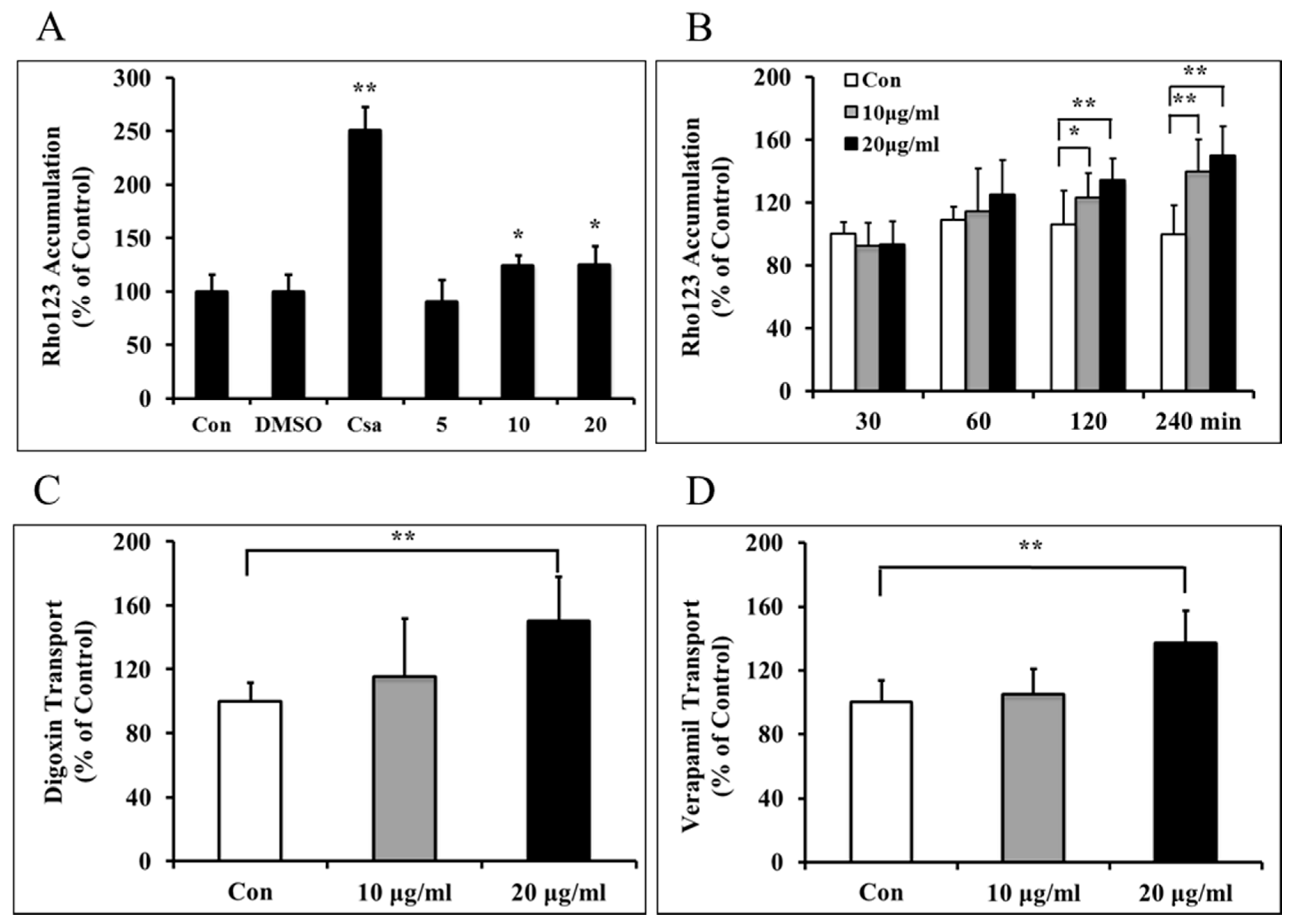
2.2. Borneol Down-Regulated P-gp Efflux Function

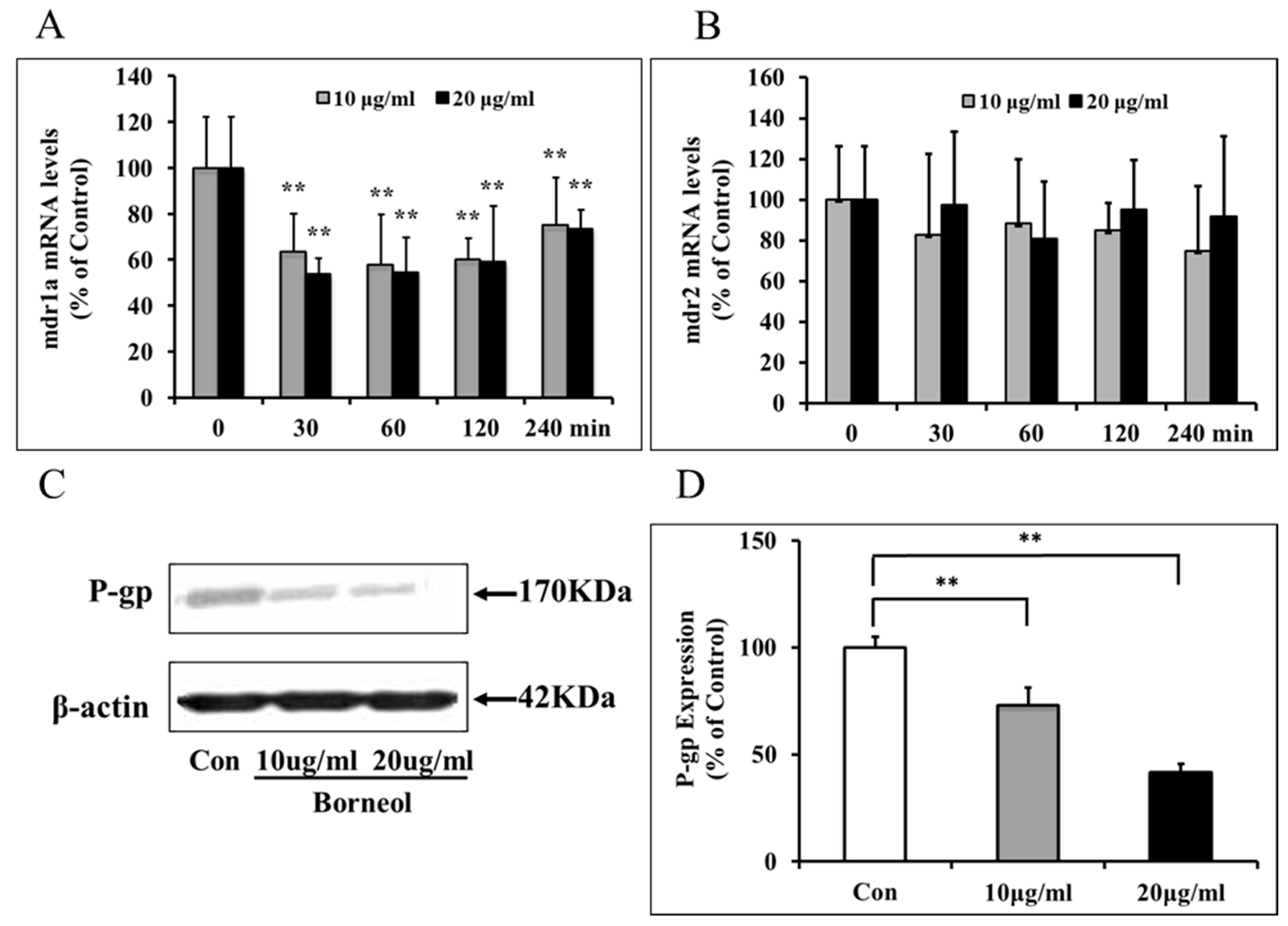
2.3. Effects of Borneol on mdr mRNA and P-gp Expression

2.4. Depressed P-gp Expression in BMECs by Borneol via a NF-κB Mediated Mechanism
3. Discussion

4. Experimental Section
4.1. Materials
4.2. Isolation and Culture of Rat Brain Microvascular Endothelial Cells
4.3. Isolation and Culture of Rat Astrocytes
4.4. The Establishment of an in Vitro BBB Model
4.5. Measurement of Transendothelial Electrical Resistance (TEER)
4.6. Measurement of γ-GT Activity in BMECs of the in Vitro BBB Model
4.7. The Effects of Borneol on Rho123 Accumulation in BMECs
4.8. The Effects of Borneol on P-gp Substrates Transport through the in Vitro BBB Model
4.9. Real-Time RT-PCR Analysis
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abbott, N.J.; Romero, I.A. Transporting therapeutics across the blood-brain barrier. Mol. Med. Today 1996, 2, 106–113. [Google Scholar] [CrossRef]
- Schinkel, A.H. P-Glycoprotein, a gatekeeper in the blood-brain barrier. Adv. Drug Deliv. Rev. 1999, 36, 179–194. [Google Scholar] [CrossRef]
- Miller, D.S.; Nobmann, S.N.; Gutmann, H.; Toeroek, M.; Drewe, J.; Fricker, G. Xenobiotic transport across isolated brain microvessels studied by confocal microscopy. Mol. Pharm. 2000, 58, 1357–1367. [Google Scholar]
- Litman, T.; Druley, T.E.; Stein, W.D.; Bates, S.E. From MDR to MXR: New understanding of multidrug resistance systems, their properties and clinical significance. Cell. Mol. Life Sci. 2001, 58, 931–959. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.F.; Sarangi, F.; Zastawny, R.L.; Veinot-Drebot, L.; Ling, V. Identification of members of the P-glycoprotein multigene family. Mol. Cell. Biol. 1989, 9, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Letrent, S.P.; Pollack, G.M.; Brouwer, K.R.; Brouwer, K.L. Effects of a potent and specific P-glycoprotein inhibitor on the blood-brain barrier distribution and antinociceptive effect of morphine in the rat. Drug Metab. Dispos. Biol. Fate Chem. 1999, 27, 827–834. [Google Scholar] [PubMed]
- Potschka, H.; Loscher, W. In vivo evidence for P-glycoprotein-mediated transport of phenytoin at the blood-brain barrier of rats. Epilepsia 2001, 42, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Van der Sandt, I.C.; Smolders, R.; Nabulsi, L.; Zuideveld, K.P.; de Boer, A.G.; Breimer, D.D. Active efflux of the 5-HT(1A) receptor agonist flesinoxan via P-glycoprotein at the blood-brain barrier. Eur. J. Pharm. Sci. 2001, 14, 81–86. [Google Scholar] [CrossRef]
- Glynn, S.L.; Yazdanian, M. In vitro blood-brain barrier permeability of nevirapine compared to other HIV antiretroviral agents. J. Pharm. Sci. 1998, 87, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Karssen, A.M.; Meijer, O.C.; van der Sandt, I.C.; Lucassen, P.J.; de Lange, E.C.; de Boer, A.G.; de Kloet, E.R. Multidrug resistance P-glycoprotein hampers the access of cortisol but not of corticosterone to mouse and human brain. Endocrinology 2001, 142, 2686–2694. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, C.; Graff, C.L.; Pollack, G.M. Variable modulation of opioid brain uptake by P-glycoprotein in mice. Biochem. Pharm. 2004, 67, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.-Y.; Ping, Q.N.; Chen, Z.-P. The enhancing effect of synthetical borneol on the absorption of tetramethylpyrazine phosphate in mouse. Int. J. Pharm. 2007, 337, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Hou, S.; Li, Y.; Zhao, B.; Yang, Z.; Xu, S.; Pu, J. Effect of borneol on the distribution of gastrodin to the brain in mice via oral administration. J. Drug Target. 2008, 16, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Li, X.; Li, Y.; Wang, L.; Xue, M. Pharmacokinetic interaction between puerarin and edaravone, and effect of borneol on the brain distribution kinetics of puerarin in rats. J. Pharm. Pharm. 2010, 62, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Ruan, M.; Cui, X.B.; Guo, J.M.; Xu, L.; Dong, X.P. Effects of borneol on the pharmacokinetics of geniposide in cortex, hippocampus, hypothalamus and striatum of conscious rat by simultaneous brain microdialysis coupled with UPLC-MS. J. Pharm. Biomed. Anal. 2013, 77, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wu, D.; Wu, J.; Ou, Y.; Mu, C.; Han, B.; Zhang, Q. Improved blood-brain barrier distribution: Effect of borneol on the brain pharmacokinetics of kaempferol in rats by in vivo microdialysis sampling. J. Ethnopharm. 2015, 162, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liao, Q.; Yao, M.; Xu, X.; Zhou, Y.; Hou, X.; Xie, Z. Effect of natural borneol on the pharmacokinetics and distribution of nimodipine in mice. Eur. J. Drug Metab. Pharm. 2014, 39, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gong, X.; Lu, Y.; Du, S.; Yang, Z.; Bai, J.; Li, P.; Wu, H. Enhancing effect of borneol and muscone on geniposide transport across the human nasal epithelial cell monolayer. PLoS ONE 2014, 9, 101414. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Z.; Lu, Y.; Du, S.Y.; Shang, K.X.; Cai, C.B. Influence of borneol and muscone on geniposide transport through MDCK and MDCK-MDR1 cells as blood-brain barrier in vitro model. Int. J. Pharm. 2013, 456, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Wang, N.S. Effect of borneol on the intercellular tight junction and pinocytosis vesicles in vitro blood-brain barrier model. Zhongguo Zhong Xi Yi Jie He Za Zhi 2004, 24, 632–634. [Google Scholar] [PubMed]
- Yu, B.; Lu, G.H.; Sun, Y.; Lin, X.; Fang, T.H. Effect of electroacupuncture combined with intragastric administration of borneol on the permeability of blood-brain barrier in the mouse. Zhen Ci Yan Jiu 2011, 36, 335–340. [Google Scholar] [PubMed]
- Yu, B.; Ruan, M.; Dong, X.; Yu, Y.; Cheng, H. The mechanism of the opening of the blood-brain barrier by borneol: A pharmacodynamics and pharmacokinetics combination study. J. Ethnopharm. 2013, 150, 1096–1108. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, S.; Renftel, M.; Brillault, J.; Fenart, L.; Cecchelli, R.; Dehouck, M.P. Prediction of drug transport through the blood-brain barrier in vivo: A comparison between two in vitro cell models. Pharm. Res. 2002, 19, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, S.; Renftel, M. The use of in vitro cell culture models for mechanistic studies and as permeability screens for the blood-brain barrier in the pharmaceutical industry--background and current status in the drug discovery process. Vasc. Pharm. 2002, 38, 355–364. [Google Scholar] [CrossRef]
- Cohen-Kashi Malina, K.; Cooper, I.; Teichberg, V.I. Closing the gap between the in vivo and in vitro blood-brain barrier tightness. Brain Res. 2009, 1284, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Imaizumi, S.; Kato, I.; Yoshimoto, T. Isolation and culture of brain endothelial cells and establishment of in vitro blood-brain barrier model. Cell Transplant. 1994, 3, 35–37. [Google Scholar]
- Ichikawa, N.; Naora, K.; Hirano, H.; Hashimoto, M.; Masumura, S.; Iwamoto, K. Isolation and primary culture of rat cerebral microvascular endothelial cells for studying drug transport in vitro. J. Pharm. Toxicol. Methods 1996, 36, 45–52. [Google Scholar] [CrossRef]
- Wolburg, H.; Neuhaus, J.; Kniesel, U.; Krauss, B.; Schmid, E.M.; Ocalan, M.; Farrell, C.; Risau, W. Modulation of tight junction structure in blood-brain barrier endothelial cells. Effects of tissue culture, second messengers and cocultured astrocytes. J. Cell Sci. 1994, 107, 1347–1357. [Google Scholar] [PubMed]
- Griepp, E.B.; Dolan, W.J.; Robbins, E.S.; Sabatini, D.D. Participation of plasma membrane proteins in the formation of tight junctions by cultured epithelial cells. J. Cell Biol. 1983, 96, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, A.A.; Berman, J.W.; Rashbaum, W.K.; Lyman, W.D. Human fetal astrocytes induce the expression of blood-brain barrier specific proteins by autologous endothelial cells. Brain Res. 1993, 625, 238–243. [Google Scholar] [CrossRef]
- Hayashi, Y.; Nomura, M.; Yamagishi, S.; Harada, S.; Yamashita, J.; Yamamoto, H. Induction of various blood-brain barrier properties in non-neural endothelial cells by close apposition to co-cultured astrocytes. Glia 1997, 19, 13–26. [Google Scholar] [CrossRef]
- Fontaine, M.; Elmquist, W.F.; Miller, D.W. Use of rhodamine 123 to examine the functional activity of P-glycoprotein in primary cultured brain microvessel endothelial cell monolayers. Life Sci. 1996, 59, 1521–1531. [Google Scholar] [CrossRef]
- Kageyama, M.; Namiki, H.; Fukushima, H.; Terasaka, S.; Togawa, T.; Tanaka, A.; Ito, Y.; Shibata, N.; Takada, K. Effect of chronic administration of ritonavir on function of cytochrome P450 3A and P-glycoprotein in rats. Biol. Pharm. Bull. 2005, 28, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Schinkel, A.H.; Smit, J.J.; van Tellingen, O.; Beijnen, J.H.; Wagenaar, E.; van Deemter, L.; Mol, C.A.; van der Valk, M.A.; Robanus-Maandag, E.C.; te Riele, H.P.; et al. Disruption of the mouse mdr1a P-glycoprotein gene leads to a deficiency in the blood-brain barrier and to increased sensitivity to drugs. Cell 1994, 77, 491–502. [Google Scholar] [CrossRef]
- Fardel, O.; Lecureur, V.; Guillouzo, A. The P-glycoprotein multidrug transporter. Gen. Pharm. 1996, 27, 1283–1291. [Google Scholar] [CrossRef]
- Kuo, M.T.; Liu, Z.; Wei, Y.; Lin-Lee, Y.C.; Tatebe, S.; Mills, G.B.; Unate, H. Induction of human MDR1 gene expression by 2-acetylaminofluorene is mediated by effectors of the phosphoinositide 3-kinase pathway that activate NF-κB signaling. Oncogene 2002, 21, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Felix, R.A.; Barrand, M.A. P-glycoprotein expression in rat brain endothelial cells: Evidence for regulation by transient oxidative stress. J. Neurochem. 2002, 80, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Nwaozuzu, O.M.; Sellers, L.A.; Barrand, M.A. Signalling pathways influencing basal and H2O2-induced P-glycoprotein expression in endothelial cells derived from the blood-brain barrier. J. Neurochem. 2003, 87, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Bentires-Alj, M.; Barbu, V.; Fillet, M.; Chariot, A.; Relic, B.; Jacobs, N.; Gielen, J.; Merville, M.P.; Bours, V. NF-κB transcription factor induces drug resistance through MDR1 expression in cancer cells. Oncogene 2003, 22, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Thevenod, F.; Friedmann, J.M.; Katsen, A.D.; Hauser, I.A. Up-regulation of multidrug resistance P-glycoprotein via nuclear factor-κB activation protects kidney proximal tubule cells from cadmium- and reactive oxygen species-induced apoptosis. J. Biol. Chem. 2000, 275, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Ros, J.E.; Schuetz, J.D.; Geuken, M.; Streetz, K.; Moshage, H.; Kuipers, F.; Manns, M.P.; Jansen, P.L.; Trautwein, C.; Muller, M. Induction of Mdr1b expression by tumor necrosis factor-α in rat liver cells is independent of p53 but requires NF-κB signaling. Hepatology 2001, 33, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Ogretmen, B.; Safa, A.R. Negative regulation of MDR1 promoter activity in MCF-7, but not in multidrug resistant MCF-7/Adr, cells by cross-coupled NF-κB/p65 and c-Fos transcription factors and their interaction with the CAAT region. Biochemistry 1999, 38, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Sukhai, M.; Piquette-Miller, M. Regulation of the multidrug resistance genes by stress signals. J. Pharm. Pharm. Sci. 2000, 3, 268–280. [Google Scholar] [PubMed]
- Abbott, N.J.; Hughes, C.C.; Revest, P.A.; Greenwood, J. Development and characterisation of a rat brain capillary endothelial culture: Towards an in vitro blood-brain barrier. J. Cell Sci. 1992, 103, 23–37. [Google Scholar] [PubMed]
- McCarthy, K.D.; de Vellis, J. Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef] [PubMed]
- Rutten, M.J.; Hoover, R.L.; Karnovsky, M.J. Electrical resistance and macromolecular permeability of brain endothelial monolayer cultures. Brain Res. 1987, 425, 301–310. [Google Scholar] [CrossRef]
- Orlowski, M.; Meister, A. The γ-glutamyl cycle: A possible transport system for amino acids. Proc. Nat. Acad. Sci. USA 1970, 67, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guo, Y.; Zhao, X.; Li, H.; Fan, G.; Mao, H.; Miao, L.; Gao, X. An estrogen receptor dependent mechanism of Oroxylin A in the repression of inflammatory response. PLoS ONE 2013, 8, 69555. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, X.; Zhang, B. Tanshinone: An inhibitor of proliferation of vascular smooth muscle cells. J. Ethnopharm. 2005, 99, 93–98. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Chai, L.; Zhang, H.; Wang, Y.; Zhang, B.; Gao, X. Borneol Depresses P-Glycoprotein Function by a NF-κB Signaling Mediated Mechanism in a Blood Brain Barrier in Vitro Model. Int. J. Mol. Sci. 2015, 16, 27576-27588. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161126051
Fan X, Chai L, Zhang H, Wang Y, Zhang B, Gao X. Borneol Depresses P-Glycoprotein Function by a NF-κB Signaling Mediated Mechanism in a Blood Brain Barrier in Vitro Model. International Journal of Molecular Sciences. 2015; 16(11):27576-27588. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161126051
Chicago/Turabian StyleFan, Xiang, Lijuan Chai, Han Zhang, Yuefei Wang, Boli Zhang, and Xiumei Gao. 2015. "Borneol Depresses P-Glycoprotein Function by a NF-κB Signaling Mediated Mechanism in a Blood Brain Barrier in Vitro Model" International Journal of Molecular Sciences 16, no. 11: 27576-27588. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161126051