
gga-miR-101-3p Plays a Key Role in Mycoplasma gallisepticum (HS Strain) Infection of Chicken

 and
and
Abstract
:1. Introduction
2. Results
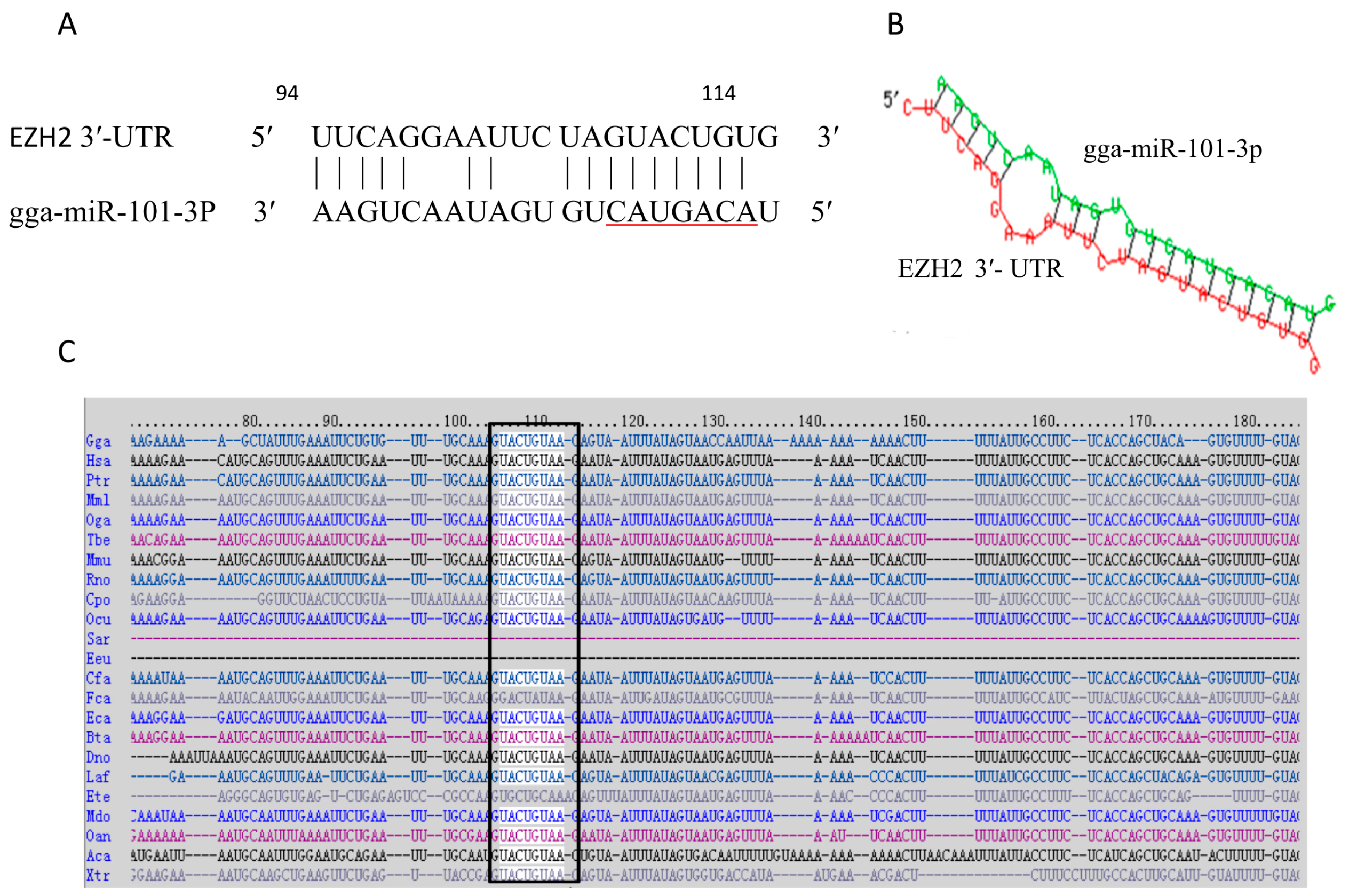
2.1. Prediction of the Target Gene of gga-miR-101-3p

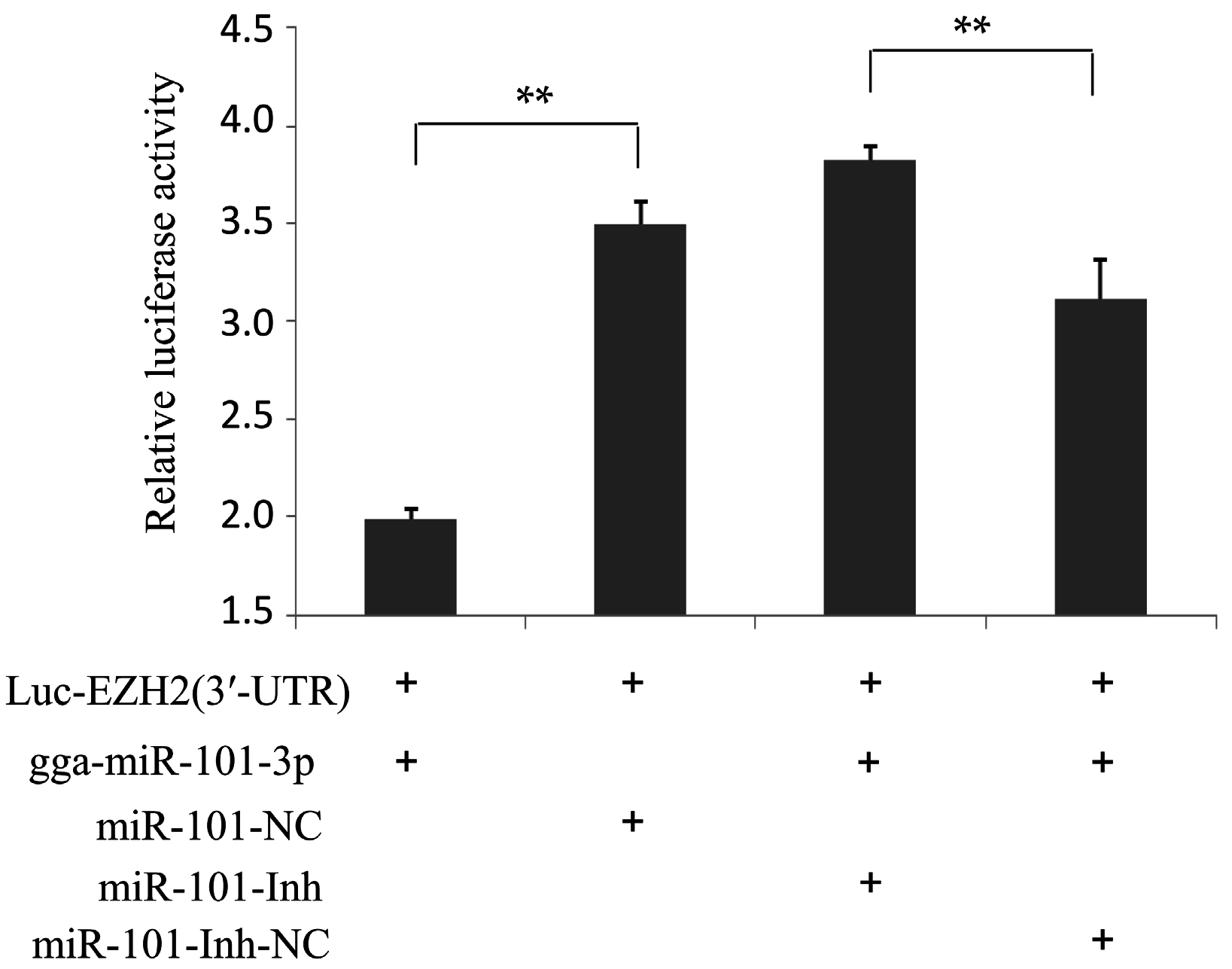
2.2. EZH2 Is the Direct Target of gga-miR-101-3p

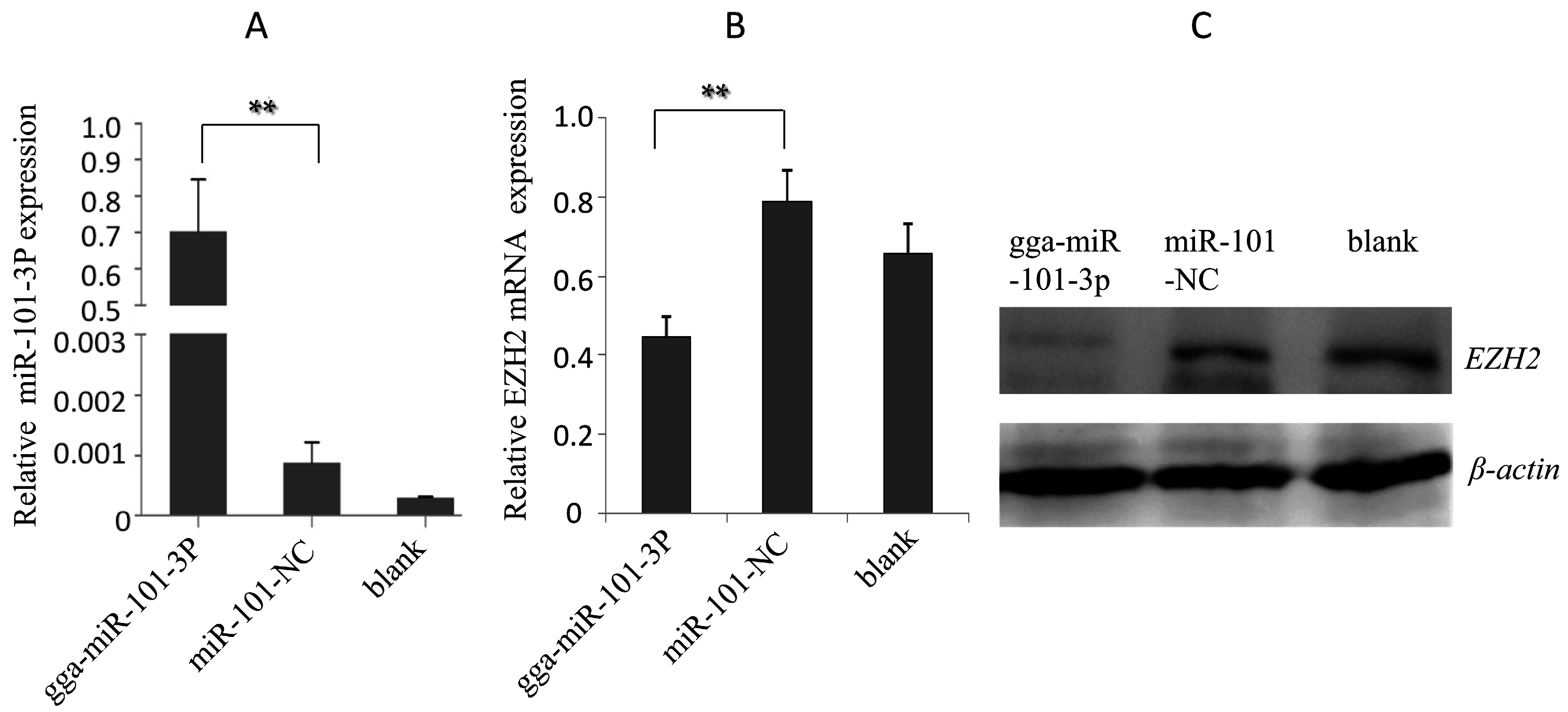
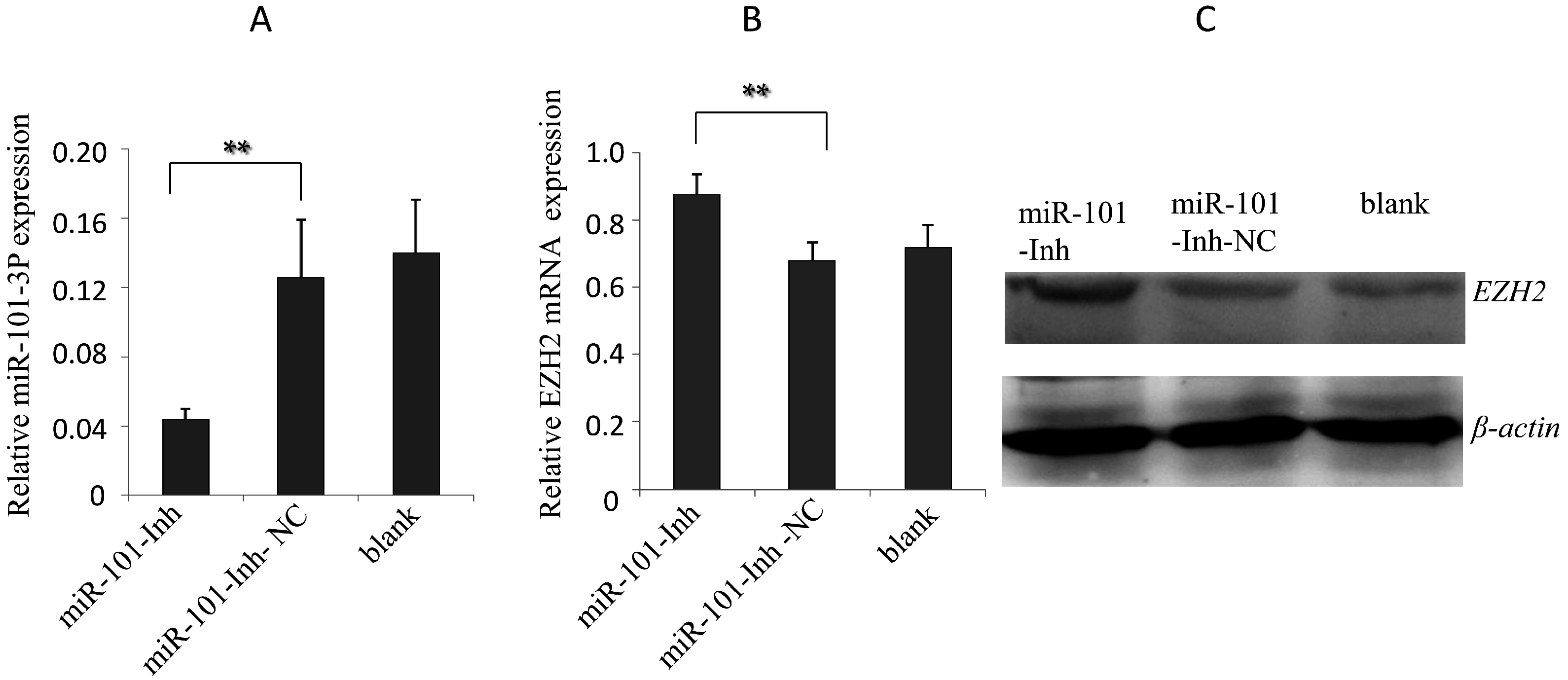
2.3. Effects of gga-miR-101-3p on EZH2 Expression


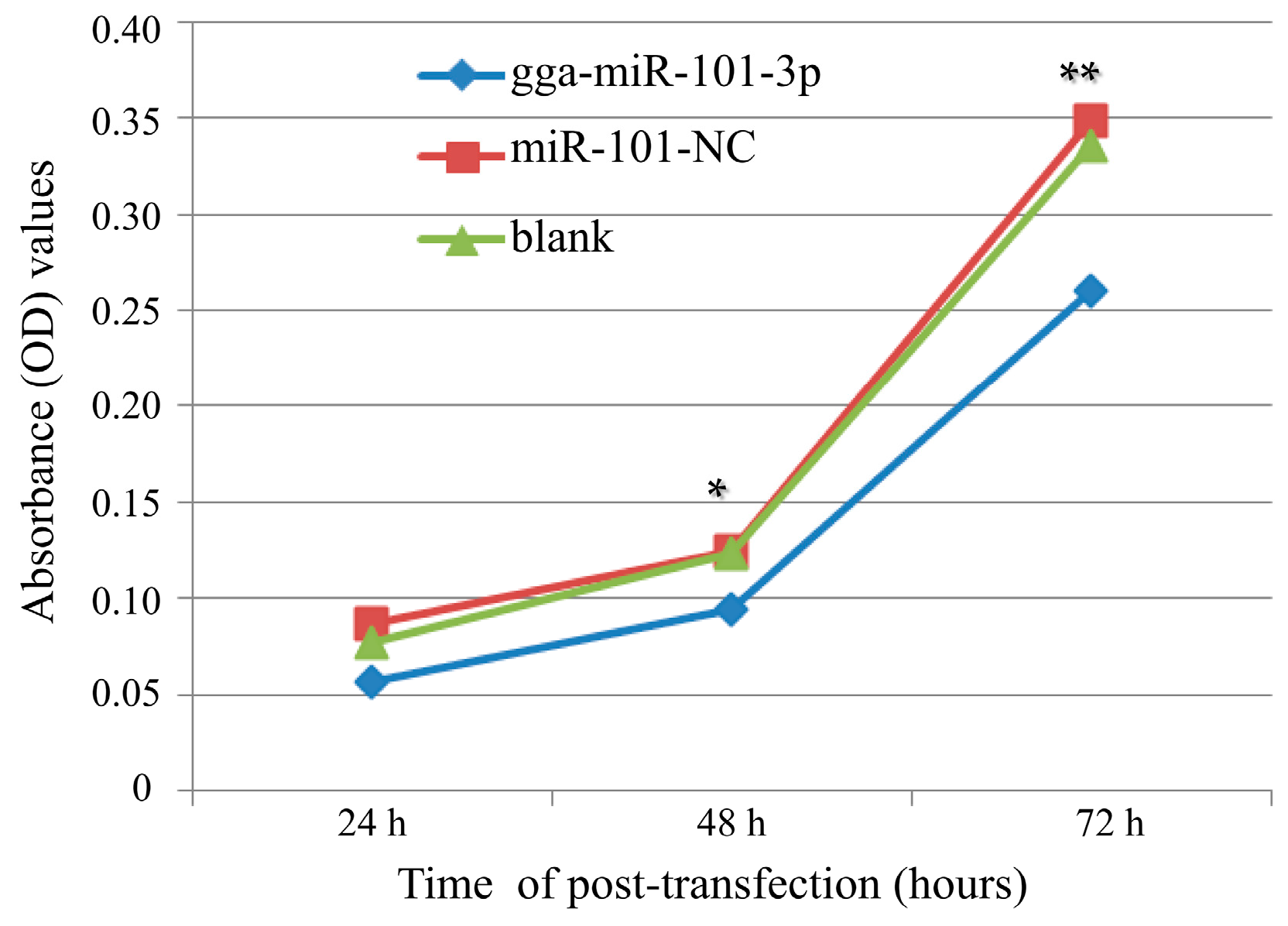
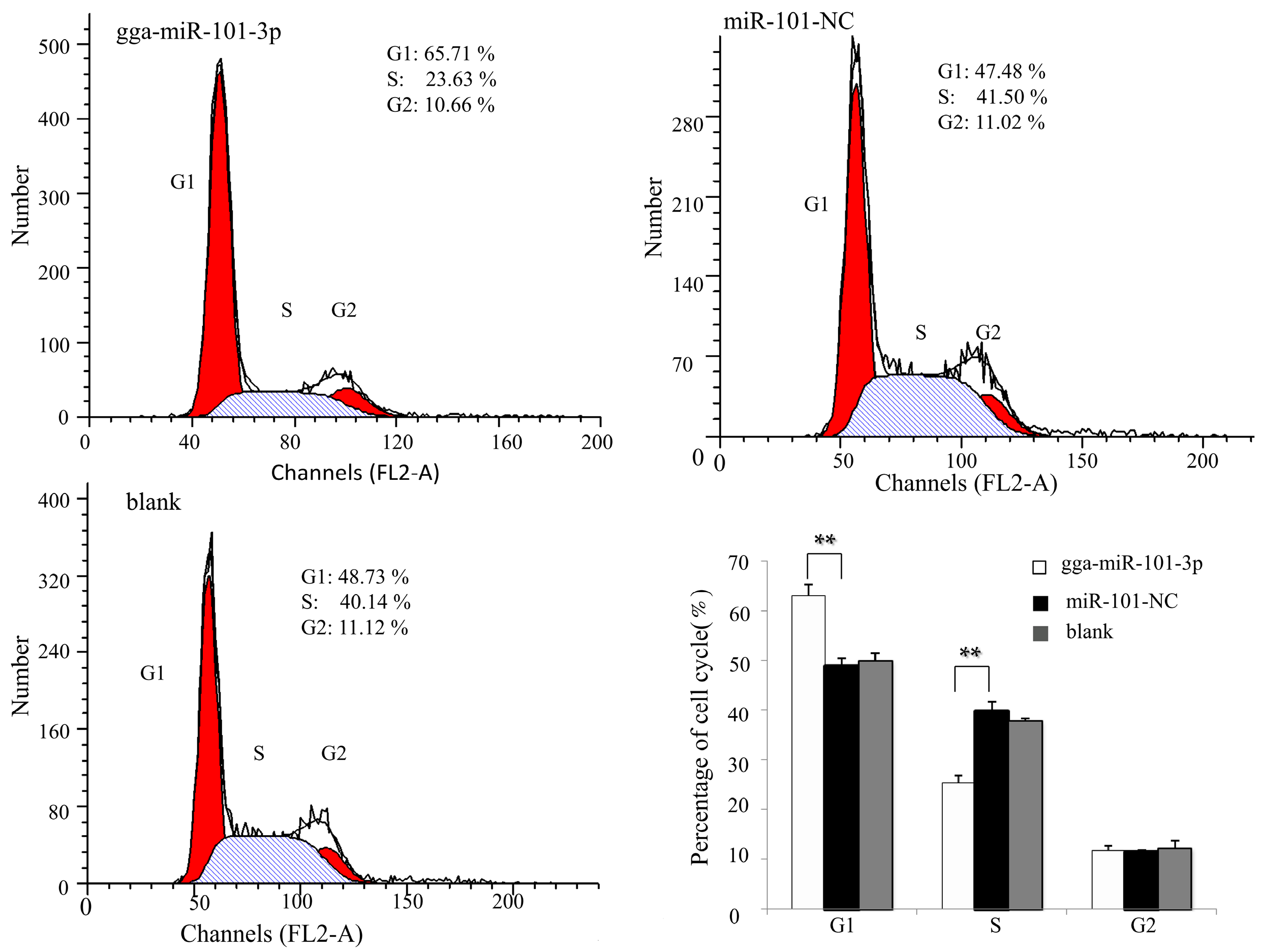
2.4. Effects of gga-miR-101-3P on Cell Proliferation and Cell Cycle


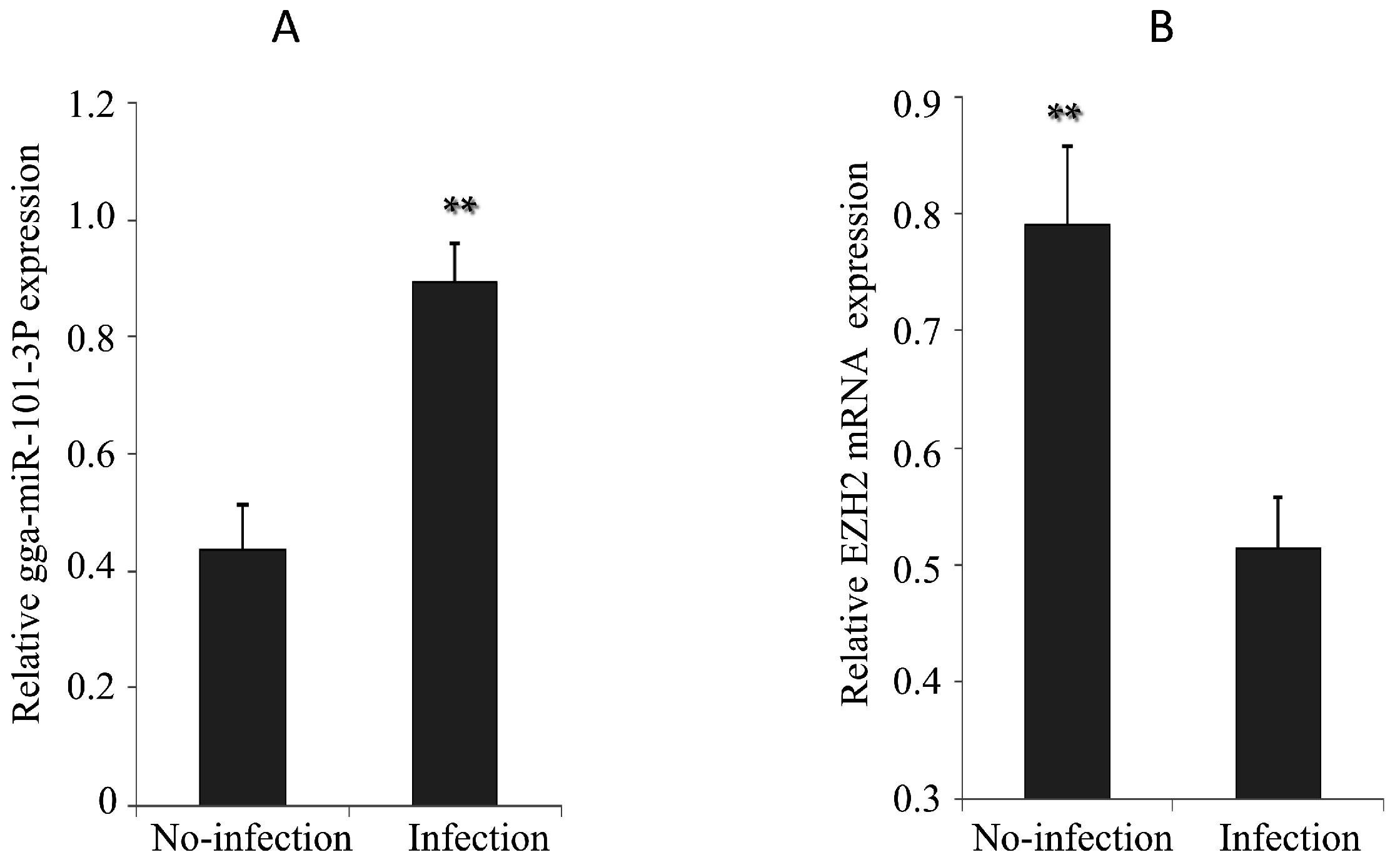
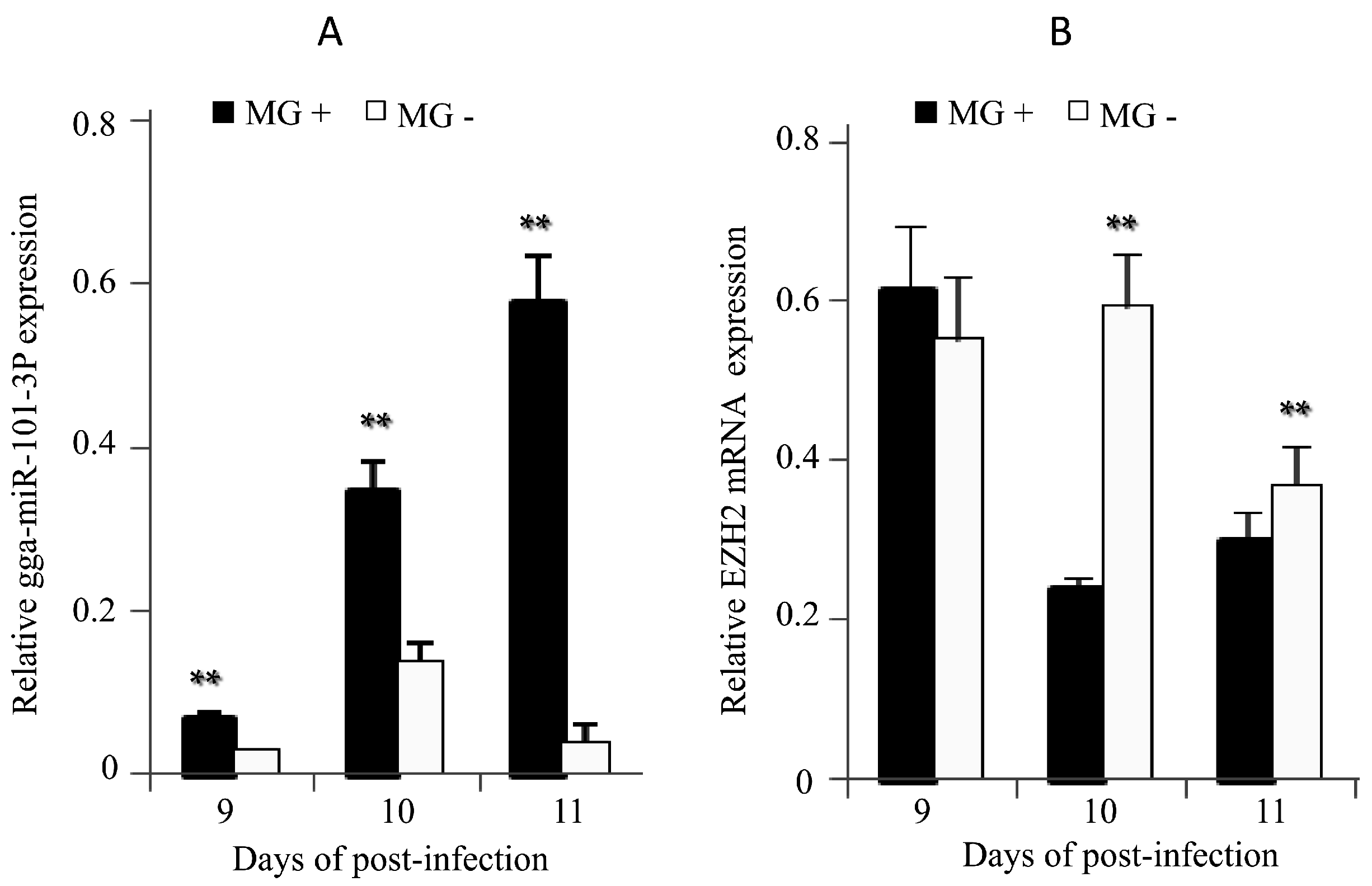
2.5. Expression of gga-miR-101-3p and EZH2 in MG-infected DF-1 Cells and Chicken Embryos


3. Discussion
4. Materials and Methods
4.1. Predication of gga-miR-101-3p Target Genes
4.2. Design of DNA Primers and Synthesis of RNA Oligonucleotides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence (5′–3′) | Accession No. |
|---|---|---|
| Primers for 3′-UTR Cloning | ||
| EZH2 3′-UTR-F | GCGCTCGAGCTGCCTTATCTTC | XM 004935061 |
| EZH2 3′-UTR-R | ATAGCGGCCGCTAGCTGGTGAGA | XM 004935061 |
| Primers for RT-qPCR | ||
| GAPDH-F | GAGGGTAGTGAAGGCTGCTG | NM 204305 |
| GAPDH-R | CACAACACGGTTGCTGTATC | NM 204305 |
| EZH2-F | TGCCTATAATGTACTCATGGTCAC | XM 004935061 |
| EZH2-R | TCATCATTGATAAATCCACACTCT | XM 004935061 |
| RT-gga-miR-101-3p | CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTTCAGTTA | MIMAT0001185 |
| gga-miR-101-3p-F | CTGGTAGGGTACAGTACTGTGATA | MIMAT0001185 |
| gga-miR-101-3p-R | CTGGTGTCGTGGAGTCGGC | MIMAT0001185 |
| gga-5s-rRNA-F | CCATACCACCCTGGAAACGC | |
| gga-5s-rRNA-R | TACTAACCGAGCCCGACCCT | |
4.3. Cell Culture
4.4. Plasmid Construction
4.5. Dual-Luciferase Reporter Assay
4.6. Reverse Transcription and Quantitative RT-PCR Analysis
4.7. Western Blot Analysis
4.8. Cell Proliferation and Cell Cycle Assay
4.9. Mycoplasma Strains and Growth Condition
4.10. Infection Experiments
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chazel, M.; Tardy, F.; le Grand, D.; Calavas, D.; Poumarat, F. Mycoplasmoses of ruminants in France: Recent data from the national surveillance network. BMC Vet. Res. 2010, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, T.P.; Balish, M.F.; Waites, K.B. Epidemiology, clinical manifestations, pathogenesis and laboratory detection of Mycoplasma pneumoniae infections. FEMS Microbiol. Rev. 2008, 32, 956–973. [Google Scholar] [CrossRef] [PubMed]
- Gambarini, M.; Kunz, T.; Oliveira Filho, B.; Porto, R.; Oliveira, C.; Brito, W.; Viu, M. Granular Vulvovagini Syndrome in nelore pubertal and post pubertal replacement heifers under tropical conditions: Role of Mycoplasma spp., Ureaplasma diversum and BHV-1. Trop. Anim. Health Prod. 2009, 41, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- May, M.; Brown, D.R. Secreted sialidase activity of canine Mycoplasmas. Vet. Microbiol. 2009, 137, 380–383. [Google Scholar] [CrossRef] [PubMed]
- McVey, J.P. An overview of the US/Japan natural resource’s panel on aquaculture: Past, present and future. Bull. Fish. Res. Agency 2010, 29, 1–8. [Google Scholar]
- Osman, K.; Aly, M.; Amin, Z.; Hasan, B. Mycoplasma gallisepticum: An emerging challenge to the poultry industry in Egypt. Rev. Sci. Technol. 2009, 28, 1015–1023. [Google Scholar]
- Sykes, J.E. Feline hemotropic Mycoplasmas. J. Vet. Emerg. Crit. Care 2010, 20, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Ley, D.H. Mycoplasma gallisepticum infection. In Diseases of Poultry, 11th ed.; Iowa State Univerity Press: Ames, IA, USA, 2003; pp. 722–743. [Google Scholar]
- Yoder, H.W., Jr. Mycoplasma gallisepticum infection. Dis. Poult. 1991, 9, 198–212. [Google Scholar]
- Davidson, W.; Nettles, V.; Couvillion, C.; Yoder, H., Jr. Infectious sinusitis in wild turkeys. Avian Dis. 1982, 26, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Jordan, F. Infection of the chicken with a virulent or avirulent strain of Mycoplasma gallisepticum alone and together with Newcastle disease virus or E. coli or both. Vet. Microbiol. 1979, 4, 35–45. [Google Scholar] [CrossRef]
- Stipkovits, L.; Egyed, L.; Palfi, V.; Beres, A.; Pitlik, E.; Somogyi, M.; Szathmary, S.; Denes, B. Effect of low-pathogenicity influenza virus H3N8 infection on Mycoplasma gallisepticum infection of chickens. Avian Pathol. 2012, 41, 51–57. [Google Scholar] [CrossRef]
- Fürnkranz, U.; Siebert-Gulle, K.; Rosengarten, R.; Szostak, M.P. Factors influencing the cell adhesion and invasion capacity of Mycoplasma gallisepticum. Acta Vet. Scand. 2013, 55, 63. [Google Scholar] [CrossRef] [PubMed]
- Indiková, I.; Much, P.; Stipkovits, L.; Siebert-Gulle, K.; Szostak, M.P.; Rosengarten, R.; Citti, C. Role of the GapA and CrmA cytadhesins of Mycoplasma gallisepticum in promoting virulence and host colonization. Infect. Immunity 2013, 81, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Geary, S.J.; Gladd, M.; Djordjevic, S.P. The Mycoplasma gallisepticum OsmC-like protein MG1142 resides on the cell surface and binds heparin. Microbiology 2007, 153, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Winner, F.; Rosengarten, R.; Citti, C. In vitro cell invasion of Mycoplasma gallisepticum. Infect. Immun. 2000, 68, 4238–4244. [Google Scholar] [CrossRef] [PubMed]
- Bi, D.; Ji, X. The isolation and identification of the Mycoplasma gallisepticum. Acta Vet. Zootechnol. Sin. 1988, 1, 146–148. [Google Scholar]
- Bi, D.; Xu, Q. Study on pathogenicity of HS strain Mycoplasma gallisepticum. Chin. J. Anim. Poult. Infect. Dis. 1997, 5, 24–26. [Google Scholar]
- Marois, C.; Dufour-Gesbert, F.; Kempf, I. Polymerase chain reaction for detection of Mycoplasma gallisepticum in environmental samples. Avian Pathol. 2002, 31, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Pennycott, T.; Dare, C.; Yavari, C.; Bradbury, J. Mycoplasma sturni and Mycoplasma gallisepticum in wild birds in Scotland. Vet. Rec. 2005, 156, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- miRBase: The microRNA Database. Available online: http://www.mirbase.org/cgi-bin/query.pl?terms=gga (accessed on 27 January 2015).
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Shyu, A.B.; Wilkinson, M.F.; van Hoof, A. Messenger RNA regulation: To translate or to degrade. EMBO J. 2008, 27, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Control of protein synthesis and mRNA degradation by microRNAs. Curr. Opin. Cell Biol. 2008, 20, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Rigoutsos, I. New tricks for animal microRNAS: Targeting of amino acid coding regions at conserved and nonconserved sites. Cancer Res. 2009, 69, 3245–3248. [Google Scholar] [CrossRef] [PubMed]
- Di Leva, G.; Calin, G.A.; Croce, C.M. MicroRNAs: Fundamental facts and involvement in human diseases. Birth Defects Res. C Embryo Today 2006, 78, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Glasner, M.E.; Yekta, S.; Burge, C.B.; Bartel, D.P. Vertebrate microRNA genes. Science 2003, 299, 1540. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shang, H.; Shu, D.; Zhang, H.; Ji, J.; Sun, B.; Li, H.; Xie, Q. gga-miR-375 plays a key role in tumorigenesis post subgroup J avian leukosis virus infection. PLoS ONE 2014, 9, e90878. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Gao, Y.; Ji, X.; Qi, X.; Qin, L.; Gao, H.; Wang, Y.; Wang, X. Differential expression of microRNAs in avian leukosis virus subgroup J-induced tumors. Vet. Microbiol. 2013, 162, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Brahmakshatriya, V.; Zhu, H.; Lupiani, B.; Reddy, S.M.; Yoon, B.J.; Gunaratne, P.H.; Kim, J.H.; Chen, R.; Wang, J. Identification of differentially expressed miRNAs in chicken lung and trachea with avian influenza virus infection by a deep sequencing approach. BMC Genom. 2009, 10, 512. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Ouyang, W.; Pan, Q.X.; Wang, X.L.; Xia, X.X.; Bi, Z.W.; Wang, Y.Q.; Wang, X.M. Overexpression of microRNA gga-miR-21 in chicken fibroblasts suppresses replication of infectious bursal disease virus through inhibiting VP1 translation. Antivir. Res. 2013, 100, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhao, Y.; Xu, H.; Smith, L.P.; Lawrie, C.H.; Watson, M.; Nair, V. MicroRNA profile of Marek’s disease virus-transformed T-cell line MSB-1: Predominance of virus-encoded microRNAs. J. Virol. 2008, 82, 4007–4015. [Google Scholar] [CrossRef] [PubMed]
- Lian, L.; Qu, L.; Chen, Y.; Lamont, S.J.; Yang, N. A systematic analysis of miRNA transcriptome in Marek’s disease virus-induced lymphoma reveals novel and differentially expressed miRNAs. PLoS ONE 2012, 7, e51003. [Google Scholar] [CrossRef] [PubMed]
- Stik, G.; Dambrine, G.; Pfeffer, S.; Rasschaert, D. The oncogenic microRNA OncomiR-21 overexpressed during Marek’s disease lymphomagenesis is transactivated by the viral oncoprotein Meq. J. Virol. 2013, 87, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lian, L.; Zhang, D.; Qu, L.; Yang, N. gga-miR-26a targets NEK6 and suppresses Marek’s disease lymphoma cell proliferation. Poult. Sci. 2014, 93, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Jeong, W.; Kim, J.H.; Kim, J.; Bazer, F.W.; Han, J.Y.; Song, G. Distinct expression pattern and post-transcriptional regulation of cell cycle genes in the glandular epithelia of avian ovarian carcinomas. PLoS ONE 2012, 7, e51592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.G.; Guo, J.F.; Liu, D.L.; Liu, Q.; Wang, J.J. MicroRNA-101 exerts tumor-suppressive functions in non-small cell lung cancer through directly targeting enhancer of zeste homolog 2. J. Thorac. Oncol. 2011, 6, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; Jeon, H.S.; Lee, S.Y.; Jeong, K.J.; Park, S.Y.; Lee, H.Y.; Lee, J.U.; Kwon, S.J.; Choi, E.; Na, M.J. microRNA-101 inhibits lung cancer invasion through the regulation of enhancer of zeste homolog 2. Exp. Ther. Med. 2011, 2, 963–967. [Google Scholar] [PubMed]
- Yang, J.; Lu, Y.; Lu, M.; Leng, R.; Pan, H.; Ye, D. MicroRNA-101, mitogen-activated protein kinases and mitogen-activated protein kinases phosphatase-1 in systemic lupus erythematosus. Lupus 2013, 22, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.Y.; Liu, Q.; Chen, J.X.; Lan, K.; Ge, B.X. MicroRNA-101 targets MAPK phosphatase-1 to regulate the activation of MAPKs in macrophages. J. Immunol. 2010, 185, 7435–7442. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Lin, C.; Shi, Y.; Kuerban, G. MicroRNA-101 inhibits cell proliferation, invasion, and promotes apoptosis by regulating cyclooxygenase-2 in HeLa cervical carcinoma cells. Asian Pac. J. Cancer Prev. 2013, 14, 5915. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Target Scan Human. Available online: http://www.targetscan.org/ (accessed on 10 November 2014).
- miRecords. Available online: http://c1.accurascience.com/miRecords/ (accessed on 10 November 2014).
- miRDB. Available online: http://www.mirdb.org/miRDB/ (accessed on 10 November 2014).
- RNAhybrid. Available online: http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/ (accessed on 10 November 2014).
- AmiGO 2. Available online: http://amigo.geneontology.org (accessed on 10 December 2014).
- Wu, C.; Chen, Z.; Kuchroo, V.K. EZH2 lines up the chromatin in T regulatory cells. Immunity 2015, 42, 201–203. [Google Scholar] [CrossRef] [PubMed]
- DuPage, M.; Chopra, G.; Quiros, J.; Rosenthal, W.L.; Morar, M.M.; Holohan, D.; Zhang, R.; Turka, L.; Marson, A.; Bluestone, J.A. The chromatin-modifying enzyme EZH2 is critical for the maintenance of regulatory T cell identity after activation. Immunity 2015, 42, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, N.; Okamoto, K.; Sawa, S.; Nakashima, T.; Oh-hora, M.; Kodama, T.; Tanaka, S.; Bluestone, J.A.; Takayanagi, H. Pathogenic conversion of Foxp3+ T cells into TH17 cells in autoimmune arthritis. Nat. Med. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Su, I.H.; Basavaraj, A.; Krutchinsky, A.N.; Hobert, O.; Ullrich, A.; Chait, B.T.; Tarakhovsky, A. EZH2 controls B cell development through histone H3 methylation and Igh rearrangement. Nat. Immunol. 2003, 4, 124–131. [Google Scholar] [CrossRef] [PubMed]
- O’Carroll, D.; Erhardt, S.; Pagani, M.; Barton, S.C.; Surani, M.A.; Jenuwein, T. The Polycomb-group gene EZH2 Is required for early mouse development. Mol. Cell. Biol. 2001, 21, 4330–4336. [Google Scholar] [CrossRef] [PubMed]
- http://www.gene.ucl.ac.uk/nomenclature/guidelines.html (accessed on 10 December 2014).
- Yan, J.; Gumireddy, K.; Li, A.; Huang, Q. Regulation of mesenchymal phenotype by MicroRNAs in cancer. Curr. Cancer Drug Targets 2013, 13, 930. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Coukos, G.; Zhang, L. miRNAs in human cancer. Methods Mol. Biol. 2012, 822, 295–306. [Google Scholar] [PubMed]
- Barton, G.M.; Medzhitov, R. Toll-like receptor signaling pathways. Science 2003, 300, 1524–1525. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B. Inferences, questions and possibilities in Toll-like receptor signalling. Nature 2004, 430, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Muzio, M.; Polentarutti, N.; Bosisio, D.; Manoj, K.P.; Mantovani, A. Toll-like receptor family and signalling pathway. Biochem. Soc. Trans. 2000, 28, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Valente, A.J.; Yoshida, T.; Gardner, J.D.; Somanna, N.; Delafontaine, P.; Chandrasekar, B. Interleukin-17A stimulates cardiac fibroblast proliferation and migration via negative regulation of the dual-specificity phosphatase MKP-1/DUSP-1. Cell Signal. 2012, 24, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Tonini, T.; D’Andrilli, G.; Fucito, A.; Gaspa, L.; Bagella, L. Importance of EZH2 polycomb protein in tumorigenesis process interfering with the pathway of growth suppressive key elements. J. Cell. Physiol. 2008, 214, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, I.M.; Halvorsen, O.J.; Collett, K.; Stefansson, I.M.; Straume, O.; Haukaas, S.A.; Salvesen, H.B.; Otte, A.P.; Akslen, L.A. EZH2 expression is associated with high proliferation rate and aggressive tumor subgroups in cutaneous melanoma and cancers of the endometrium, prostate, and breast. J. Clin. Oncol. 2006, 24, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Varambally, S.; Cao, Q.; Mani, R.S.; Shankar, S.; Wang, X.; Ateeq, B.; Laxman, B.; Cao, X.; Jing, X.; Ramnarayanan, K. Genomic loss of microRNA-101 leads to overexpression of histone methyltransferase Ezh2 in cancer. Science 2008, 322, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, C.; Zhou, B.; Bi, L.; Huang, H.; Lin, T.; Xu, K. Role of EZH2 in the growth of prostate cancer stem cells isolated from LNCaP cells. Int. J. Mol. Sci. 2013, 14, 11981–11993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, P.; Deng, Z.; Wan, M.; Huang, W.; Cramer, S.D.; Xu, J.; Lei, M.; Sui, G. Research MicroRNA-101 negatively regulates EZH2 and its expression is modulated by androgen receptor and HIF-1α/HIF-1β. Mol. Cancer 2010, 9, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xiang, W.; Wang, M.; Huang, T.; Xiao, X.; Wang, L.; Tao, D.; Dong, L.; Zeng, F.; Jiang, G. Methyl jasmonate sensitizes human bladder cancer cells to gambogic acid-induced apoptosis through down-regulation of EZH2 expression by miR-101. Br. J. Pharmacol. 2014, 171, 618–635. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Liang, G.; Liu, C.C.; Wolff, E.M.; Tsai, Y.C.; Ye, W.; Zhou, X.; Jones, P.A. The putative tumor suppressor microRNA-101 modulates the cancer epigenome by repressing the polycomb group protein EZH2. Cancer Res. 2009, 69, 2623–2629. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Ruan, H.J.; He, X.J.; Ma, Y.Y.; Jiang, X.T.; Xia, Y.J.; Ye, Z.Y.; Tao, H.Q. MicroRNA-101 is down-regulated in gastric cancer and involved in cell migration and invasion. Eur. J. Cancer 2010, 46, 2295–2303. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Shen, F.; Wu, J.; Zhang, W.; Wang, J.; Zhang, L. miR-101, downregulated in retinoblastoma, functions as a tumor suppressor in human retinoblastoma cells by targeting EZH2. Oncol. Rep. 2014, 32, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, J.; Yu, L.; Cai, J.; Gui, T.; Tang, H.; Song, L.; Wang, J.; Han, F.; Yang, C. miR-101 regulates expression of EZH2 and contributes to progression of and cisplatin resistance in epithelial ovarian cancer. Tumor Biol. 2014, 35, 12619–12626. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, M.; Wu, H.; Ru, P.; Hwang, L.; Trieu, V.; Mo, Y. MicroRNA-101-mediated Akt activation and estrogen-independent growth. Oncogene 2011, 30, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.P.; Jiang, K.; Hirahara, K.; Vahedi, G.; Afzali, B.; Sciume, G.; Bonelli, M.; Sun, H.W.; Jankovic, D.; Kanno, Y. EZH2 is crucial for both differentiation of regulatory T cells and T effector cell expansion. Sci. Rep. 2015, 5, 10643. [Google Scholar] [CrossRef] [PubMed]
- Raaphorst, F.M.; Otte, A.P.; van Kemenade, F.J.; Blokzijl, T.; Fieret, E.; Hamer, K.M.; Satijn, D.P.; Meijer, C.J. Distinct BMI-1 and EZH2 expression patterns in thymocytes and mature T cells suggest a role for Polycomb genes in human T cell differentiation. J. Immunol. 2001, 166, 5925–5934. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Calus, D.; Maes, D.; Vranckx, K.; Villareal, I.; Pasmans, F.; Haesebrouck, F. Validation of ATP luminometry for rapid and accurate titration of Mycoplasma hyopneumoniae in Friis medium and a comparison with the color changing units assay. J. Microbiol. Methods 2010, 83, 335–340. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Wang, Z.; Bi, D.; Hou, Y.; Zhao, Y.; Sun, J.; Peng, X. gga-miR-101-3p Plays a Key Role in Mycoplasma gallisepticum (HS Strain) Infection of Chicken. Int. J. Mol. Sci. 2015, 16, 28669-28682. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226121
Chen J, Wang Z, Bi D, Hou Y, Zhao Y, Sun J, Peng X. gga-miR-101-3p Plays a Key Role in Mycoplasma gallisepticum (HS Strain) Infection of Chicken. International Journal of Molecular Sciences. 2015; 16(12):28669-28682. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226121
Chicago/Turabian StyleChen, Jiao, Zaiwei Wang, Dingren Bi, Yue Hou, Yabo Zhao, Jianjun Sun, and Xiuli Peng. 2015. "gga-miR-101-3p Plays a Key Role in Mycoplasma gallisepticum (HS Strain) Infection of Chicken" International Journal of Molecular Sciences 16, no. 12: 28669-28682. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226121