
IL-18 and Cutaneous Inflammatory Diseases


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
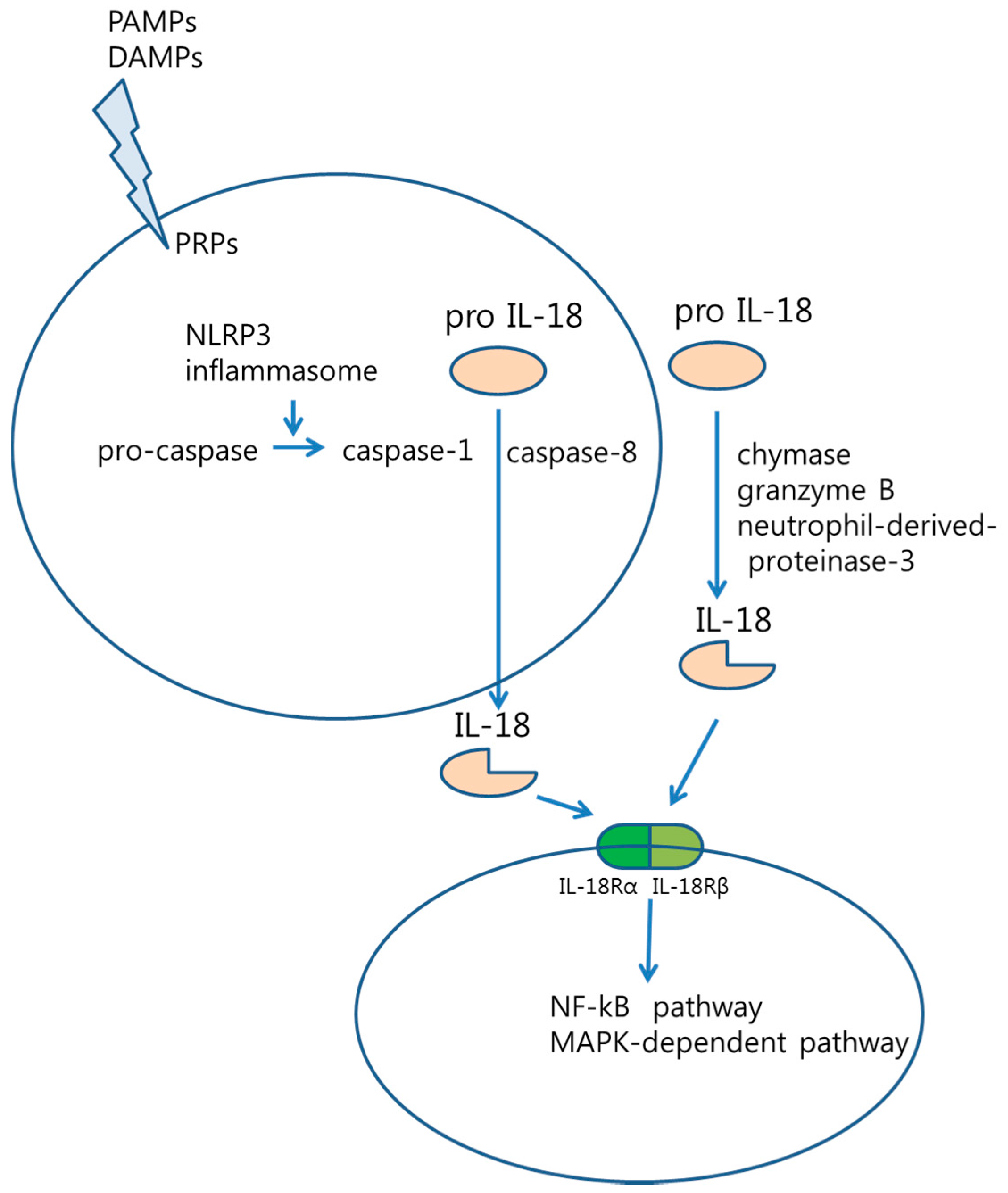
 : pro IL-18,
: pro IL-18,  : active IL-18,
: active IL-18,  : IL-18Rα,
: IL-18Rα,  : IL-18Rβ).
: pro IL-18, : active IL-18, : IL-18Rα, : IL-18Rβ).
: IL-18Rβ).
: pro IL-18, : active IL-18, : IL-18Rα, : IL-18Rβ).
2. IL-18 in Inflammatory and Autoimmune Cutaneous Diseases
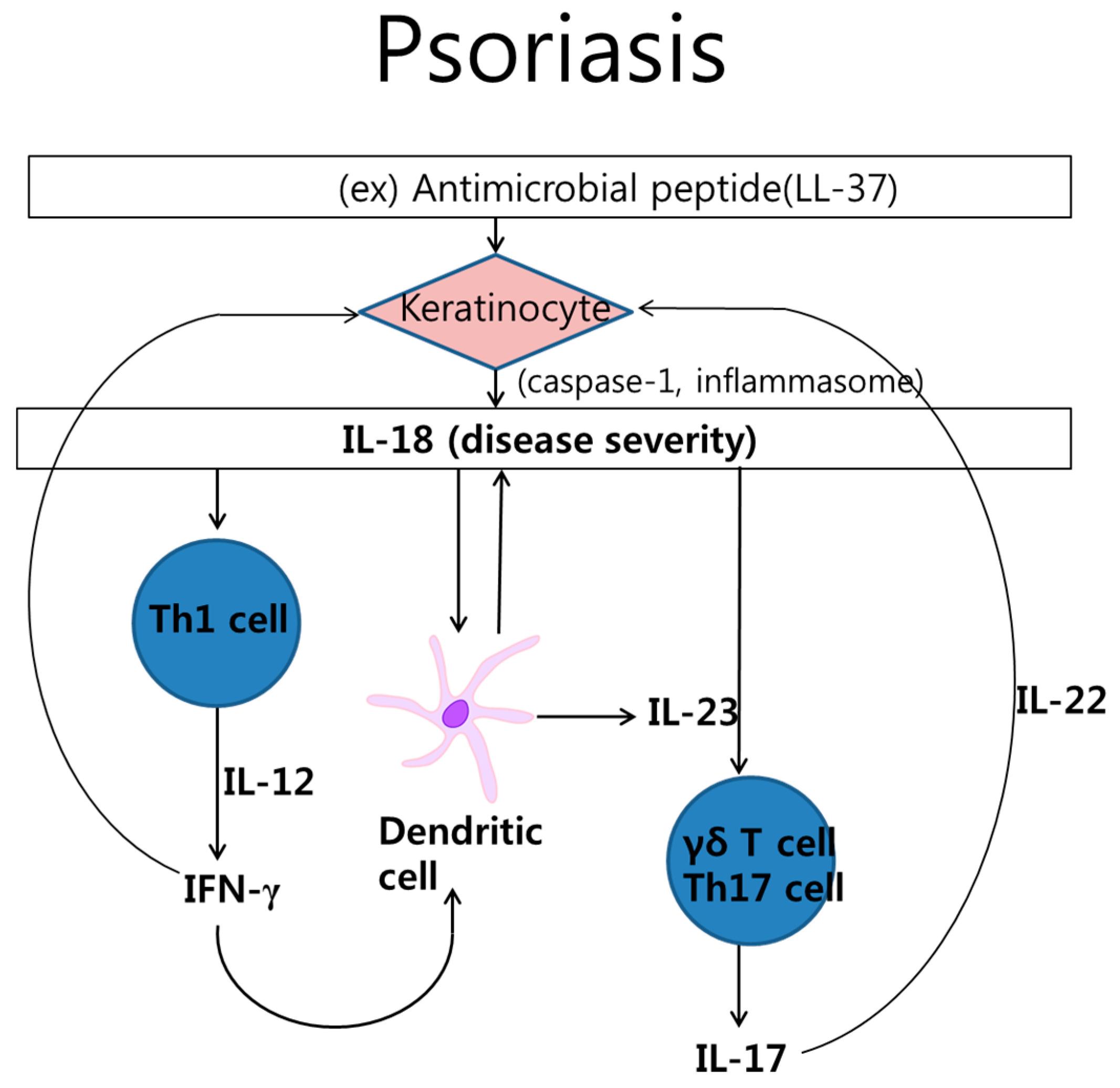
2.1. Psoriasis

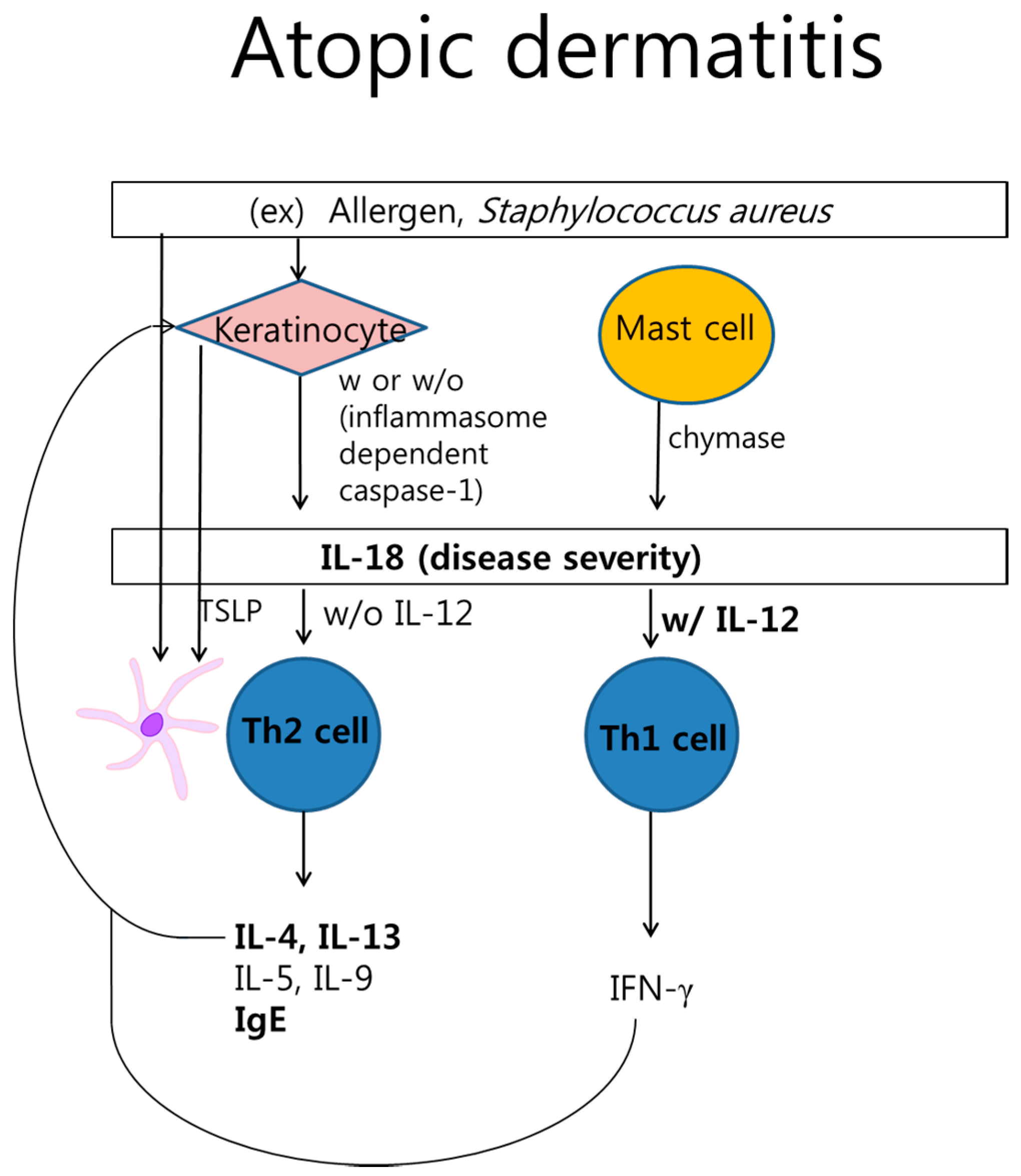
2.2. Atopic Dermatitis

2.3. Urticaria
2.4. Contact Dermatitis
2.5. Alopecia Areata
2.6. Drug Allergy and Cutaneous Drug Eruption
2.7. Cutaneous Lupus Erythematosus
2.8. Graft-Versus-Host Disease (GVHD)
2.9. Miscellaneous Diseases
3. Conclusions
4. Future Plans and Expert Commentary
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arend, W.P.; Palmer, G.; Gabay, C. IL-1, IL-18, and IL-33 families of cytokines. Immunol. Rev. 2008, 223, 20–38. [Google Scholar] [CrossRef]
- Okamura, H.; Nagata, K.; Komatsu, T.; Tanimoto, T.; Nukata, Y.; Tanabe, F.; Akita, K.; Torigoe, K.; Okura, T.; Fukuda, S.; et al. A novel costimulatory factor for γ interferon induction found in the livers of mice causes endotoxic shock. Infect. Immun. 1995, 63, 3966–3972. [Google Scholar] [PubMed]
- Nolan, K.F.; Greaves, D.R.; Waldmann, H. The human interleukin 18 gene IL18 maps to 11q22.2–q22.3, closely linked to the DRD2 gene locus and distinct from mapped IDDM loci. Genomics 1998, 51, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Bellora, F.; Castriconi, R.; Doni, A.; Cantoni, C.; Moretta, L.; Mantovani, A.; Moretta, A.; Bottino, C. M-CSF induces the expression of a membrane-bound form of IL-18 in a subset of human monocytes differentiating in vitro toward macrophages. Eur. J. Immunol. 2012, 42, 1618–1626. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, N.; Sukhova, G.K.; Libby, P.; Reynolds, R.S.; Young, J.L.; Schonbeck, U. Expression of interleukin (IL)-18 and functional IL-18 receptor on human vascular endothelial cells, smooth muscle cells, and macrophages: Implications for atherogenesis. J. Exp. Med. 2002, 195, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Moller, B.; Kessler, U.; Rehart, S.; Kalina, U.; Ottmann, O.G.; Kaltwasser, J.P.; Hoelzer, D.; Kukoc-Zivojnov, N. Expression of interleukin-18 receptor in fibroblast-like synoviocytes. Arthritis Res. 2002, 4, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.E. IL-1 and IL-18 receptors, and their extended family. Curr. Opin. Immunol. 2002, 14, 117–122. [Google Scholar] [CrossRef]
- Gutzmer, R.; Langer, K.; Mommert, S.; Wittmann, M.; Kapp, A.; Werfel, T. Human dendritic cells express the IL-18R and are chemoattracted to IL-18. J. Immunol. 2003, 171, 6363–6371. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, I.; Raffaghello, L.; Cocco, C.; Guglielmino, R.; Roncella, S.; Fedeli, F.; Gambini, C.; Pistoia, V. Heterogeneous expression of interleukin-18 and its receptor in B-cell lymphoproliferative disorders deriving from naive, germinal center, and memory B lymphocytes. Clin. Cancer Res. 2004, 10, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.B.; Ting, J.P. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Ogawa, Y.; Aoki, R.; Shimada, S. Innate and intrinsic antiviral immunity in skin. J. Dermatol. Sci. 2014, 75, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Dinarello, C.A. IL-18 in autoimmunity: Review. Eur. Cytokine Netw. 2006, 17, 224–252. [Google Scholar] [PubMed]
- Omoto, Y.; Tokime, K.; Yamanaka, K.; Habe, K.; Morioka, T.; Kurokawa, I.; Tsutsui, H.; Yamanishi, K.; Nakanishi, K.; Mizutani, H. Human mast cell chymase cleaves pro-IL-18 and generates a novel and biologically active IL-18 fragment. J. Immunol. 2006, 177, 8315–8319. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Kim, S.H.; Lewis, E.C.; Azam, T.; Reznikov, L.L.; Dinarello, C.A. Differences in signaling pathways by IL-1β and IL-18. Proc. Natl. Acad. Sci. USA 2004, 101, 8815–8820. [Google Scholar] [CrossRef] [PubMed]
- Barksby, H.E.; Lea, S.R.; Preshaw, P.M.; Taylor, J.J. The expanding family of interleukin-1 cytokines and their role in destructive inflammatory disorders. Clin. Exp. Immunol. 2007, 149, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 binding protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Novick, D.; Kim, S.H.; Fantuzzi, G.; Reznikov, L.L.; Dinarello, C.A.; Rubinstein, M. Interleukin-18 binding protein: A novel modulator of the Th1 cytokine response. Immunity 1999, 10, 127–136. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 regulates both Th1 and Th2 responses. Annu. Rev. Immunol. 2001, 19, 423–474. [Google Scholar] [CrossRef] [PubMed]
- Muhl, H.; Pfeilschifter, J. Interleukin-18 bioactivity: A novel target for immunopharmacological anti-inflammatory intervention. Eur. J. Pharmacol. 2004, 500, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.; Macdonald, A.; Renne, J. IL-18 and skin inflammation. Autoimmun. Rev. 2009, 9, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Lebre, M.C.; Antons, J.C.; Kalinski, P.; Schuitemaker, J.H.; van Capel, T.M.; Kapsenberg, M.L.; de Jong, E.C. Double-stranded RNA-exposed human keratinocytes promote Th1 responses by inducing a type-1 polarized phenotype in dendritic cells: Role of keratinocyte-derived tumor necrosis factor α, type I interferons, and interleukin-18. J. Investig. Dermatol. 2003, 120, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Drenker, M.; Eiz-Vesper, B.; Werfel, T.; Wittmann, M. Evidence for a pathogenetic role of interleukin-18 in cutaneous lupus erythematosus. Arthritis Rheum. 2008, 58, 3205–3215. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, J.E.; Lee, J.Y.; Cho, B.K.; Lee, W.J.; Kim, T.; Yoon, D.; Cho, D. Increased expression of IL-18 in cutaneous graft-versus-host disease. Immunol. Lett. 2004, 95, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Boehncke, W.H.; Schon, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Krueger, J.G. Hiding under the skin: A welcome surprise in psoriasis. Nat. Med. 2012, 18, 1750–1751. [Google Scholar] [CrossRef] [PubMed]
- Lessard, J.C.; Pina-Paz, S.; Rotty, J.D.; Hickerson, R.P.; Kaspar, R.L.; Balmain, A.; Coulombe, P.A. Keratin 16 regulates innate immunity in response to epidermal barrier breach. Proc. Natl. Acad. Sci. USA 2013, 110, 19537–1942. [Google Scholar] [CrossRef] [PubMed]
- Roth, W.; Kumar, V.; Beer, H.D.; Richter, M.; Wohlenberg, C.; Reuter, U.; Thiering, S.; Staratschek-Jox, A.; Hofmann, A.; Kreusch, F.; et al. Keratin 1 maintains skin integrity and participates in an inflammatory network in skin through interleukin-18. J. Cell Sci. 2012, 125 Pt 22, 5269–5279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, D.; Seung Kang, J.; Hoon Park, J.; Kim, Y.I.; Hahm, E.; Lee, J.; Yang, Y.; Jeon, J.; Song, H.; Park, H.; et al. The enhanced IL-18 production by UVB irradiation requires ROI and AP-1 signaling in human keratinocyte cell line (HaCaT). Biochem. Biophys. Res. Commun. 2002, 298, 289–295. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ushio, H.; Nagaoka, I.; Okumura, K.; Ogawa, H. The human β-defensins (-1, -2, -3, -4) and cathelicidin LL-37 induce IL-18 secretion through p38 and ERK MAPK activation in primary human keratinocytes. J. Immunol. 2005, 175, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Grando, S.A.; Li, Y.C. Regulation of IL-1 family cytokines IL-1α, IL-1 receptor antagonist, and IL-18 by 1,25-dihydroxyvitamin D3 in primary keratinocytes. J. Immunol. 2006, 176, 3780–3787. [Google Scholar] [CrossRef] [PubMed]
- Yawalkar, N.; Karlen, S.; Hunger, R.; Brand, C.U.; Braathen, L.R. Expression of interleukin-12 is increased in psoriatic skin. J. Investig. Dermatol. 1998, 111, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Sutton, C.E.; Mielke, L.A.; Mills, K.H. IL-17-producing γδ T cells and innate lymphoid cells. Eur. J. Immunol. 2012, 42, 2221–2231. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Hamada, Y.; Katsuoka, K. Expression of IL-18 in psoriasis. Arch. Dermatol. Res. 2001, 293, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Gangemi, S.; Merendino, R.A.; Guarneri, F.; Minciullo, P.L.; DiLorenzo, G.; Pacor, M.; Cannavo, S.P. Serum levels of interleukin-18 and s-ICAM-1 in patients affected by psoriasis: Preliminary considerations. J. Eur. Acad. Dermatol. Venereol. 2003, 17, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.; Moeller, K.; Kragballe, K.; Iversen, L. The activity of caspase-1 is increased in lesional psoriatic epidermis. J. Investig. Dermatol. 2007, 127, 2857–2864. [Google Scholar] [CrossRef] [PubMed]
- Companjen, A.; van der Wel, L.; van der Fits, L.; Laman, J.; Prens, E. Elevated interleukin-18 protein expression in early active and progressive plaque-type psoriatic lesions. Eur. Cytokine Netw. 2004, 15, 210–216. [Google Scholar] [PubMed]
- Park, H.J.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Gallo, R.L.; Cho, D.H. Adrenocorticotropin hormone stimulates interleukin-18 expression in human HaCaT keratinocytes. J. Investig. Dermatol. 2007, 127, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Cho, D.H.; Kim, H.S.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Park, H.J. Expression of the corticotropin-releasing hormone-proopiomelanocortin axis in the various clinical types of psoriasis. Exp. Dermatol. 2007, 16, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, H.J.; Lee, J.H.; Lee, J.Y.; Cho, B.K.; Kang, J.S.; Kang, H.; Yang, Y.; Cho, D.H. Corticotropin-releasing hormone (CRH) downregulates interleukin-18 expression in human HaCaT keratinocytes by activation of p38 mitogen-activated protein kinase (MAPK) pathway. J. Investig. Dermatol. 2005, 124, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Yu, X.; Cai, D.; Liu, C.; Li, C. Role of corticotropin-releasing hormone and receptor in the pathogenesis of psoriasis. Med. Hypotheses 2009, 73, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.; Doble, R.; Bachmann, M.; Pfeilschifter, J.; Werfel, T.; Muhl, H. IL-27 Regulates IL-18 binding protein in skin resident cells. PLoS ONE 2012, 7, e38751. [Google Scholar] [CrossRef] [PubMed]
- Bieber, T. Atopic dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Eichenfield, L.F.; Tom, W.L.; Chamlin, S.L.; Feldman, S.R.; Hanifin, J.M.; Simpson, E.L.; Berger, T.G.; Bergman, J.N.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 1. Diagnosis and assessment of atopic dermatitis. J. Am. Acad. Dermatol. 2014, 70, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Novak, N.; Valenta, R.; Bohle, B.; Laffer, S.; Haberstok, J.; Kraft, S.; Bieber, T. FcεRI engagement of Langerhans cell-like dendritic cells and inflammatory dendritic epidermal cell-like dendritic cells induces chemotactic signals and different T-cell phenotypes in vitro. J. Allergy Clin. Immunol. 2004, 113, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tsutsui, H.; Yoshimoto, T.; Kotani, M.; Matsumoto, M.; Fujita, A.; Wang, W.; Higa, S.; Koshimoto, T.; Nakanishi, K.; et al. Interleukin-18 is elevated in the sera from patients with atopic dermatitis and from atopic dermatitis model mice, NC/Nga. Int. Arch. Allergy Immunol. 2001, 125, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Tsutsui, H.; Murakami, T.; Yumikura-Futatsugi, S.; Yamanaka, K.; Tanaka, M.; Iwakura, Y.; Suzuki, N.; Takeda, K.; Akira, S.; et al. IL-18 contributes to the spontaneous development of atopic dermatitis-like inflammatory skin lesion independently of IgE/stat6 under specific pathogen-free conditions. Proc. Natl. Acad. Sci. USA 2002, 99, 11340–11345. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, T.; Tsutsui, H.; Tominaga, K.; Hoshino, K.; Okamura, H.; Akira, S.; Paul, W.E.; Nakanishi, K. IL-18, although antiallergic when administered with IL-12, stimulates IL-4 and histamine release by basophils. Proc. Natl. Acad. Sci. USA 1999, 96, 13962–13966. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Yagita, H.; Ortaldo, J.R.; Wiltrout, R.H.; Young, H.A. In vivo administration of IL-18 can induce IgE production through Th2 cytokine induction and up-regulation of CD40 ligand (CD154) expression on CD4+ T cells. Eur. J. Immunol. 2000, 30, 1998–2006. [Google Scholar] [CrossRef]
- Tsutsui, H.; Yoshimoto, T.; Hayashi, N.; Mizutani, H.; Nakanishi, K. Induction of allergic inflammation by interleukin-18 in experimental animal models. Immunol. Rev. 2004, 202, 115–138. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Aihara, M.; Kirino, M.; Harada, I.; Komori-Yamaguchi, J.; Yamaguchi, Y.; Nagashima, Y.; Ikezawa, Z. Interleukin-18 is elevated in the horny layer in patients with atopic dermatitis and is associated with Staphylococcus aureus colonization. Br. J. Dermatol. 2011, 164, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Terada, M.; Tsutsui, H.; Imai, Y.; Yasuda, K.; Mizutani, H.; Yamanishi, K.; Kubo, M.; Matsui, K.; Sano, H.; Nakanishi, K. Contribution of IL-18 to atopic-dermatitis-like skin inflammation induced by Staphylococcus aureus product in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 8816–8821. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.L.; Sato, M.N.; Takaoka, R.; Azor, M.H.; Rivitti, E.A.; Hanifin, J.M.; Aoki, V. Atopic dermatitis in adults: Evaluation of peripheral blood mononuclear cells proliferation response to Staphylococcus aureus enterotoxins A and B and analysis of interleukin-18 secretion. Exp. Dermatol. 2009, 18, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Choi, J.K.; Jung, H.J.; Park, K.H.; Jang, Y.H.; Lee, W.J.; Lee, S.J.; Kim, S.H.; Kang, H.Y.; Kim, J.M.; et al. Effects of topical application of a recombinant staphylococcal enterotoxin A on DNCB and dust mite extract-induced atopic dermatitis-like lesions in a murine model. Eur. J. Dermatol. 2014, 24, 186–193. [Google Scholar] [PubMed]
- Buentke, E.; Heffler, L.C.; Wallin, R.P.; Lofman, C.; Ljunggren, H.G.; Scheynius, A. The allergenic yeast Malassezia furfur induces maturation of human dendritic cells. Clin. Exp. Allergy 2001, 31, 1583–1593. [Google Scholar] [CrossRef] [PubMed]
- Zedan, K.; Rasheed, Z.; Farouk, Y.; Alzolibani, A.A.; Saif, G.B.; Ismail, H.A.; al Robaee, A.A. Immunoglobulin E, interleukin-18 and interleukin-12 in patients with atopic dermatitis: Correlation with disease activity. J. Clin. Diagn. Res. 2015, 9, WC01-5. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; McEwen, B.S. Enhancing versus suppressive effects of stress hormones on skin immune function. Proc. Natl. Acad. Sci. USA 1999, 96, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kwon, Y.S.; Park, C.O.; Oh, S.H.; Lee, J.H.; Wu, W.H.; Chang, N.S.; Lee, M.G.; Lee, K.H. Corticotropin-releasing factor decreases IL-18 in the monocyte-derived dendritic cell. Exp. Dermatol. 2009, 18, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Hao, Y.; Zhou, W.; Ma, Y. The relationship between interleukin-18 polymorphisms and allergic disease: A meta-analysis. Biomed. Res. Int. 2014, 2014, 290687. [Google Scholar] [CrossRef] [PubMed]
- Ring, J.; Brockow, K.; Ollert, M.; Engst, R. Antihistamines in urticaria. Clin. Exp. Allergy 1999, 29, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, A.; Lorini, M.; Suli, C.; Asero, R. Serum interleukin-18 in patients with chronic ordinary urticaria: Association with disease activity. Clin. Exp. Dermatol. 2007, 32, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Puxeddu, I.; Italiani, P.; Giungato, P.; Pratesi, F.; Panza, F.; Bartaloni, D.; Rocchi, V.; del Corso, I.; Boraschi, D.; Migliorini, P. Free IL-18 and IL-33 cytokines in chronic spontaneous urticaria. Cytokine 2013, 61, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Rasool, R.; Ashiq, I.; Shera, I.A.; Yousuf, Q.; Shah, Z.A. Study of serum interleukin (IL) 18 and IL-6 levels in relation with the clinical disease severity in chronic idiopathic urticaria patients of Kashmir (North India). Asia Pac. Allergy 2014, 4, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Wiener, Z.; Pocza, P.; Racz, M.; Nagy, G.; Tolgyesi, G.; Molnar, V.; Jaeger, J.; Buzas, E.; Gorbe, E.; Papp, Z.; et al. IL-18 induces a marked gene expression profile change and increased Ccl1 (I-309) production in mouse mucosal mast cell homologs. Int. Immunol. 2008, 20, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, P.; del Corso, I.; Tommasi, C.; Boraschi, D. Free circulating interleukin-18 is increased in Schnitzler syndrome: A new autoinflammatory disease? Eur. Cytokine Netw. 2009, 20, 108–111. [Google Scholar] [PubMed]
- Bourke, J.; Coulson, I.; English, J.; British Association of Dermatologists Therapy, G.; Audit, S. Guidelines for the management of contact dermatitis: An update. Br. J. Dermatol. 2009, 160, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Worm, M. Allergic contact dermatitis beyond IL-1β role of additional family members. Exp. Dermatol. 2014, 23, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, V.; Papale, A.; Galli, C.L.; Marinovich, M.; Corsini, E. Role of ROS and HMGB1 in contact allergen-induced IL-18 production in human keratinocytes. J. Investig. Dermatol. 2014, 134, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- McFadden, J.P.; Puangpet, P.; Basketter, D.A.; Dearman, R.J.; Kimber, I. Why does allergic contact dermatitis exist? Br. J. Dermatol. 2013, 168, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Gaide, O.; Petrilli, V.; Martinon, F.; Contassot, E.; Roques, S.; Kummer, J.A.; Tschopp, J.; French, L.E. Activation of the IL-1β-processing inflammasome is involved in contact hypersensitivity. J. Investig. Dermatol. 2007, 127, 1956–1963. [Google Scholar] [CrossRef] [PubMed]
- Kambe, N.; Nakamura, Y.; Saito, M.; Nishikomori, R. The inflammasome, an innate immunity guardian, participates in skin urticarial reactions and contact hypersensitivity. Allergol. Int. 2010, 59, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, C.; Cumberbatch, M.; Mee, J.B.; Dearman, R.J.; Wei, X.Q.; Liew, F.Y.; Kimber, I.; Groves, R.W. IL-18 is a key proximal mediator of contact hypersensitivity and allergen-induced Langerhans cell migration in murine epidermis. J. Leukoc. Biol. 2008, 83, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Klekotka, P.A.; Yang, L.; Yokoyama, W.M. Contrasting roles of the IL-1 and IL-18 receptors in MyD88-dependent contact hypersensitivity. J. Investig. Dermatol. 2010, 130, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Alkhalifah, A.; Alsantali, A.; Wang, E.; McElwee, K.J.; Shapiro, J. Alopecia areata update: Part I. Clinical picture, histopathology, and pathogenesis. J. Am. Acad. Dermatol. 2010, 62, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Nickoloff, B.J.; Ito, T. A ‘hairy’ privilege. Trends Immunol. 2005, 26, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Gilhar, A.; Kam, Y.; Assy, B.; Kalish, R.S. Alopecia areata induced in C3H/HeJ mice by interferon-γ: Evidence for loss of immune privilege. J. Investig. Dermatol. 2005, 124, 288–289. [Google Scholar] [CrossRef] [PubMed]
- Manolache, L.; Benea, V. Stress in patients with alopecia areata and vitiligo. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Cho, D.H.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Park, H.J. Immunoreactivity of corticotropin-releasing hormone, adrenocorticotropic hormone and α-melanocyte-stimulating hormone in alopecia areata. Exp. Dermatol. 2006, 15, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Hong, S.K.; Park, S.W.; Hur, D.Y.; Shon, J.H.; Shin, J.G.; Hwang, S.W.; Sung, H.S. Serum levels of IL-18 and sIL-2R in patients with alopecia areata receiving combined therapy with oral cyclosporine and steroids. Exp. Dermatol. 2010, 19, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Park, H.J.; Chung, J.H.; Kim, J.W.; Seok, H.; Lew, B.L.; Sim, W.Y. Association between interleukin 18 polymorphisms and alopecia areata in Koreans. J. Interferon Cytokine Res. 2014, 34, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Roujeau, J.C.; Stern, R.S. Severe adverse cutaneous reactions to drugs. N. Engl. J. Med. 1994, 331, 1272–1285. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, H.S.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Lee, A.W.; Yoon, D.Y.; Cho, D.H. Immunohistochemical characterization of cutaneous drug eruptions by STI571. J. Dermatol. Sci. 2005, 38, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Ye, Y.M.; Hur, G.Y.; Shin, S.Y.; Han, J.H.; Park, H.S. Neutrophil activation in patients with ASA-induced urticaria. J. Clin. Immunol. 2008, 28, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Son, J.K.; Yang, E.M.; Kim, J.E.; Park, H.S. A functional promoter polymorphism of the human IL18 gene is associated with aspirin-induced urticaria. Br. J. Dermatol. 2011, 165, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, J.N.; Sontheimer, R.D. Distinctive cutaneous subsets in the spectrum of lupus erythematosus. J. Am. Acad. Dermatol. 1981, 4, 471–475. [Google Scholar] [CrossRef]
- Walling, H.W.; Sontheimer, R.D. Cutaneous lupus erythematosus: Issues in diagnosis and treatment. Am. J. Clin. Dermatol. 2009, 10, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, P.; Anzilotti, C.; Pratesi, F.; Quattroni, P.; Bargagna, M.; Dinarello, C.A.; Boraschi, D. Serum and urinary levels of IL-18 and its inhibitor IL-18BP in systemic lupus erythematosus. Eur. Cytokine Netw. 2010, 21, 264–271. [Google Scholar] [PubMed]
- Kinoshita, K.; Yamagata, T.; Nozaki, Y.; Sugiyama, M.; Ikoma, S.; Funauchi, M.; Kanamaru, A. Blockade of IL-18 receptor signaling delays the onset of autoimmune disease in MRL-Faslpr mice. J. Immunol. 2004, 173, 5312–5318. [Google Scholar] [CrossRef] [PubMed]
- Maczynska, I.; Millo, B.; Ratajczak-Stefanska, V.; Maleszka, R.; Szych, Z.; Kurpisz, M.; Giedrys-Kalemba, S. Proinflammatory cytokine (IL-1β, IL-6, IL-12, IL-18 and TNF-α) levels in sera of patients with subacute cutaneous lupus erythematosus (SCLE). Immunol. Lett. 2006, 102, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.S.; Werth, V.P. The role of cytokines in the pathogenesis of cutaneous lupus erythematosus. Cytokine 2015, 73, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Kahlenberg, J.M.; Kaplan, M.J. The inflammasome and lupus: Another innate immune mechanism contributing to disease pathogenesis? Curr. Opin. Rheumatol. 2014, 26, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.A.; Chiang, B.L. Inflammasomes and human autoimmunity: A comprehensive review. J. Autoimmun. 2015, 61, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Wan, L.; Lee, C.C.; Huang, C.M.; Tsai, Y.; Tsai, C.H.; Shin, T.L.; Chao, K.; Liu, C.M.; Xiao, J.W.; Tsai, F.J. Disease association of the interleukin-18 promoter polymorphisms in Taiwan Chinese systemic lupus erythematosus patients. Genes Immun. 2007, 8, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Y.; Hsieh, C.W.; Chen, K.S.; Chen, Y.M.; Lin, F.J.; Lan, J.L. Association of interleukin-18 promoter polymorphisms with WHO pathological classes and serum IL-18 levels in Chinese patients with lupus nephritis. Lupus 2009, 18, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Choi, J.M.; Kim, J.E.; Cho, B.K.; Kim, D.W.; Park, H.J. Clinicopathologic characteristics of cutaneous chronic graft-versus-host diseases: A retrospective study in Korean patients. Int. J. Dermatol. 2010, 49, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Teshima, T.; Kukuruga, M.; Ordemann, R.; Liu, C.; Lowler, K.; Ferrara, J.L. Interleukin-18 regulates acute graft-versus-host disease by enhancing Fas-mediated donor T cell apoptosis. J. Exp. Med. 2001, 194, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, Y.; Takatsuka, H.; Takemoto, Y.; Hara, H.; Okamura, H.; Nakanishi, K.; Kakishita, E. Elevated interleukin (IL)-18 levels during acute graft-versus-host disease after allogeneic bone marrow transplantation. Br. J. Haematol. 2000, 109, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, C.; Chen, W.; Pan, B.; Kong, F.; Zheng, K.; Tang, R.; Zeng, L. Protective effect of neutralizing anti-IL-18α monoclonal antibody on a mouse model of acute graft-versus-host disease. Oncol. Rep. 2015, 34, 2031–2039. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T. Cutaneous manifestations associated with adult-onset Still's disease: Important diagnostic values. Rheumatol. Int. 2012, 32, 2233–2237. [Google Scholar] [CrossRef] [PubMed]
- Janssen, R.; Verhard, E.; Lankester, A.; Ten Cate, R.; van Dissel, J.T. Enhanced interleukin-1β and interleukin-18 release in a patient with chronic infantile neurologic, cutaneous, articular syndrome. Arthritis Rheum. 2004, 50, 3329–3333. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, H.; Kalina, U.; Muhl, H.; Pfeilschifter, J.; Frank, S. Counterregulation of interleukin-18 mRNA and protein expression during cutaneous wound repair in mice. J. Investig. Dermatol. 1999, 113, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Sabuncu, T.; Eren, M.A.; Tabur, S.; Dag, O.F.; Boduroglu, O. High serum concentration of interleukin-18 in diabetic patients with foot ulcers. J. Am. Podiatr. Med. Assoc. 2014, 104, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Do, D.V.; Ong, C.T.; Khoo, Y.T.; Carbone, A.; Lim, C.P.; Wang, S.; Mukhopadhyay, A.; Cao, X.; Cho, D.H.; Wei, X.Q.; et al. Interleukin-18 system plays an important role in keloid pathogenesis via epithelial-mesenchymal interactions. Br. J. Dermatol. 2012, 166, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Scala, E.; Pallotta, S.; Frezzolini, A.; Abeni, D.; Barbieri, C.; Sampogna, F.; de Pita, O.; Puddu, P.; Paganelli, R.; Russo, G. Cytokine and chemokine levels in systemic sclerosis: Relationship with cutaneous and internal organ involvement. Clin. Exp. Immunol. 2004, 138, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.F.; Wang, J.; Leng, R.X.; Li, X.P.; Ye, D.Q. Interleukin-18: Friend or foe for systemic sclerosis? J. Investig. Dermatol. 2011, 131, 2495–2495. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Song, S.B.; Choi, J.M.; Kim, K.M.; Cho, B.K.; Cho, D.H.; Park, H.J. IL-18 downregulates collagen production in human dermal fibroblasts via the ERK pathway. J. Investig. Dermatol. 2010, 130, 706–715. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Cho, D.H.; Park, H.J. IL-18 and Cutaneous Inflammatory Diseases. Int. J. Mol. Sci. 2015, 16, 29357-29369. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226172
Lee JH, Cho DH, Park HJ. IL-18 and Cutaneous Inflammatory Diseases. International Journal of Molecular Sciences. 2015; 16(12):29357-29369. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226172
Chicago/Turabian StyleLee, Ji Hyun, Dae Ho Cho, and Hyun Jeong Park. 2015. "IL-18 and Cutaneous Inflammatory Diseases" International Journal of Molecular Sciences 16, no. 12: 29357-29369. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226172