
Effects of Atmospheric Pressure Plasmas on Isolated and Cellular DNA—A Review


Abstract
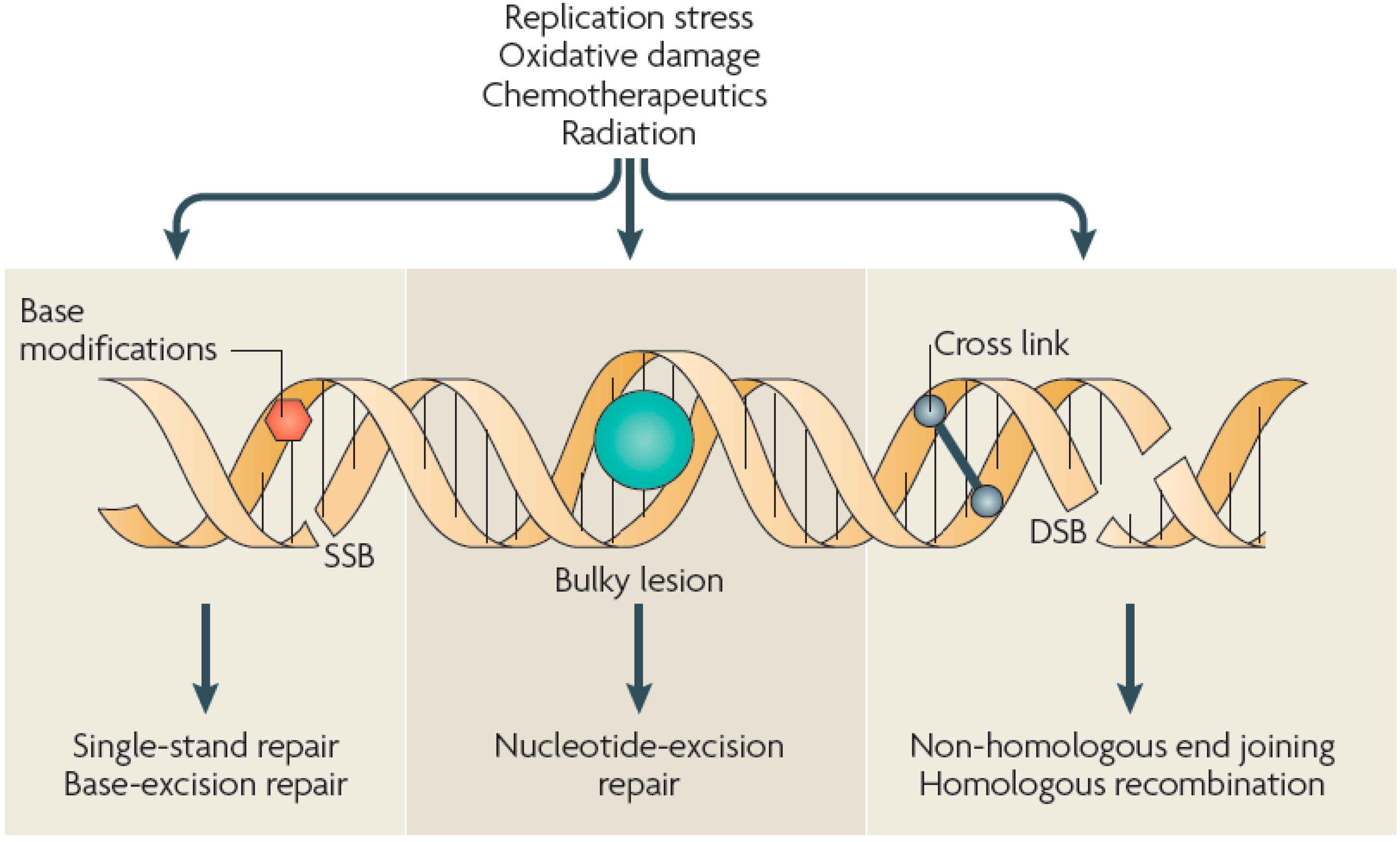
:1. Introduction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
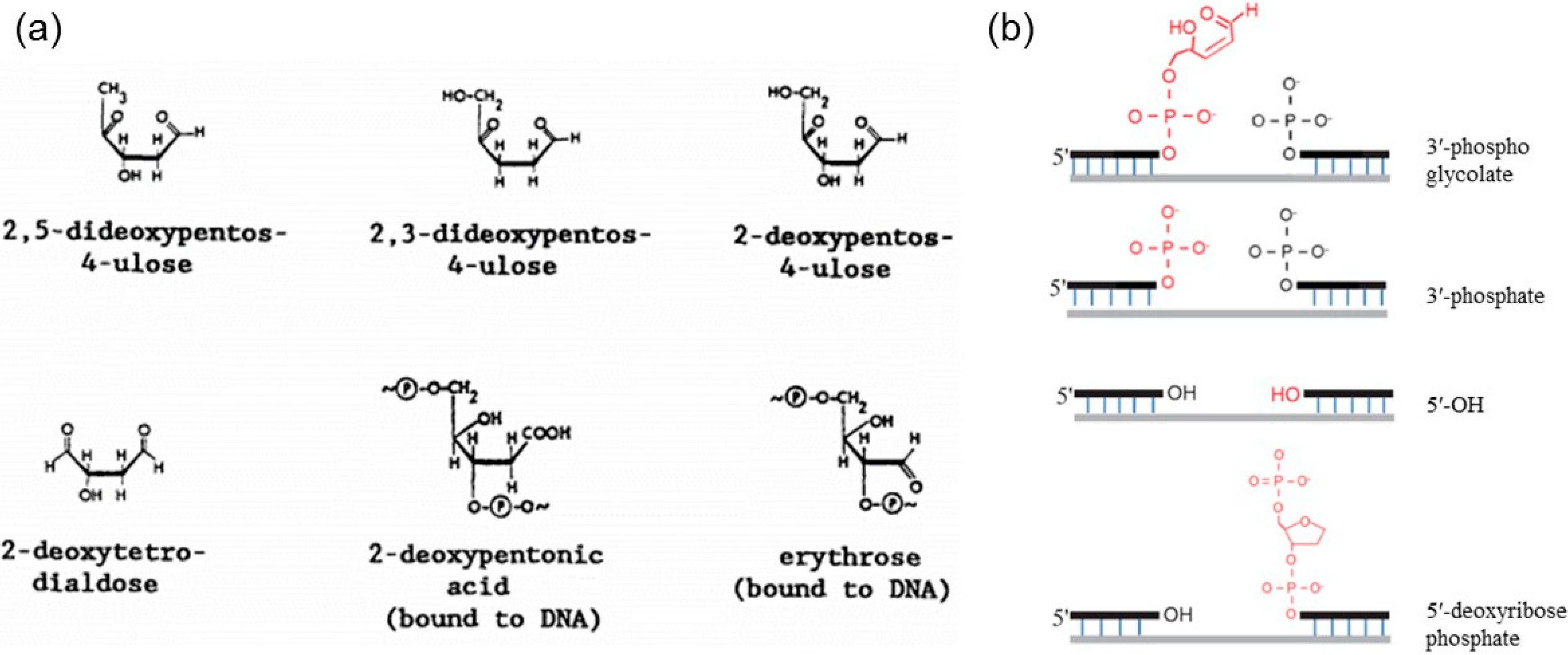{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
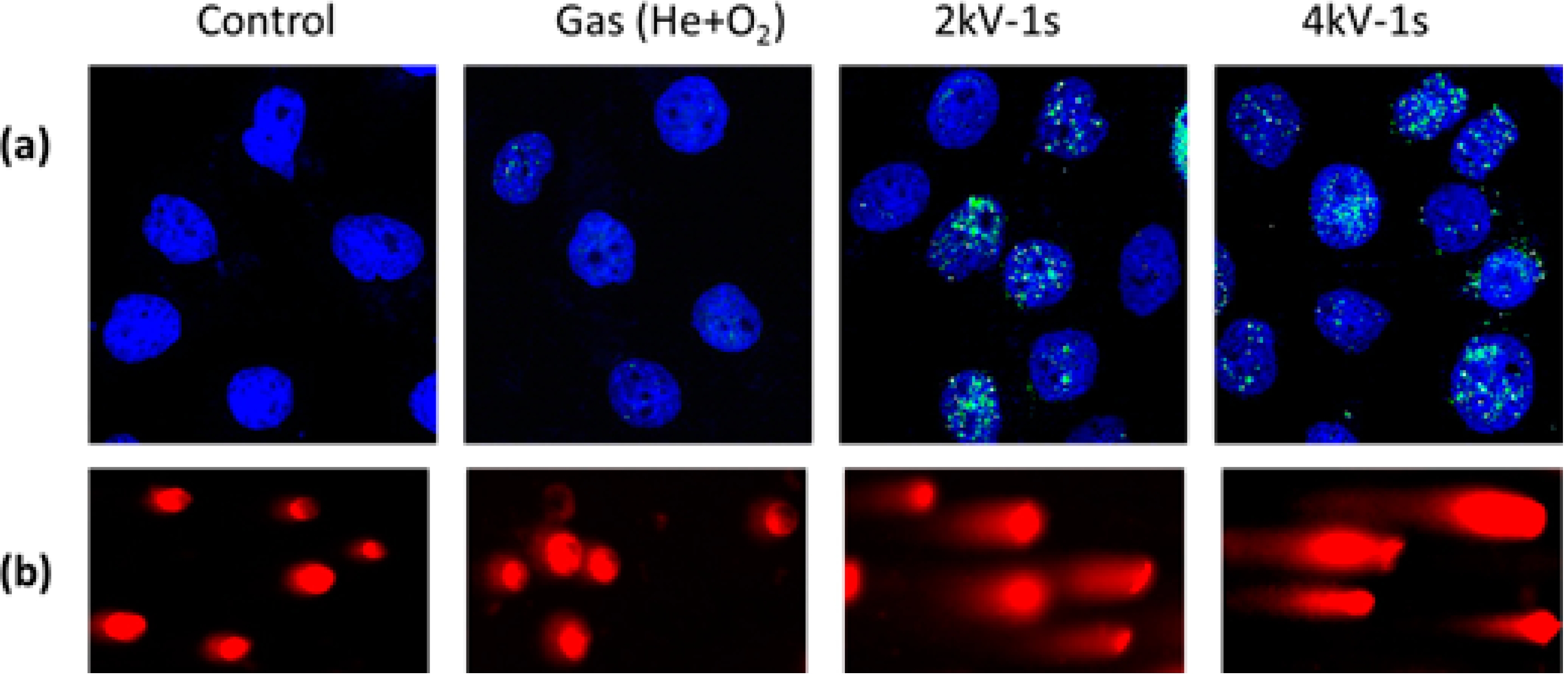{kind=link}
{kind=link}
| Author | Year | Type of APP | Feed Gas | Reference |
|---|---|---|---|---|
| Leduc et al. | 2009, 2010 | Plasma jet | He | [29,46] |
| Alkawareek et al. | 2014 | Plasma jet | 99.5% He/0.5% O2 | [45] |
| Bahnev et al. | 2014 | Plasma jet | He | [47] |
| Han et al. | 2014 | Plasma jet | He | [12] |
| Hosseinzadeh Colagar et al. | 2013 | Plasma jet | 99% Ar/1% Air | [48] |
| Kim et al. | 2006 | Plasma jet | He/O2 | [49] |
| Li et al. | 2008 | Plasma jet | He | [50] |
| Niemi et al. | 2012 | Plasma jet | He/O2 | [51] |
| O’Connell et al. | 2011 | Plasma jet | He/0.5% O2 | [52] |
| Ptasinska et al. | 2010 | Plasma jet | He | [53] |
| Stypczyńska et al. | 2010 | Plasma jet | He | [54] |
| Yan et al. | 2009 | Plasma jet | He/1% O2 | [55] |
| Young Kim et al. | 2012 | Plasma jet | He/O2 | [56] |
| Lackmann et al. | 2012, 2013 | Plasma jet | He/0.6% O2 | [57,58] |
| Antoniu et al. | 2012 | Plasma jet | He | [59] |
| Kurita et al. | 2011, 2014 | Plasma jet | Ar | [60,61] |
| Sousa et al. | 2010, 2012 | Micro cathode sustained discharge (MCSD) array | He/O2/NO | [62,63] |
| Blackert et al. | 2013 | DBD | Ambient air | [8] |
| Brun et al. | 2011 | Plasma jet | He | [64] |
| Chang et al. | 2014 | Plasma jet “torch with spray type” | He/O2 | [65] |
| Han et al. | 2013, 2014 | Plasma jet | N2 | [13,66] |
| Isbary et al. | 2013 | SMD | Ambient air | [15] |
| Kalghatgi et al. | 2010, 2011, 2012 | DBD | Ambient air | [7,67,68] |
| Kim et al. | 2010 | Surface type APP | Ambient air | [69] |
| Kim et al. | 2011 | Plasma jet with micronozzle array | N2 | [70] |
| Ma et al. | 2014 | DBD | He | [71] |
| Morales-Ramirez et al. | 2013 | Plasma needle | He | [72] |
| Plewa et al. | 2014 | Plasma jet | He | [73] |
| Vandamme et al. | 2011 | FE-DBD | Ambient air | [33] |
| Volotskova et al. | 2012 | Plasma jet | He | [74] |
| Wende et al. | 2014 | Plasma jet | Ar | [14] |
| Yan et al. | 2010 | Plasma jet | He | [75] |
| Choi et al. | 2012 | Microwave plasma | Ar | [76] |
| Lazovic et al. | 2014 | Plasma needle | He | [77] |
| Wu et al. | 2013 | FE-DBD | Ambient air | [9] |
| Lee et al. | 2014 | Plasma jet | N2/air | [78] |
| Ryu et al. | 2013 | Plasma jet | Ar | [79] |
| Joshi et al. | 2011 | DBD | Ambient air | [80] |
| Kvam et al. | 2012 | FE-DBD | Ambient air | [81] |
| Tseng et al. | 2012 | Plasma jet | He/N2 | [82] |
| Mols et al. | 2013 | Plasma jet | N2 | [83] |
| Sharma et al. | 2009 | Plasma jet | Ar | [84] |
| Winter et al. | 2011 | DBD | Ar | [85] |
| Lu et al. | 2014 | DBD | Air, 90% N2/10% O2, and 65% O2/30% CO2/5% N2 | [86] |
| Venezia et al. | 2008 | Plasma glow | 1% ethylene/50% O2/49% N2 | [87] |
| Yasuda et al. | 2010 | DBD | Ambient air | [88] |
| Wang et al. | 2010 | Plasma jet | He | [89] |
| Fang et al. | 2013 | Plasma jet | He | [90] |
2. Reactive Species Involved in DNA Damage
| Free Radicals | Non-Radicals |
|---|---|
| Reactive Oxygen Species (ROS) | |
| Superoxide, O2●− | Hydrogen peroxide, H2O2 |
| Hydroxyl, ●OH | Ozone, O3 |
| Hydroperoxyl, HO2● (protonated superoxide) | Singlet, 1O2 |
| Carbonate, CO3●− | Organic peroxides, ROOH |
| Alkoxyl, RO● | Peroxynitrite, ONOO− |
| Peroxyl, RO2● | Nitrosoperoxycarbonate, ONOOCO2 |
| Carbon dioxide radical, CO2●− | |
| Reactive Nitrogen Species (RNS) | |
| Nitric oxide, ●NO | Nitrous acid, HNO2 |
| Nitrogen dioxide, ●NO2 | Peroxynitrite, ONOO− |
| Peroxynitrous acid, ONOOH | |
| Alkyl peroxynitrites, ROONO | |
| Alkyl peroxynitrates, RO2ONO | |
| Species | Reaction | E (V/NHE) |
|---|---|---|
| Hydroxyl Radical | ●OH + H+ + e− ⇌ H2O | 2.80 |
| ●OH + e− ⇌ OH− | 1.89 | |
| Ozone | O3 + 2 H+ + 2 e− ⇌ O2 + H2O | 2.08 |
| O3 + H2O + 2 e− ⇌ O2 + 2 OH− | 1.24 | |
| Hydrogen peroxide | H2O2 + 2 H+ + 2 e− ⇌ 2 H2O | 1.78 |
| H2O2 + 2 e− ⇌ 2 OH− | 0.88 | |
| Singlet Oxygen | 1O2 + 4 H+ + 4 e− ⇌ 2 H2O | 1.79 (pH 7.0) |
| Carbonate Radical | CO3●− + e− ⇌ CO32− | 1.59 |
| Dissolved Oxygen | O2 + 4 H+ + 4 e− ⇌ 2 H2O | 1.23 |
| O2 + 2 H2O + 4 e− ⇌ 4 OH− | 0.40 | |
| Nitrogen Dioxide Radicals | NO2● + e− ⇌ NO2− | 1.04 |
| Superoxide | O2 + H+ + e− ⇌ HO2 | −0.05 |
| O2 + e− ⇌ O2●− | −0.33 |
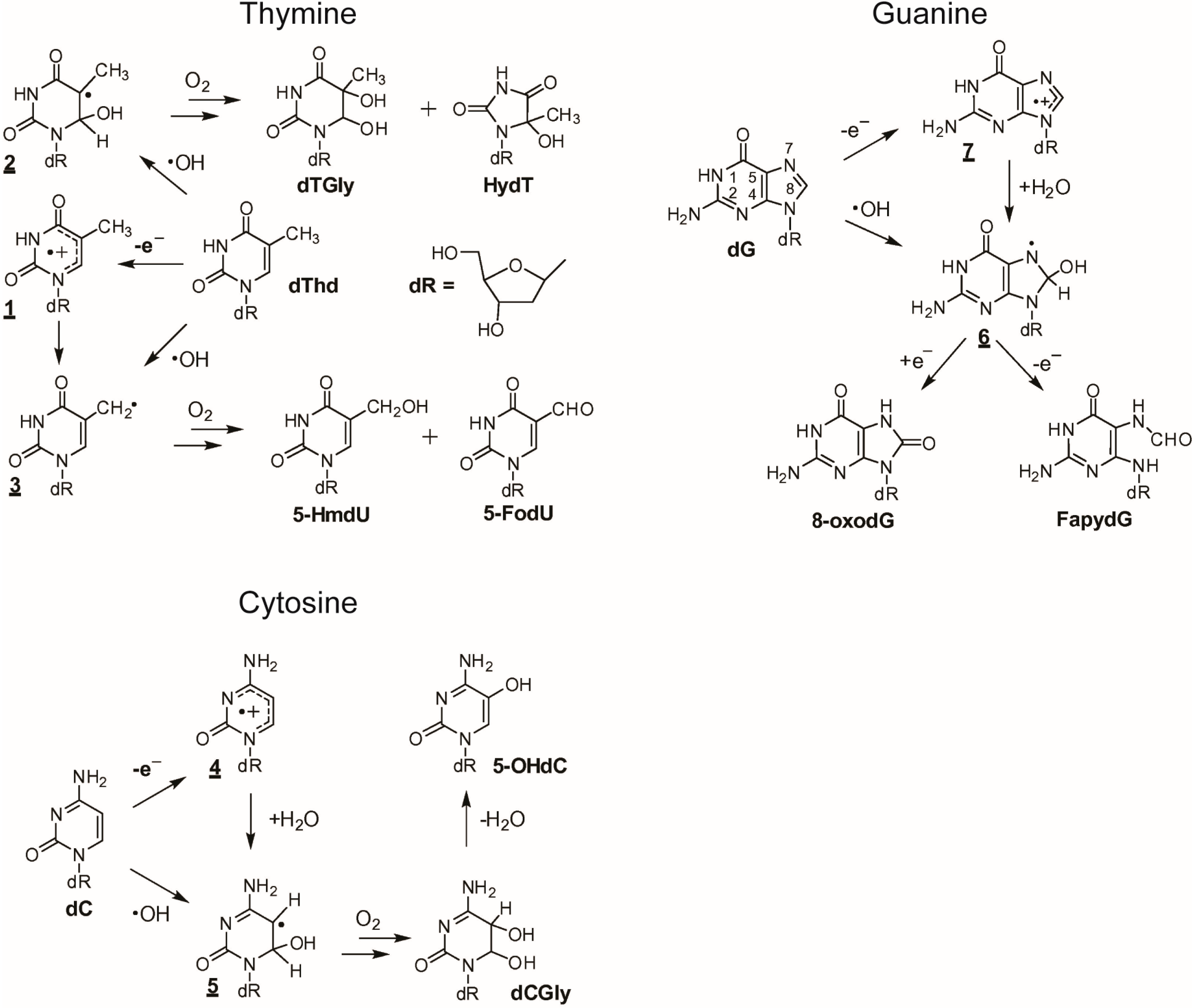
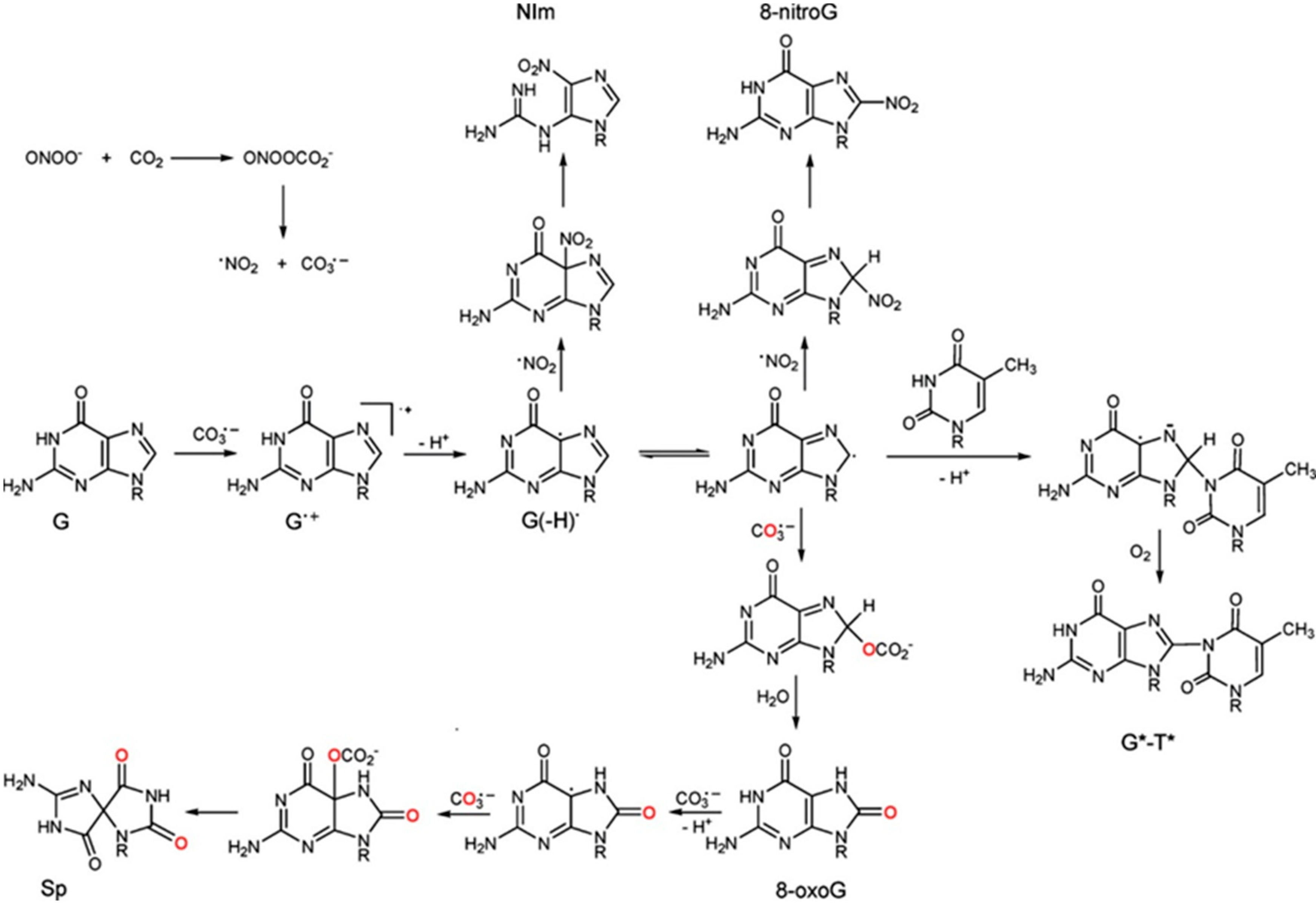
2.1. Reactivity of ROS towards Nucleobases
| Substrate | pKa | ●OH 1 | Major Products of Residues |
|---|---|---|---|
| Adenine | 4.15, 9.8 | 6.1 × 109 | dR-derivatives of 4,6-diamino-5-formamidopyrimidine, 8-oxo-7,8-dihydroadenine and, in some cases, 2-hydroxy-adenine also forms. |
| Cytosine | 4.6, 12.2 | 6.1 × 109 | dR-derivatives of 5,6-dihydroxy-5,6-dihydrocytosine, 5-hydroxycytosine,6-hydroxy-5,6-dihydrocytosine, 5,6-dihydroxy-5,6-dihydro-uracil, 5-hydroxyuracil, 5-hydroxyhydantoin, trans-1-car-bamoyl-4,5-dihydroxyimidazolidin-2-one and dR-isodialuric acid. |
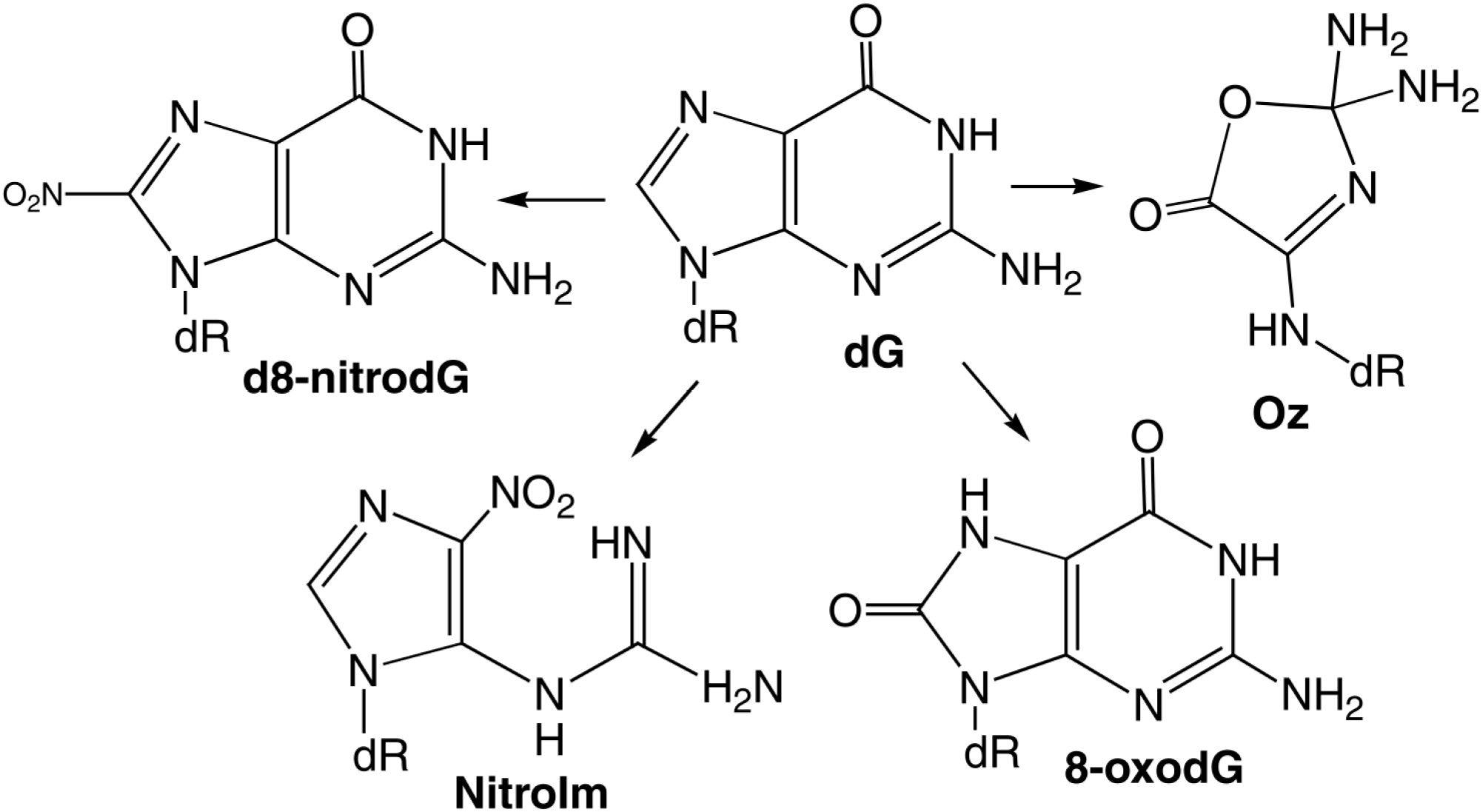
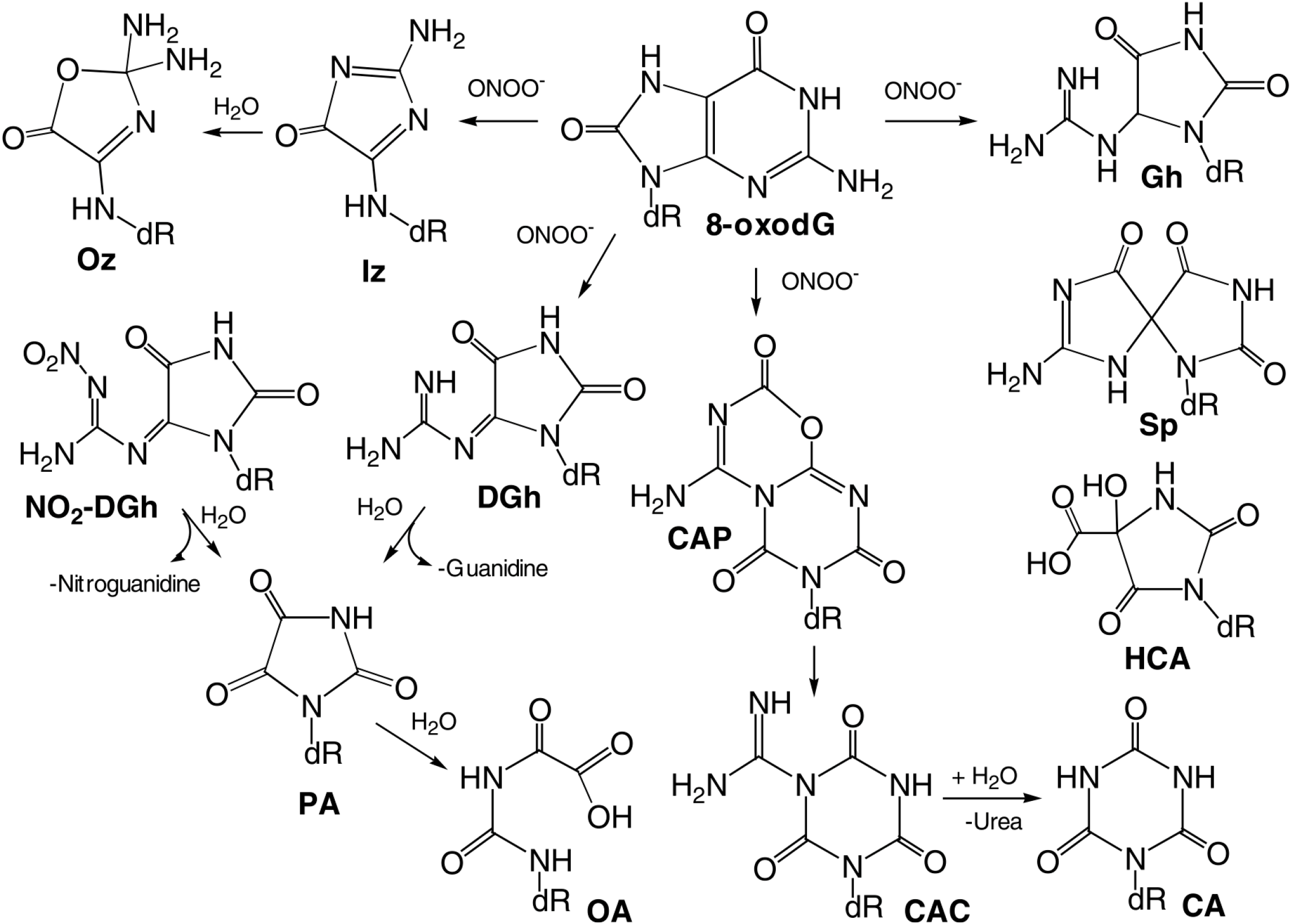
| Guanine | 3.2, 9.8 | 9.2 × 109 | dR-derivatives of imidazolone, oxazolone, 8-oxo-7,8-dihydroguanine and 2,6-di-amino-4-hydroxy-5-formamidopyrimidine. In addition, 1,N(2)-adducts of glyoxal and propene to guanine can be formed. |
| Thymine | 9.9, >13 | 6.4 × 109 | dR-derivatives of 5,6-dihydroxy-5,6-dihydrocytosine, 5-hydroxycytosine, 6-hydroxy-5,6-dihydrocytosine, 5,6-dihydroxy-5,6-dihydro-uracil, 5-hydroxyuracil, 5-hydroxyhydantoin, trans-1-car-bamoyl-4,5-dihydroxyimidazolidin-2-one and dR-isodialuric acid. |
| DNA | - | 4.0 × 108 |

2.2. Reactivity of RNS towards Nucleobases



2.3. DNA Strand Breaks Induced by ROS/RNS

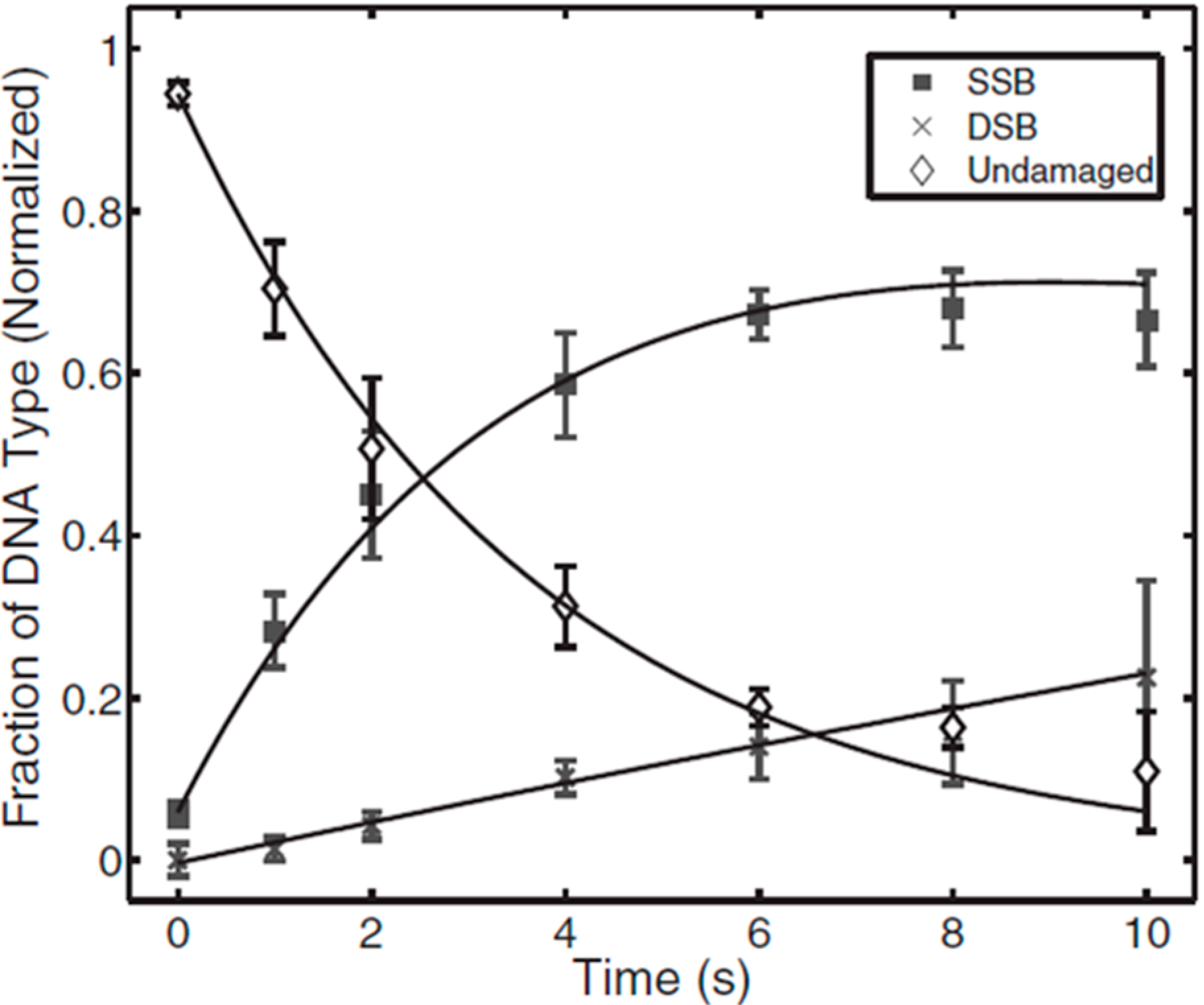
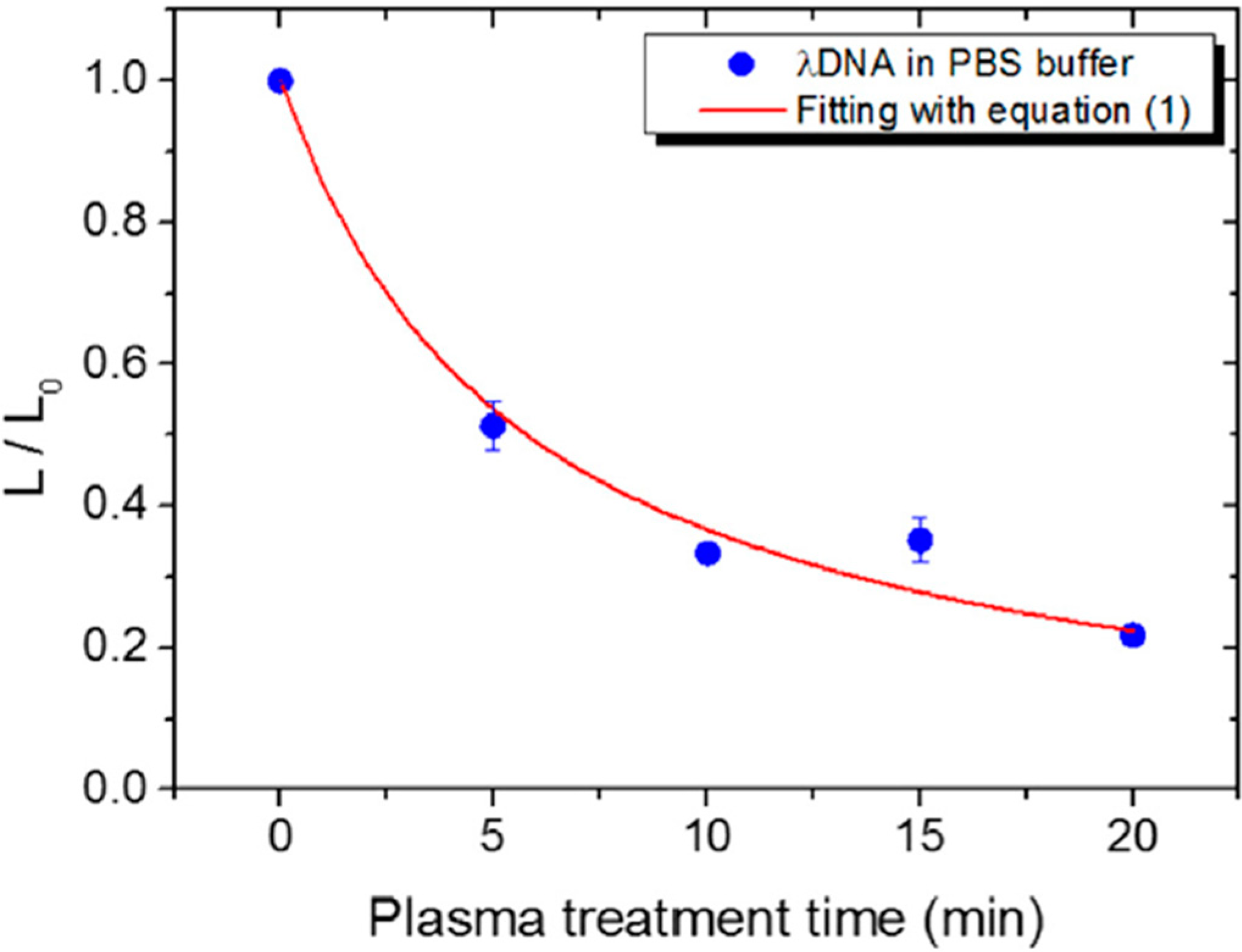
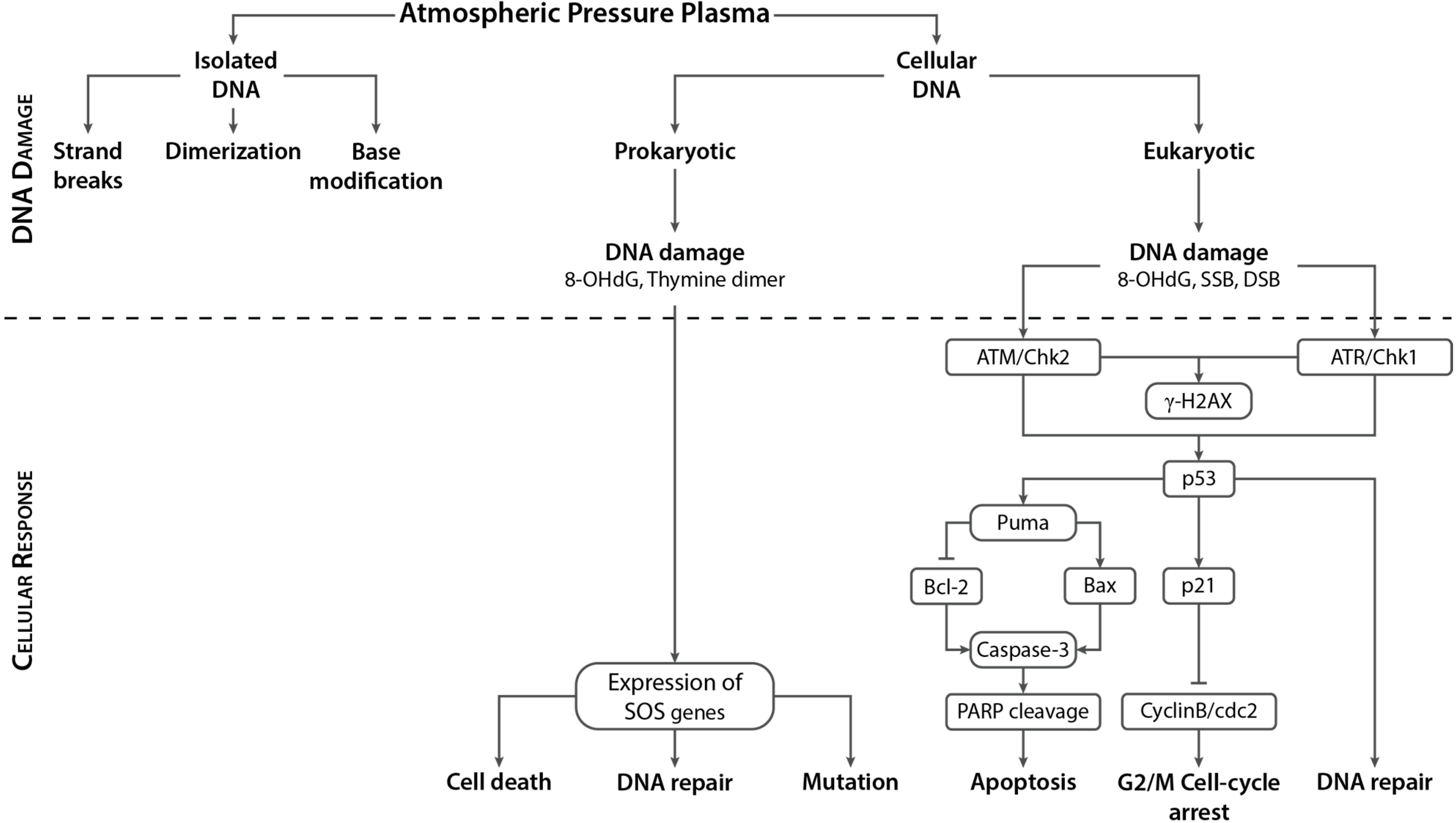
3. APP Interactions with Isolated DNA
3.1. Types of DNA Damage Induced by APPs in Isolated DNA




3.2. Evaluation of Effects of APP Components on Strand Break Formation
4. APP Interactions with Cellular DNA
4.1. Cellular Responses to DNA Damage in Eukaryotic (Mammalian) Systems
4.1.1. DNA Damage Response (DDR) and Cell Cycle Checkpoints
4.1.2. Dual Function of Tumor Suppressor p53
4.1.3. DNA Damage Repair Mechanisms in Response to Oxidative Stress

4.2. APP-Induced DNA Damage in Eukaryotic Cells and Associated Cellular Responses


4.3. APP-induced DNA Damage in Prokaryotic Cells and Associated Response
5. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Von Woedtke, T.; Reuter, S.; Masur, K.; Weltmann, K.D. Plasmas for medicine. Phys. Rep. 2013, 530, 291–320. [Google Scholar] [CrossRef]
- Xiong, Q.; Lu, X.; Jiang, Z.H.; Tang, Z.Y.; Hu, J.; Xiong, Z.L.; Pan, Y. An atmospheric pressure nonequilibrium plasma jet device. Plasma Sci. IEEE Trans. 2008, 36, 986–987. [Google Scholar] [CrossRef]
- Graves, D.B. The emerging role of reactive oxygen and nitrogen species in redox biology and some implications for plasma applications to medicine and biology. J. Phys. D 2012, 45. [Google Scholar] [CrossRef]
- Isbary, G.; Shimizu, T.; Li, Y.; Stolz, W.; Thomas, H.M.; Morfill, G.E.; Zimmermann, J.L. Cold atmospheric plasma devices for medical issues. Expert Rev. Med. Devices 2013, 10, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Arjunan, K.P.; Friedman, G.; Fridman, A.; Clyne, A.M. Non-thermal dielectric barrier discharge plasma induces angiogenesis through reactive oxygen species. J. R. Soc. Inter. 2011. [Google Scholar] [CrossRef]
- Fridman, G.; Peddinghaus, M.; Balasubramanian, M.; Ayan, H.; Fridman, A.; Gutsol, A.; Brooks, A. Blood coagulation and living tissue sterilization by floating-electrode dielectric barrier discharge in air. Plasma Chem. Plasma Process. 2006, 26, 425–442. [Google Scholar] [CrossRef]
- Kalghatgi, S.; Kelly, C.M.; Cerchar, E.; Torabi, B.; Alekseev, O.; Fridman, A.; Friedman, G.; Azizkhan-Clifford, J. Effects of non-thermal plasma on mammalian cells. PLoS One 2011, 6, e16270. [Google Scholar] [CrossRef] [PubMed]
- Blackert, S.; Haertel, B.; Wende, K.; von Woedtke, T.; Lindequist, U. Influence of non-thermal atmospheric pressure plasma on cellular structures and processes in human keratinocytes (HaCaT). J. Dermatol. Sci. 2013, 70, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.S.; Kalghatgi, S.; Dobrynin, D.; Sensenig, R.; Cerchar, E.; Podolsky, E.; Dulaimi, E.; Paff, M.; Wasko, K.; Arjunan, K.; et al. Porcine intact and wounded skin responses to atmospheric nonthermal plasma. J. Surg. Res. 2013, 179, e1–e12. [Google Scholar] [CrossRef] [PubMed]
- Kuchenbecker, M.; Bibinov, N.; Kaemlimg, A.; Wandke, D.; Awakowicz, P.; Viöl, W. Characterization of DBD plasma source for biomedical applications. J. Phys. D 2009, 42. [Google Scholar] [CrossRef]
- Fridman, G.; Shereshevsky, A.; Jost, M.M.; Brooks, A.D.; Fridman, A.; Gutsol, A.; Vasilets, V.; Friedman, G. Floating electrode dielectric barrier discharge plasma in air promoting apoptotic behavior in melanoma skin cancer cell lines. Plasma Chem. Plasma Process. 2007, 27, 163–176. [Google Scholar] [CrossRef]
- Han, X.; Cantrell, W.A.; Escobar, E.E.; Ptasinska, S. Plasmid DNA damage induced by helium atmospheric pressure plasma jet. Eur. Phys. J. D 2014, 68, 1–7. [Google Scholar] [CrossRef]
- Han, X.; Klas, M.; Liu, Y.; Sharon, S.M.; Ptasinska, S. DNA damage in oral cancer cells induced by nitrogen atmospheric pressure plasma jets. Appl. Phys. Lett. 2013, 102. [Google Scholar] [CrossRef]
- Wende, K.; Straßenburg, S.; Haertel, B.; Harms, M.; Holtz, S.; Barton, A.; Masur, K.; von Woedtke, T.; Lindequist, U. Atmospheric pressure plasma jet treatment evokes transient oxidative stress in HaCaT keratinocytes and influences cell physiology. Cell Biol. Int. 2014, 38, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Isbary, G.; Köritzer, J.; Mitra, A.; Li, Y.F.; Shimizu, T.; Schroeder, J.; Schlegel, J.; Morfill, G.E.; Stolz, W.; Zimmermann, J.L. Ex vivo human skin experiments for the evaluation of safety of new cold atmospheric plasma devices. Clin. Plasma Med. 2013, 1, 36–44. [Google Scholar] [CrossRef]
- Morfill, G.E.; Shimizu, T.; Steffes, B.; Schmidt, H.U. Nosocomial infections—A new approach towards preventive medicine using plasmas. New J. Phys. 2009, 11. [Google Scholar] [CrossRef]
- Hoffmann, C.; Berganza, C.; Zhang, J. Cold atmospheric plasma: Methods of production and application in dentistry and oncology. Med. Gas Res. 2013, 3. [Google Scholar] [CrossRef]
- Maisch, T.; Shimizu, T.; Isbary, G.; Heinlin, J.; Karrer, S.; Klämpfl, T.G.; Li, Y.; Morfill, G.; Zimmermann, J.L. Contact-free inactivation of candida albicans biofilms by cold atmospheric air plasma. Appl. Environ. Microbiol. 2012, 78, 4242–4247. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, J.L.; Dumler, K.; Shimizu, T.; Morfill, G.E.; Wolf, A.; Boxhammer, V.; Schlegel, J.; Gansbacher, B.; Anton, M. Effects of cold atmospheric plasmas on adenoviruses in solution. J. Phys. D 2011, 44. [Google Scholar] [CrossRef]
- Hähnel, M.; von Woedtke, T.; Weltmann, K.-D. Influence of the air humidity on the reduction of bacillus spores in a defined environment at atmospheric pressure using a dielectric barrier surface discharge. Plasma Process. Polym. 2010, 7, 244–249. [Google Scholar] [CrossRef]
- Klämpfl, T.G.; Isbary, G.; Shimizu, T.; Li, Y.; Zimmermann, J.L.; Stolz, W.; Schlegel, J.; Morfill, G.E.; Schmidt, H. Cold atmospheric air plasma sterilization against spores and other microorganisms of clinical interest. Appl. Environ. Microbiol. 2012, 78, 5077–5082. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.P.; Tarasenko, O.; Chang, J.; Popovic, S.; Chen, C.Y.; Fan, H.W.; Scott, A.; Lahiani, M.; Alusta, P.; Drake, J.D.; et al. Contribution of a portable air plasma torch to rapid blood coagulation as a method of preventing bleeding. New J. Phys. 2009, 11. [Google Scholar] [CrossRef]
- Isbary, G.; Morfill, G.; Schmidt, H.U.; Georgi, M.; Ramrath, K.; Heinlin, J.; Karrer, S.; Landthaler, M.; Shimizu, T.; Steffes, B.; et al. A first prospective randomized controlled trial to decrease bacterial load using cold atmospheric argon plasma on chronic wounds in patients. Br. J. Dermatol. 2010, 163, 78–82. [Google Scholar] [PubMed]
- Isbary, G.; Heinlin, J.; Shimizu, T.; Zimmermann, J.L.; Morfill, G.; Schmidt, H.U.; Monetti, R.; Steffes, B.; Bunk, W.; Li, Y.; et al. Successful and safe use of 2 min cold atmospheric argon plasma in chronic wounds: Results of a randomized controlled trial. Br. J. Dermatol. 2012, 167, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Baxter, H.C.; Campbell, G.A.; Richardson, P.R.; Jones, A.C.; Whittle, I.R.; Casey, M.; Whittaker, A.G.; Baxter, R.L. Surgical instrument decontamination: Efficacy of introducing an argon:oxygen RF gas-plasma cleaning step as part of the cleaning cycle for stainless steel instruments. Plasma Sci. IEEE Trans. 2006, 34, 1337–1344. [Google Scholar] [CrossRef]
- Whittaker, A.G.; Graham, E.M.; Baxter, R.L.; Jones, A.C.; Richardson, P.R.; Meek, G.; Campbell, G.A.; Aitken, A.; Baxter, H.C. Plasma cleaning of dental instruments. J. Hosp. Infect. 2004, 56, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Baxter, H.C.; Campbell, G.A.; Whittaker, A.G.; Jones, A.C.; Aitken, A.; Simpson, A.H.; Casey, M.; Bountiff, L.; Gibbard, L.; Baxter, R.L. Elimination of transmissible spongiform encephalopathy infectivity and decontamination of surgical instruments by using radio-frequency gas-plasma treatment. J. Gen. Virol. 2005, 86, 2393–2399. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.D.; Ayan, H.; Vasilets, V.N.; Fridman, A.; Guceri, S.; Sun, W. Effect of dielectric barrier discharge plasma on the attachment and proliferation of osteoblasts cultured over poly(ε-caprolactone) scaffolds. Plasma Process. Polym. 2008, 5, 58–66. [Google Scholar] [CrossRef]
- Leduc, M.; Guay, D.; Leask, R.L.; Coulombe, S. Cell permeabilization using a non-thermal plasma. New J. Phys. 2009, 11. [Google Scholar] [CrossRef]
- Sakai, Y.; Khajoee, V.; Ogawa, Y.; Kusuhara, K.; Katayama, Y.; Hara, T. A novel transfection method for mammalian cells using gas plasma. J. Biotechnol. 2006, 121, 299–308. [Google Scholar]
- Chang, Y.-T.; Chen, G. Oral bacterial inactivation using a novel low-temperature atmospheric-pressure plasma device. J. Dent. Sci. 2014. [Google Scholar] [CrossRef]
- Sun, P.; Pan, J.; Tian, Y.; Bai, N.; Wu, H.; Wang, L.; Yu, C.; Zhang, J.; Zhu, W.; Becker, K.H.; et al. Tooth whitening with hydrogen peroxide assisted by a direct-current cold atmospheric-pressure air plasma microjet. Plasma Sci. IEEE Trans. 2010, 38, 1892–1896. [Google Scholar] [CrossRef]
- Vandamme, M.; Robert, E.; Dozias, S.; Sobilo, J.; Lerondel, S.; Le Pape, A.; Pouvesle, J. Response of Human Glioma U87 Xenografted on Mice to Non Thermal Plasma Treatment. Plasma Med. 2011, 1, 27–43. [Google Scholar] [CrossRef]
- Kim, S.J.; Chung, T.H.; Bae, S.H.; Leem, S.H. Induction of apoptosis in human breast cancer cells by a pulsed atmospheric pressure plasma jet. Appl. Phys. Lett. 2010, 97. [Google Scholar] [CrossRef]
- Partecke, L.I.; Evert, K.; Haugk, J.; Doering, F.; Normann, L.; Diedrich, S.; Weiss, F.; Evert, M.; Huebner, N.; Guenther, C.; et al. Tissue tolerable plasma (TTP) induces apoptosis in pancreatic cancer cells in vitro and in vivo. BMC Cancer 2012, 12. [Google Scholar] [CrossRef]
- Kaushik, N.K.; Kim, Y.H.; Han, Y.G.; Choi, E.H. Effect of jet plasma on T98G human brain cancer cells. Curr. Appl. Phys. 2013, 13, 176–180. [Google Scholar] [CrossRef]
- Barekzi, N.; Laroussi, M. Dose-dependent killing of leukemia cells by low-temperature plasma. J. Phys. D 2012, 45. [Google Scholar] [CrossRef]
- Kim, C.-H.; Kwon, S.; Bahn, J.H.; Lee, K.; Jun, S.I.; Rack, P.D.; Baek, S.J. Effects of atmospheric nonthermal plasma on invasion of colorectal cancer cells. Appl. Phys. Lett. 2010, 96. [Google Scholar] [CrossRef]
- Dröge, W. Free radicals in the physiological control of cell. Function 2002, 82, 47–95. [Google Scholar]
- Smith, J.; Ladi, E.; Mayer-Pröschel, M.; Noble, M. Redox state is a central modulator of the balance between self-renewal and differentiation in a dividing glial precursor cell. Proc. Natl. Acad. Sci. USA 2000, 97, 10032–10037. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Barzilai, A.; Yamamoto, K.-I. DNA damage responses to oxidative stress. DNA Repair 2004, 3, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, R.E.; Kaufmann, W.K.; Paules, R.S. Oxidative stress and cell cycle checkpoint function. Free Radic. Biol. Med. 2000, 28, 1387–1404. [Google Scholar] [CrossRef] [PubMed]
- Alkawareek, M.Y.; Gorman, S.P.; Graham, W.G.; Gilmore, B.F. Potential cellular targets and antibacterial efficacy of atmospheric pressure non-thermal plasma. Int. J. Antimicrob. Agents 2014, 43, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Leduc, M.; Guay, D.; Coulombe, S.; Leask, R.L. Effects of non-thermal plasmas on DNA and mammalian cells. Plasma Process. Polym. 2010, 7, 899–909. [Google Scholar] [CrossRef]
- Bahnev, B.; Bowden, M.D.; Stypczyńska, A.; Ptasińska, S.; Mason, N.J.; Braithwaite, N.J. A novel method for the detection of plasma jet boundaries by exploring DNA damage. Eur. Phys. J. D 2014, 68, 1–5. [Google Scholar] [CrossRef]
- Hosseinzadeh Colagar, A.; Memariani, H.; Sohbatzadeh, F.; Valinataj Omran, A. Nonthermal atmospheric argon plasma jet effects on Escherichia coli biomacromolecules. Appl. Biochem. Biotechnol. 2013, 171, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-M.; Kim, J.-I. Decomposition of biological macromolecules by plasma generated with helium and oxygen. J. Microbiol. 2006, 44, 466–471. [Google Scholar] [PubMed]
- Li, G.; Li, H.; Wang, L.; Wang, S.; Zhao, H.; Sun, W.; Xing, X.; Bao, C. Genetic effects of radio-frequency, atmospheric-pressure glow discharges with helium. Appl. Phys. Lett. 2008, 92. [Google Scholar] [CrossRef]
- Niemi, K.; O’Neill, C.; Cox, L.J.; Waskoenig, J.; Hyland, W.B.; McMahon, S.J.; Reuter, S.; Currell, F.J.; Graham, W.G.; O’Connell, D.; et al. Cold atmospheric pressure plasma jets: Interaction with plasmid DNA and tailored electron heating using dual-frequency excitation. AIP Confer. Pro. 2012, 1438, 23–28. [Google Scholar]
- O’Connell, D.; Cox, L.J.; Hyland, W.B.; McMahon, S.J.; Reuter, S.; Graham, W.G.; Gans, T.; Currell, F.J. Cold atmospheric pressure plasma jet interactions with plasmid DNA. Appl. Phys. Lett. 2011, 98. [Google Scholar] [CrossRef] [Green Version]
- Ptasinska, S.; Bahnev, B.; Stypczynska, A.; Bowden, M.; Mason, N.J.; Braithwaite, N.J. DNA strand scission induced by a non-thermal atmospheric pressure plasma jet. Phys. Chem. Chem. Phys. 2010, 12, 7779–7781. [Google Scholar] [CrossRef] [PubMed]
- Stypczyńska, A.; Ptasińska, S.; Bahnev, B.; Bowden, M.; Braithwaite, N.J.; Mason, N.J. The influence of amino acids on DNA damage induced by cold plasma radiation. Chem. Phys. Lett. 2010, 500, 313–317. [Google Scholar] [CrossRef]
- Yan, X.; Zou, F.; Lu, X.P.; He, G.; Shi, M.J.; Xiong, Q.; Gao, X.; Xiong, Z.; Li, Y.; Ma, F.Y.; et al. Effect of the atmospheric pressure nonequilibrium plasmas on the conformational changes of plasmid DNA. Appl. Phys. Lett. 2009, 95. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, D.; Ballato, J.; Cao, W.; Kim, S. Reactive oxygen species controllable non-thermal helium plasmas for evaluation of plasmid DNA strand breaks. Appl. Phys. Lett. 2012, 101. [Google Scholar] [CrossRef]
- Lackmann, J.-W.; Schneider, S.; Edengeiser, E.; Jarzina, F.; Brinckmann, S.; Steinborn, E.; Havenith, M.; Benedikt, J.; Bandow, J.E. Photons and particles emitted from cold atmospheric-pressure plasma inactivate bacteria and biomolecules independently and synergistically. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Lackmann, J.-W.; Schneider, S.; Narberhaus, F.; Benedikt, J.; Bandow, J.E. Characterization of Damage to Bacteria and Bio-macromolecules Caused by (V)UV Radiation and Particles Generated by a Microscale Atmospheric Pressure Plasma Jet. In Plasma for Bio-Decontamination, Medicine and Food Security; Machala, Z., Hensel, K.Y., Eds.; Springer Netherlands: Amsterdam, The Netherlands, 2012; pp. 17–29. [Google Scholar]
- Antoniu, A.; Nakajima, T.; Kurita, H.; Mizuno, A. Safety evaluation of nonthermal atmospheric pressure plasma liquid treatment: Single DNA molecule-based method. J. Electrost. 2014, 72, 210–217. [Google Scholar] [CrossRef]
- Kurita, H.; Nakajima, T.; Yasuda, H.; Takashima, K.; Mizuno, A.; Wilson, J.I.B.; Cunningham, S. Single-molecule measurement of strand breaks on large DNA induced by atmospheric pressure plasma jet. Appl. Phys. Lett. 2011, 99. [Google Scholar] [CrossRef]
- Kurita, H.; Shimizu, M.; Sano, K.; Nakajima, T.; Yasuda, H.; Takashima, K.; Mizuno, A. Radical reaction in aqueous media injected by atmospheric pressure plasma jet and protective effect of antioxidant reagents evaluated by single-molecule DNA measurement. Jpn. J. Appl. Phys. 2014, 53. [Google Scholar] [CrossRef]
- Sousa, J.S.; Bauville, G.; Lacour, B.; Puech, V.; Touzeau, M.; Ravanat, J. DNA oxidation by singlet delta oxygen produced by atmospheric pressure microdischarges. Appl. Phys. Lett. 2010, 97. [Google Scholar] [CrossRef]
- Sousa, J.; Girard, P.; Sage, E.; Ravanat, J.; Puech, V. DNA Oxidation by Reactive Oxygen Species Produced by Atmospheric Pressure Microplasmas. In Plasma for Bio-Decontamination, Medicine and Food Security; Machala, Z., Hensel, K.Y., Eds.; Springer Netherlands: Amsterdam, The Netherlands, 2012; pp. 107–119. [Google Scholar]
- Brun, P.; Vono, M.; Venier, P.; Tarricone, E.; Deligianni, V.; Martines, E.; Zuin, M.; Spagnolo, S.; Cavazzana, R.; Cardin, R.; et al. Disinfection of ocular cells and tissues by atmospheric-pressure cold plasma. PLoS One 2012, 7, e33245. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.W.; Kang, S.U.; Shin, Y.S.; Kim, K.I.; Seo, S.J.; Yang, S.S.; Lee, J.; Moon, E.; Baek, S.J.; Lee, K.; et al. Non-thermal atmospheric pressure plasma induces apoptosis in oral cavity squamous cell carcinoma: Involvement of DNA-damage-triggering sub-G1 arrest via the ATM/p53 pathway. Arch. Biochem. Biophys. 2014, 545, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, Y.; Stack, M.S.; Ptasinska, S. 3D Mapping of plasma effective areas via detection of cancer cell damage induced by atmospheric pressure plasma jets. J. Phys. Conf. Ser. 2014, 565, 12011–12016. [Google Scholar] [CrossRef]
- Kalghatgi, S.; Azizkhan-Clifford, J. DNA damage in mammalian cells by atmospheric pressure microsecond-pulsed dielectric barrier discharge plasma is not. Mediated via lipid peroxidation. Plasma Med. J. 2011, 1, 167–177. [Google Scholar] [CrossRef]
- Kalghatgi, S.; Fridman, A.; Azizkhan-Clifford, J.; Friedman, G. DNA damage in mammalian cells by non-thermal atmospheric pressure microsecond pulsed dielectric barrier discharge plasma is not mediated by ozone. Plasma Process. Polym. 2012, 9, 726–732. [Google Scholar] [CrossRef]
- Kim, G.J.; Kim, W.; Kim, K.T.; Lee, J.K. DNA damage and mitochondria dysfunction in cell apoptosis induced by nonthermal air plasma. Appl. Phys. Lett. 2010, 96. [Google Scholar] [CrossRef]
- Kim, K.; Choi, J.D.; Hong, Y.C.; Kim, G.; Noh, E.J.; Lee, J.; Yang, S.S. Atmospheric-pressure plasma-jet from micronozzle array and its biological effects on living cells for cancer therapy. Appl. Phys. Lett. 2011, 98. [Google Scholar] [CrossRef]
- Ma, Y.; Ha, C.S.; Hwang, S.W.; Lee, H.J.; Kim, G.C.; Lee, K.; Song, K. Non-thermal atmospheric pressure plasma preferentially induces apoptosis in p53-mutated cancer cells by activating ROS stress-response pathways. PLoS One 2014, 9, e91947. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramírez, P.; Cruz-Vallejo, V.; Peña-Eguiluz, R.; López-Callejas, R.; Rodríguez-Méndez, B.G.; Valencia-Alvarado, R.; Mercado-Cabrera, A.; Muñoz-Castro, A.E. Assessing cellular DNA damage from a helium plasma needle. Radiat. Res. 2013, 179, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Plewa, J.-M.; Yousfi, M.; Frongia, C.; Eichwald, O.; Ducommun, B.; Merbahi, N.; Lobjois, V. Low-temperature plasma-induced antiproliferative effects on multi-cellular tumor spheroids. New J. Phys. 2014, 16. [Google Scholar] [CrossRef]
- Volotskova, O.; Hawley, T.S.; Stepp, M.A.; Keidar, M. Targeting the cancer cell cycle by cold atmospheric plasma. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zou, F.; Zhao, S.; Lu, X.; He, G.; Xiong, Z.; Xiong, Q.; Zhao, Q.; Deng, P.; Huang, J.; et al. On the mechanism of plasma inducing cell apoptosis. Plasma Sci. IEEE Trans. 2010, 38, 2451–2457. [Google Scholar] [CrossRef]
- Choi, J.-H.; Lee, H.; Lee, J.; Hong, J.; Kim, G. Low-temperature atmospheric plasma increases the expression of anti-aging genes of skin cells without causing cellular damages. Arch. Dermatol. Res. 2013, 305, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lazović, S.; Maletić, D.; Leskovac, A.; Filipović, J.; Puač, N.; Malović, G.; Joksić, G.; Petrović, Z.L. Plasma induced DNA damage: Comparison with the effects of ionizing radiation. Appl. Phys. Lett. 2014, 105. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, K.; Kang, K.; Lee, J.; Yang, S.; Chung, W. Atmospheric-pressure plasma jet induces DNA double-strand breaks that require a Rad51-mediated homologous recombination for repair in Saccharomyces cerevisiae. Arch. Biochem. Biophys. 2014, 560, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.-H.; Kim, Y.; Lee, J.; Shim, G.; Uhm, H.; Park, G.; Choi, E.H. Effects of background fluid on the efficiency of inactivating yeast with non-thermal atmospheric pressure plasma. PLoS One 2013, 8, e66231. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.G.; Cooper, M.; Yost, A.; Paff, M.; Ercan, U.K.; Fridman, G.; Friedman, G.; Fridman, A.; Brooks, A.D. Nonthermal dielectric-barrier discharge plasma-induced inactivation involves oxidative DNA damage and membrane lipid peroxidation in Escherichia coli. Antimicrob. Agents Chemother. 2011, 55, 1053–1062. [Google Scholar] [CrossRef]
- Kvam, E.; Davis, B.; Mondello, F.; Garner, A.L. Nonthermal atmospheric plasma rapidly disinfects multidrug-resistant microbes by inducing cell surface damage. Antimicrob. Agents Chemother. 2012, 56, 2028–2036. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.; Abramzon, N.; Jackson, J.O.; Lin, W. Gas. discharge plasmas are effective in inactivating Bacillus and Clostridium spores. Appl. Microbiol. Biotechnol. 2012, 93, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Mols, M.; Mastwijk, H.; Nierop Groot, M.; Abee, T. Physiological and transcriptional response of Bacillus cereus treated with low-temperature nitrogen gas plasma. J. Appl. Microbiol. 2013, 115, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Collins, G.; Pruden, A. Differential gene expression in Escherichia coli following exposure to nonthermal atmospheric pressure plasma. J. Appl. Microbiol. 2009, 107, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Winter, T.; Winter, J.; Polak, M.; Kusch, K.; Mäder, U.; Sietmann, R.; Ehlbeck, J.; van Hijum, S.; Weltmann, K.; Hecker, M.; Kusch, H. Characterization of the global impact of low temperature gas plasma on vegetative microorganisms. Proteomics 2011, 11, 3518–3530. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Patil, S.; Keener, K.M.; Cullen, P.J.; Bourke, P. Bacterial inactivation by high-voltage atmospheric cold plasma: Influence of process parameters and effects on cell leakage and DNA. J. Appl. Microbiol. 2014, 116, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Venezia, R.; Orrico, M.; Houston, E.; Yin, S.; Naumova, Y.Y. Lethal activity of nonthermal plasma sterilization against microorganisms. Infect. Control Hosp. Epidemiol. 2008, 29, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H.; Miura, T.; Kurita, H.; Takashima, K.; Mizuno, A. Biological evaluation of DNA damage in bacteriophages inactivated by atmospheric pressure cold plasma. Plasma Process. Polym. 2010, 7, 301–308. [Google Scholar] [CrossRef]
- Wang, L.Y.; Huang, Z.L.; Li, G.; Zhao, H.X.; Xing, X.H.; Sun, W.T.; Li, H.P.; Gou, Z.X.; Bao, C.Y. Novel mutation breeding method for Streptomyces avermitilis using an atmospheric pressure glow discharge plasma. J. Appl. Microbiol. 2010, 108, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Jin, L.; Zhang, C.; Tan, Y.; Jiang, P.; Ge, N.; Li, H.; Xing, X. Rapid mutation of spirulina platensis by a new mutagenesis system of atmospheric and room temperature plasmas (ARTP) and generation of a mutant library with diverse phenotypes. PLoS One 2013, 8, e77046. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 634–1658. [Google Scholar] [CrossRef]
- Kuznetsova, A.A.; Dmitrii, G.K.; Olga, S.F. Oxidation of DNA and its components with reactive oxygen species. Russ. Chem. Rev. 2009, 78. [Google Scholar] [CrossRef]
- Shafirovich, V.; Crean, C.; Geacintov, N.E. Reactions of reactive nitrogen species and carbonate radical anions with DNA. In Radical and Radical Ion Reactivity in Nucleic Acid Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 325–355. [Google Scholar]
- Dedon, P.C.; Tannenbaum, S.R. Reactive nitrogen species in the chemical biology of inflammation. Arch. Biochem. Biophys. 2004, 423, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Buxton, G.V.; Greenstock, C.L.; Helman, W.P.; Ross, A.B. Critical review of rate constants for reactions of hydrated electrons, hydrogen atoms and hydroxyl radicals (●OH/●O− in aqueous solution. J. Phys. Chem. Ref. Data 1988, 17, 513–886. [Google Scholar] [CrossRef]
- Neta, P.; Huie, R.E.; Ross, A.B. Rate constants for reactions of inorganic radicals in aqueous solution. J. Phys. Chem. Ref. Data 1988, 17, 1027–1284. [Google Scholar] [CrossRef]
- Lewis, R.S.; Deen, W.M. Kinetics of the reaction of nitric oxide with oxygen in aqueous solutions. Chem. Res. Toxicol. 1994, 7, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Manoj, P.; Mohan, H.; Mittal, J.P.; Manoj, V.M.; Aravindakumar, C.T. Charge transfer from 2-aminopurine radical cation and radical anion to nucleobases: A pulse radiolysis study. Chem. Phys. 2007, 331, 351–358. [Google Scholar] [CrossRef]
- Von Sonntag, C. Free-Radical-Induced DNA Damage and Its Repair: A Chemical Perspective 2010; Springer: New York, NY, USA, 2010. [Google Scholar]
- Sharma, V.K. Oxidation of Amino Acids, Peptides, and Proteins; Wiley: New Jersey, NJ, USA, 2013. [Google Scholar]
- Dizdaroglu, M. Chemical determination of free radical-induced damage to DNA. Free Radic. Biol. Med. 1991, 10, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Aruoma, O.I. DNA damage by oxygen-derived species Its mechanism and measurement in mammalian systems. FEBS Lett. 1991, 281, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.; Wemmer, D.E.; Linn, S. Preferential binding and structural distortion by Fe2+ at RGGG-containing DNA sequences correlates with enhanced oxidative cleavage at such sequences. Nucleic Acids Res. 2005, 33, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Flint, D.H.; Tuminello, J.F.; Emptage, M.H. The inactivation of Fe-S cluster containing hydro-lyases by superoxide. J. Biol. Chem. 1993, 268, 22369–22376. [Google Scholar] [PubMed]
- Kuo, C.F.; Mashino, T.; Fridovich, I. α,β-Dihydroxyisovalerate dehydratase: A superoxide-sensitive enzyme. J. Biol. Chem. 1987, 262, 4724–4727. [Google Scholar] [PubMed]
- Keyer, K.; Gort, A.S.; Imlay, J.A. Superoxide and the production of oxidative DNA damage. J. Bacteriol. 1995, 177, 6782–6790. [Google Scholar] [PubMed]
- Sies, H.; Menck, C.F.M. Singlet oxygen induced DNA damage. Mutat. Res. DNAging 1992, 275, 367–375. [Google Scholar] [CrossRef]
- Devasagayam, T.P.A.; di Mascio, P.; Kaiser, S.; Sies, H. Singlet oxygen induced single-strand breaks in plasmid pBR322 DNA: The enhancing effect of thiols. Biochim. Biophys. Acta Gene Struct. Expr. 1991, 1088, 409–412. [Google Scholar] [CrossRef]
- Ravanat, J.-L.; Cadet, J. Reaction of singlet oxygen with 2'-deoxyguanosine and DNA. Isolation and characterization of the main oxidation products. Chem. Res. Toxicol. 1995, 8, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.E.; Price, S.; Maidt, L.; Gutteridge, J.M.; Floyd, R.A. Methylene blue plus light mediates 8-hydroxy 2'-deoxyguanosine formation in DNA preferentially over strand breakage. Nucleic Acids Res. 1990, 18, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.-J.; Kao, H.; Chan, C.; Chang, W.P. Effects of ozone on DNA single-strand breaks and 8-oxoguanine formation in A549 cells. Environ. Res. 2003, 93, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Inoue, S.; Hiraku, Y.; Kawanishi, S. Mechanism of site-specific DNA damage induced by ozone. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 585, 60–70. [Google Scholar] [CrossRef]
- Kuipers, G.K.; Lafleur, M.V.M. Characterization of DNA damage induced by γ-radiation-derived water radicals, using DNA repair enzymes. Int. J. Radiat. Biol. 1998, 74, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sevilla, M.D. Role of excited states in low-energy electron. (LEE) Induced strand breaks in DNA model systems: Influence of aqueous environment. ChemPhysChem 2009, 10, 1426–1430. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, L.J.; Moore, K.; Caulfield, J.L.; Tannenbaum, S.R.; Dedon, P.C. Quantitation of 8-oxoguanine and strand breaks produced by four oxidizing agents. Chem. Res. Toxicol. 1997, 10, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Wagner, J.R. Oxidatively generated base damage to cellular DNA by hydroxyl radical and one-electron oxidants: Similarities and differences. Arch. Biochem. Biophys. 2014, 557, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Steenken, S. Purine bases, nucleosides, and nucleotides: Aqueous solution redox chemistry and transformation reactions of their radical cations and e- and OH adducts. Chem. Rev. 1989, 89, 503–520. [Google Scholar] [CrossRef]
- Steenken, S.; Jovanovic, S.V. How easily oxidizable Is DNA? One-electron. Reduction potentials of adenosine and guanosine radicals in aqueous solution. J. Am. Chem. Soc. 1997, 119, 617–618. [Google Scholar] [CrossRef]
- Steenken, S.; Jovanovic, S.V.; Bietti, M.; Bernhard, K. The trap depth (in DNA) of 8-oxo-7,8-dihydro-2′deoxyguanosine as derived from electron-transfer equilibria in aqueous solution. J. Am. Chem. Soc. 2000, 122, 2373–2374. [Google Scholar] [CrossRef]
- Yuan, B.-F.; Feng, Y.-Q. Recent advances in the analysis of 5-methylcytosine and its oxidation products. TrAC Trends Anal. Chem. 2014, 54, 24–35. [Google Scholar] [CrossRef]
- Cadet, J.; Teoule, R. Comparative study of oxidation of nucleic acid components by hydroxyl radicals, singlet oxygen and superoxide anion radicals. Photochem. Photobiol. 1978, 28, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Misiaszek, R.; Crean, C.; Joffe, A.; Geacintov, N.E.; Shafirovich, V. Oxidative DNA damage associated with combination of guanine and superoxide radicals and repair mechanisms via radical trapping. J. Biol. Chem. 2004, 279, 32106–32115. [Google Scholar] [CrossRef] [PubMed]
- Lafleur, M.V.M.; Nieuwint, A.W.M.; Aubry, J.M.; Kortbeek, H.; Arwert, F.; Joenje, H. DNA damage by chemically generated singlet oxygen. Free Radic. Res. 1987, 2, 343–350. [Google Scholar] [CrossRef]
- Luo, W.; Muller, J.G.; Rachlin, E.M.; Burrows, C.J. Characterization of hydantoin products from one-electron oxidation of 8-oxo-7,8-dihydroguanosine in a nucleoside model. Chem. Res. Toxicol. 2001, 14, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Hayon, E.; Simic, M. Addition of hydroxyl radicals to pyrimidine bases and electron transfer reactions of intermediates to quinones. J. Am. Chem. Soc. 1973, 95, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.H.; Geacintov, N.E.; Shafirovich, V. Generation of guanine–thymidine cross-links in DNA by peroxynitrite/carbon dioxide. Chem. Res. Toxicol. 2011, 24, 1144–1152. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.S.; Cui, L.; Taghizadeh, K.; Wishnok, J.S.; Chan, W.; DeMott, M.S.; Babu, I. R.; Tannenbaum, S.R.; Dedon, P.C. In Situ Analysis of 8-oxo-7,8-dihydro-2′-deoxyguanosine oxidation reveals sequence- and agent-specific damage spectra. J. Am. Chem. Soc. 2012, 134, 18053–18064. [Google Scholar] [CrossRef] [PubMed]
- Crean, C.; Uvaydov, Y.; Geacintov, N.E.; Shafirovich, V. Oxidation of single-stranded oligonucleotides by carbonate radical anions: Generating intrastrand cross-links between guanine and thymine bases separated by cytosines. Nucleic Acids Res. 2008, 36, 742–755. [Google Scholar] [CrossRef] [PubMed]
- Madugundu, G.S.; Wagner, J.R.; Cadet, J.; Kropachev, K.; Yun, B.H.; Geacintov, N.E.; Shafirovich, V. Generation of guanine-thymine cross-links in human cells by one-electron. Oxidation mechanisms. Chem. Res. Toxicol. 2013, 26, 1031–1033. [Google Scholar] [CrossRef]
- Mitra, J.; Guerrero, E.; Hegde, P.; Wang, H.; Boldogh, I.; Rao, K.; Mitra, S.; Hegde, M. New perspectives on oxidized genome damage and repair inhibition by pro-oxidant metals in neurological diseases. Biomolecules 2014, 4, 678–703. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, P.J. DNA repair deficiency and neurological disease. Nat. Rev. Neurosci. 2009, 10, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Boldogh, I.; Izumi, T.; Hazra, T.K. Complexities of the DNA base excision repair pathway for repair of oxidative DNA damage. Environ. Mol. Mutagen. 2001, 38, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Bozzella, M.; Seluanov, A.; Gorbunova, V. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle 2008, 7, 2902–2906. [Google Scholar] [CrossRef] [PubMed]
- Shrivastav, M.; de Haro, L.P.; Nickoloff, J.A. Regulation of DNA double-strand break repair pathway choice. Cell Res. 2008, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Takata, M.; Sasaki, M.S.; Sonoda, E.; Morrison, C.; Hashimoto, M.; Utsumi, H.; Yamaguchi-Iwai, Y.; Shinohara, A.; Takeda, S. Homologous recombination and non-homologous end-joining pathways of DNA double‐strand break repair have overlapping roles in the maintenance of chromosomal integrity in vertebrate cells. EMBO J. 1998, 17, 5497–5508. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals. Reactive oxygen species and human disease: A critical evaluation with special reference to atherosclerosis. Br. J. Exp. Pathol. 1989, 70, 737–757. [Google Scholar]
- Burtscher, H.; Berner, S.; Seibl, R.; Muhlegger, K. Nucleic acids. In Ullmann’s Biotechnology and Biochemical Engineering; Wiley-VCH: Weinheim, Germany, 2007; pp. 157–194. [Google Scholar]
- Deweese, J.E.; Osheroff, M.A.; Osheroff, N. DNA topology and topoisomerases: Teaching a “knotty” subject. Biochem. Mol. Biol. Educ. 2008, 37, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Gaens, W.V.; Bogaerts, A. Kinetic modelling for an atmospheric pressure argon plasma jet in humid air. J. Phys. D 2013, 46, 275201. [Google Scholar] [CrossRef]
- Krisch, R.E.; Flick, M.B.; Trumbore, C.N. Radiation chemical mechanisms of single- and double-strand break formation in irradiated SV40 DNA. Radiat. Res. 1991, 126, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Fridman, G.; Brooks, A.D.; Balasubramanian, M.; Fridman, A.; Gutsol, A.; Vasilets, V.N.; Ayan, H.; Friedman, G. Comparison of direct and indirect effects of non-thermal atmospheric-pressure plasma on bacteria. Plasma Process. Polym. 2007, 4, 370–375. [Google Scholar] [CrossRef]
- Neyts, E.C.; Yusupov, M.; Verlackt, C.C.; Bogaerts, A. Computer simulations of plasma-biomolecule and plasma-tissue interactions for a better insight in plasma medicine. J. Phys. D 2014, 47, 293001. [Google Scholar] [CrossRef]
- Vilenchik, M.M.; Knudson, A.G. Endogenous DNA double-strand breaks: Production, fidelity of repair, and induction of cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 12871–12876. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.W.; Hunt, T. The Cell Cycle: An Introduction; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Ward, I.M.; Chen, J. Histone H2AX is phosphorylated in an ATR-dependent manner in response to replicational stress. J. Biol. Chem. 2001, 276, 47759–47762. [Google Scholar] [CrossRef] [PubMed]
- Bohgaki, T.; Bohgaki, M.; Hakem, R. DNA double-strand break signaling and human disorders. Genome Integr. 2010, 1. [Google Scholar] [CrossRef]
- Jazayeri, A.; Falck, J.; Lukas, C.; Bartek, J.; Smith, G.C.M.; Lukas, J.; Jackson, S.P. ATM- and cell cycle-dependent regulation of ATR in response to DNA double-strand breaks. Nat. Cell Biol. 2006, 8, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.E.; Medhurst, A.L.; Dart, D.A.; Lakin, N.D. Recruitment of ATR to sites of ionising radiation-induced DNA damage requires ATM and components of the MRN protein complex. Oncogene 2006, 25, 3894–3904. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, M.; Martinez-Pastor, B.; Murga, M.; Toledo, L.I.; Gutierrez-Martinez, P.; Lopez, E.; Fernandez-Capetillo, O. ATM regulates ATR chromatin loading in response to DNA double-strand breaks. J. Exp. Med. 2006, 203, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.S.; Cortez, D. Rapid Activation of ATR by Ionizing Radiation Requires ATM and Mre11. J. Biol. Chem. 2006, 281, 9346–9350. [Google Scholar] [CrossRef]
- Stiff, T.; Walker, S.A.; Cerosaletti, K.; Goodarzi, A.A.; Petermann, E.; Concannon, P.; O’Driscoll, M.; Jeggo, P.A. ATR-dependent phosphorylation and activation of ATM in response to UV treatment or replication fork stalling. EMBO J. 2006, 25, 5775–5782. [Google Scholar] [CrossRef] [PubMed]
- Helton, E.S.; Chen, X. p53 modulation of the DNA damage response. J. Cell. Biochem. 2007, 100, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-S.; Qian, Y.; Chen, X. Examination of the expanding pathways for the regulation of p21 expression and activity. Cell Signal. 2010, 22, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Okafuji, M.; Traganos, F.; Luther, E.; Holden, E.; Darzynkiewicz, Z. Assessment of histone H2AX phosphorylation induced by DNA topoisomerase I and II inhibitors topotecan and mitoxantrone and by the DNA cross-linking agent cisplatin. Cytom. Part A 2004, 58, 99–110. [Google Scholar] [CrossRef]
- Slupphaug, G.; Kavli, B.; Krokan, H.E. The interacting pathways for prevention and repair of oxidative DNA damage. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2003, 531, 231–251. [Google Scholar] [CrossRef]
- Breen, A.P.; Murphy, J.A. Reactions of oxyl radicals with DNA. Free Radic. Biol. Med. 1995, 18, 1033–1077. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Bulky DNA lesions induced by reactive oxygen species. Chem. Res. Toxicol. 2008, 21, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.M., III; McNeill, D.R. Base excision repair and the central nervous system. Neuroscience 2007, 145, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Le May, N.; Egly, J.; Coin, F. True lies: The double life of the nucleotide excision repair factors in transcription and DNA repair. J. Nucleic Acids 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Hader, D.-P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Graham, W.G.; Schaper, L.; Muir, M.; Currell, F.J. The effect of electrical discharges in the cell media on their viability and DNA damage and comparison with the effect of X-rays. Plasma Med. J. 2012, 2, 169–178. [Google Scholar] [CrossRef]
- Vandamme, M.; Robert, E.; Lerondel, S.; Sarron, V.; Ries, D.; Dozias, S.; Sobilo, J.; Gosset, D.; Kieda, C.; Legrain, B.; et al. ROS implication in a new antitumor strategy based on non-thermal plasma. Int. J. Cancer 2012, 130, 2185–2194. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [PubMed]
- Arends, M.J.; Morris, R.G.; Wyllie, A.H. Apoptosis the role of the endonuclease. Am. J. Pathol. 1990, 136, 593–608. [Google Scholar] [PubMed]
- Oberhammer, F.; Wilson, J.W.; Dive, C.; Morris, I.D.; Hickman, J.A.; Wakeling, A.E.; Walker, P.R.; Sikorska, M. Apoptotic death in epithelial cells: Cleavage of DNA to 300 and/or 50 kb fragments prior to or in the absence of internucleosomal fragmentation. EMBO J. 1993, 12, 3679–3684. [Google Scholar] [PubMed]
- MacPhail, S.H.; Banáth, J.P.; Yu, Y.; Chu, E.; Olive, P.L. Cell cycle-dependent expression of phosphorylated histone H2AX: Reduced expression in unirradiated but not X-irradiated G1-phase cells. Radiat. Res. 2003, 159, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singh, K.; Almasan, A. Histone H2AX phosphorylation: A marker for DNA damage. In DNA Repair Protocols; Humana Press-Springer: New York, NY, USA, 2012. [Google Scholar]
- Dantzer, F.; de la Rubia, G.; Ménissier-de Murcia, J.; Hostomsky, Z.; de Murcia, G.; Schreiber, V. Base excision repair is impaired in mammalian cells lacking poly(ADP-ribose) polymerase-1. Biochemistry 2000, 39, 7559–7569. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.; Desnoyers, S.; Ottaviano, Y.; Davidson, N.E.; Poirier, G.G. Specific proteolytic cleavage of poly(ADP-ribose) polymerase: An early marker of chemotherapy-induced apoptosis. Cancer Res. 1993, 53, 3976–3985. [Google Scholar] [PubMed]
- Los, M.; Mozoluk, M.; Ferrari, D.; Stepczynska, A.; Stroh, C.; Renz, A.; Herceg, Z.; Wang, Z.; Schulze-Osthoff, K. Activation and caspase-mediated inhibition of PARP: A molecular switch between fibroblast necrosis and apoptosis in death receptor signaling. Mol. Biol. Cell 2002, 13, 978–988. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arjunan, K.P.; Sharma, V.K.; Ptasinska, S. Effects of Atmospheric Pressure Plasmas on Isolated and Cellular DNA—A Review. Int. J. Mol. Sci. 2015, 16, 2971-3016. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16022971
Arjunan KP, Sharma VK, Ptasinska S. Effects of Atmospheric Pressure Plasmas on Isolated and Cellular DNA—A Review. International Journal of Molecular Sciences. 2015; 16(2):2971-3016. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16022971
Chicago/Turabian StyleArjunan, Krishna Priya, Virender K. Sharma, and Sylwia Ptasinska. 2015. "Effects of Atmospheric Pressure Plasmas on Isolated and Cellular DNA—A Review" International Journal of Molecular Sciences 16, no. 2: 2971-3016. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16022971