
Molecular Dynamics Simulations of Acylpeptide Hydrolase Bound to Chlorpyrifosmethyl Oxon and Dichlorvos

Abstract
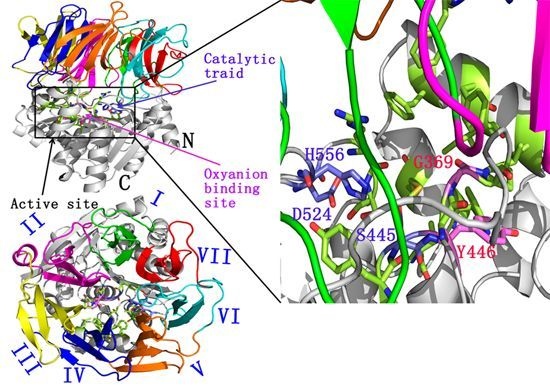
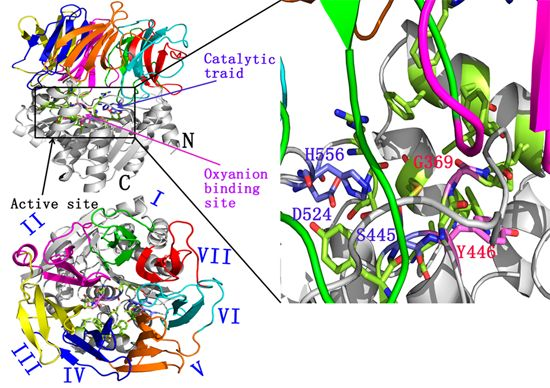
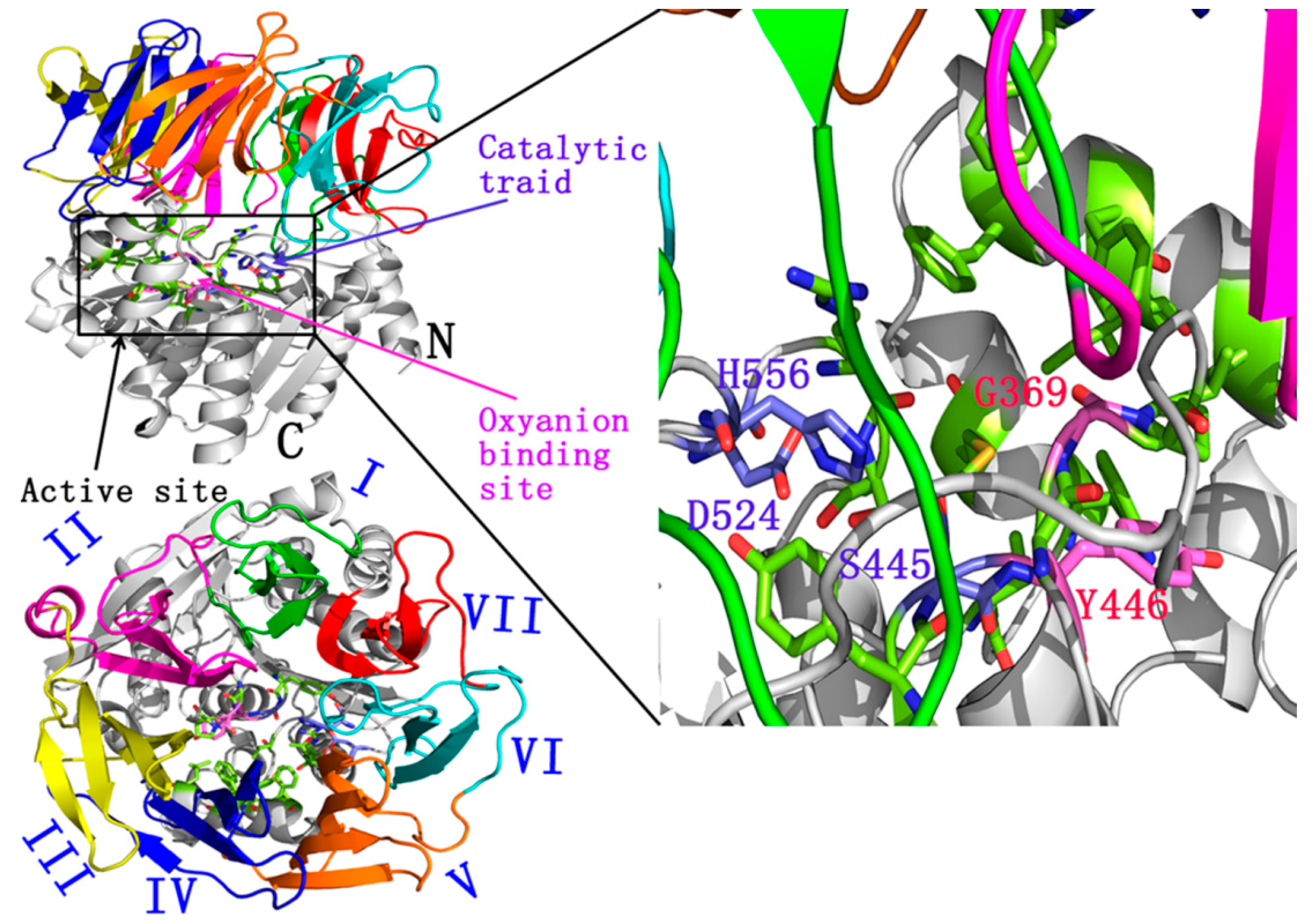
:
1. Introduction

2. Results and Discussion
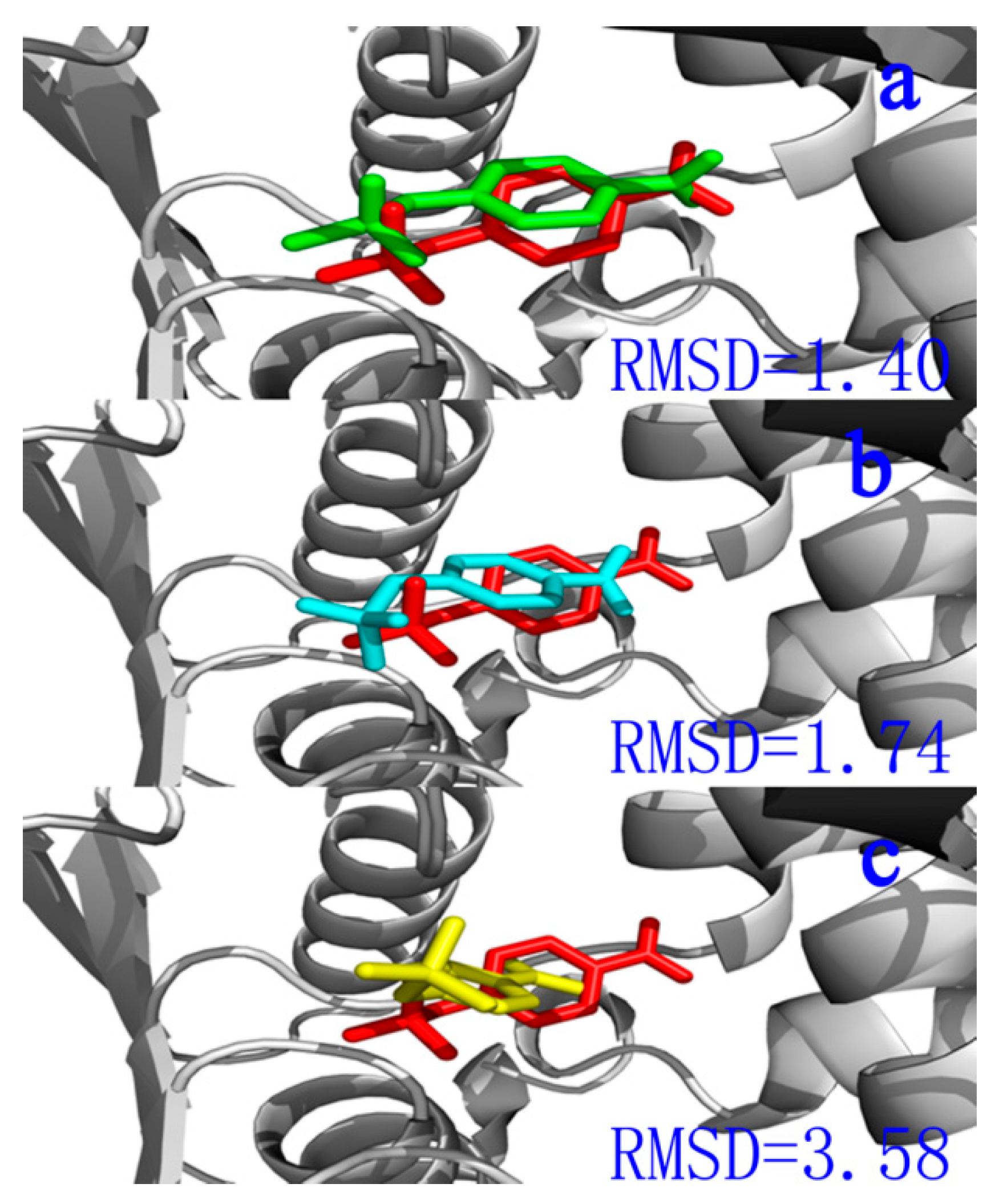
2.1. Docking Study





{kind=link}
{kind=link}
{kind=link}
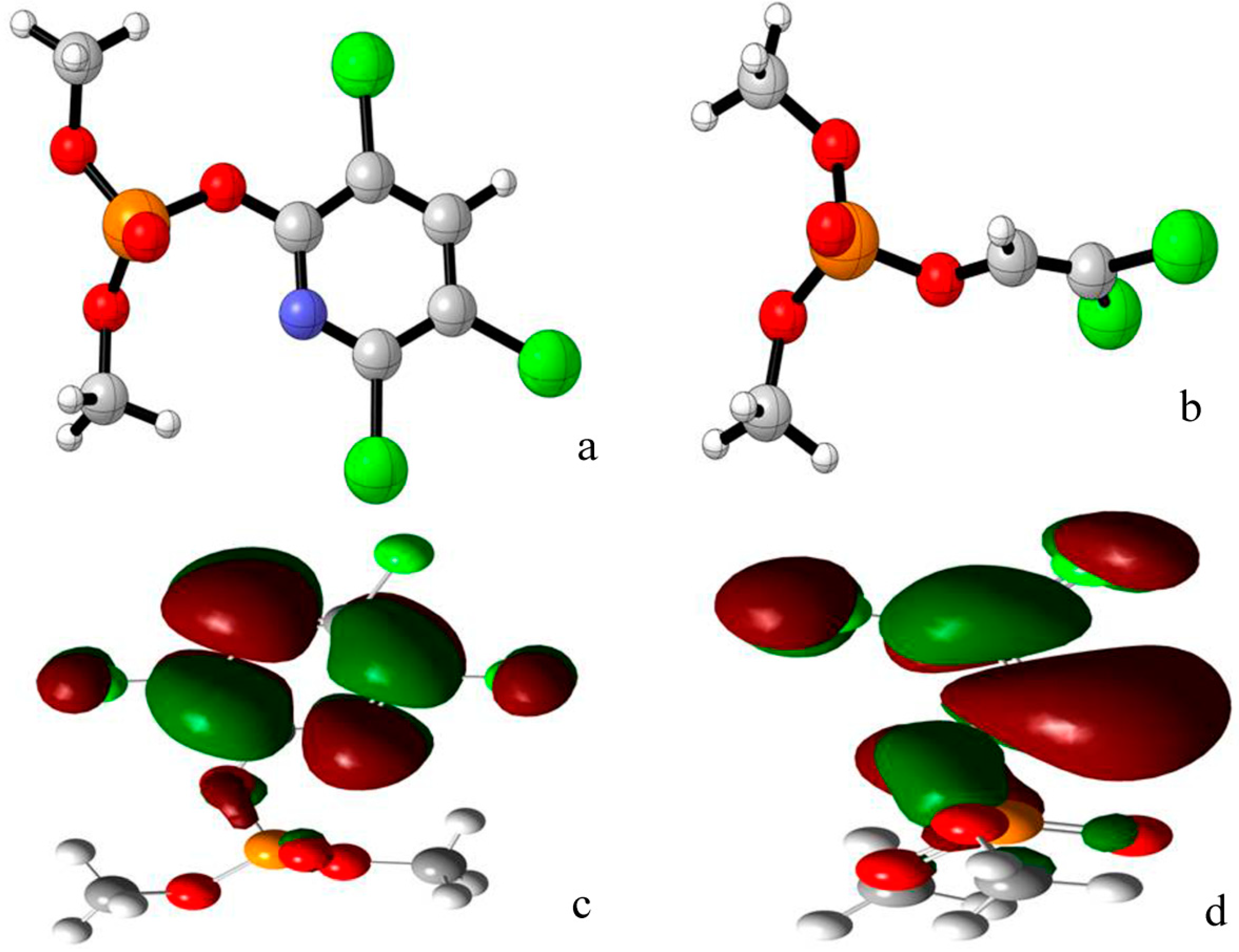{kind=link}
{kind=link}
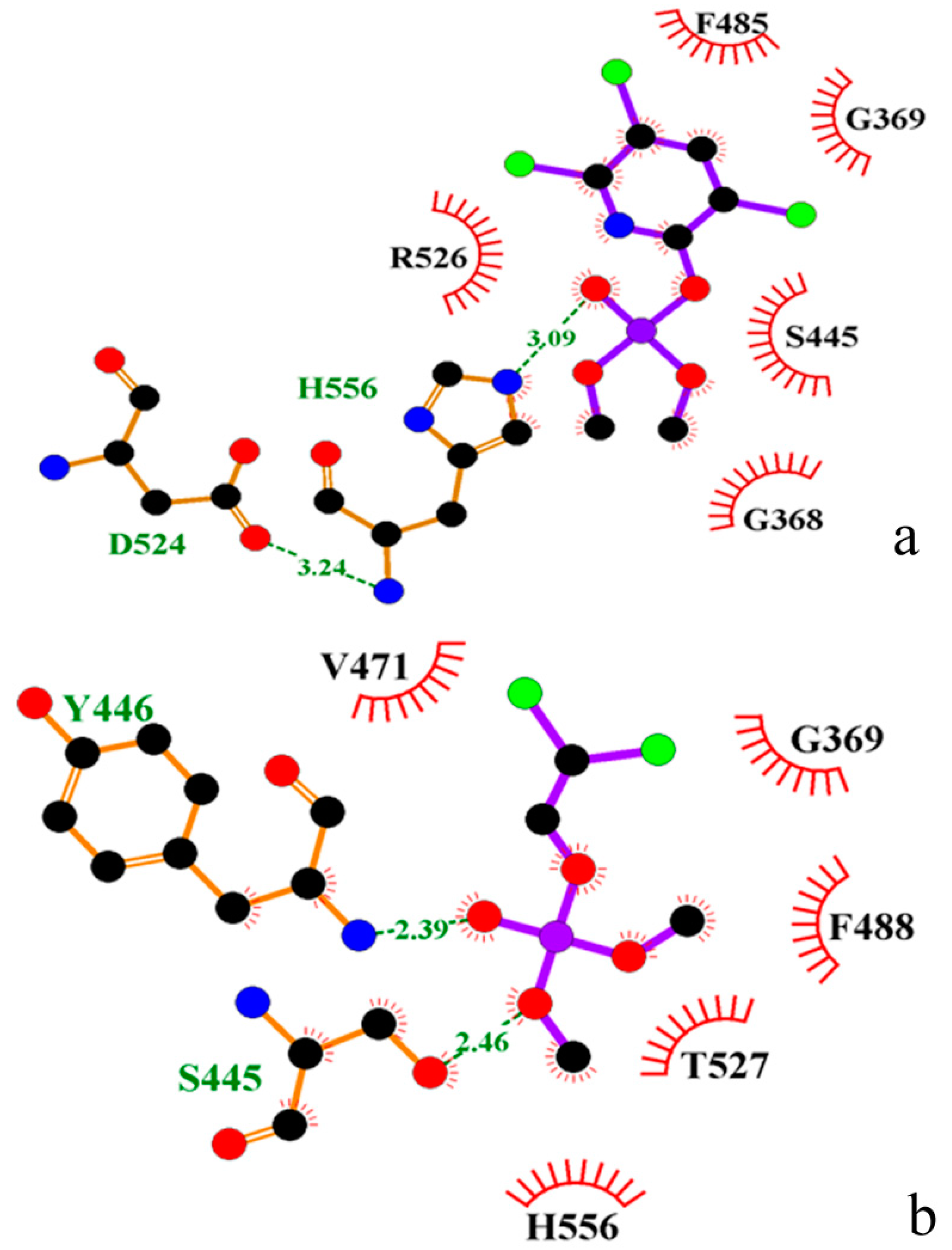{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Residue Number a | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 526 | 369 | 527 | 445 | 488 | 485 | 556 | 446 | 471 | 368 | 524 | |
| A. pernix | Arg | Gly | Thr | Ser | Phe | Phe | His | Tyr | Val | Gly | Asp |
| Human | Arg | Gly | Val | Ser | Cys | Phe | His | His | Val | Gly | Asp |
| Pig | Arg | Gly | Val | Ser | Ser | Phe | His | His | Val | Gly | Asp |
| Rat | Arg | Gly | Val | Ser | Leu | Leu | His | His | Val | Gly | Asp |
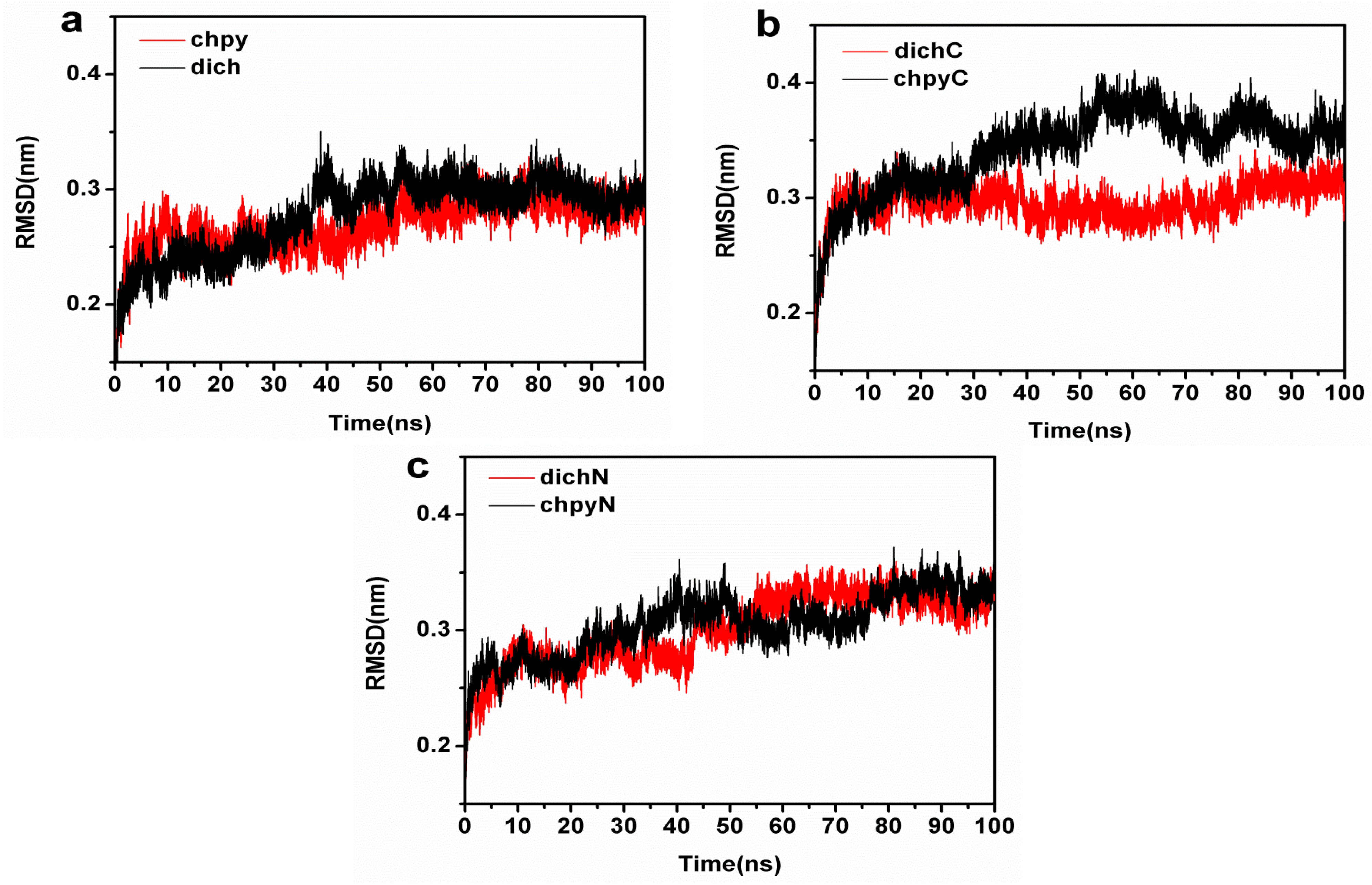
2.2. Structural Stability in Conventional MD Simulations


| Hydrogen Bonds | Distance (Å) | Chorpyrifosmethyl Oxon-APH | Dichlorvos-APH |
|---|---|---|---|
| VAL46:HN-ASN559:O | 2.41 | <10 | 0.19 |
| ARG113:HH22-SER525:O | 2.43 | <10 | 0.12 |
| GLY173:HN-GLN491:OE1 | 1.63 | <10 | 0.26 |
| ASN284:HD21-ASP376:O | 1.64 | 0.39 | 0.77 |
| LEU302:HN-ASP376:OD1 | 2.13 | 0.93 | 0.94 |
| ARG327:HN-PRO323:O | 1.91 | 0.91 | 0.92 |
| GLU405:HN-THR214:O | 1.98 | <10 | 0.45 |
| ARG408:HH22-GLY173:O | 1.88 | <10 | 0.15 |

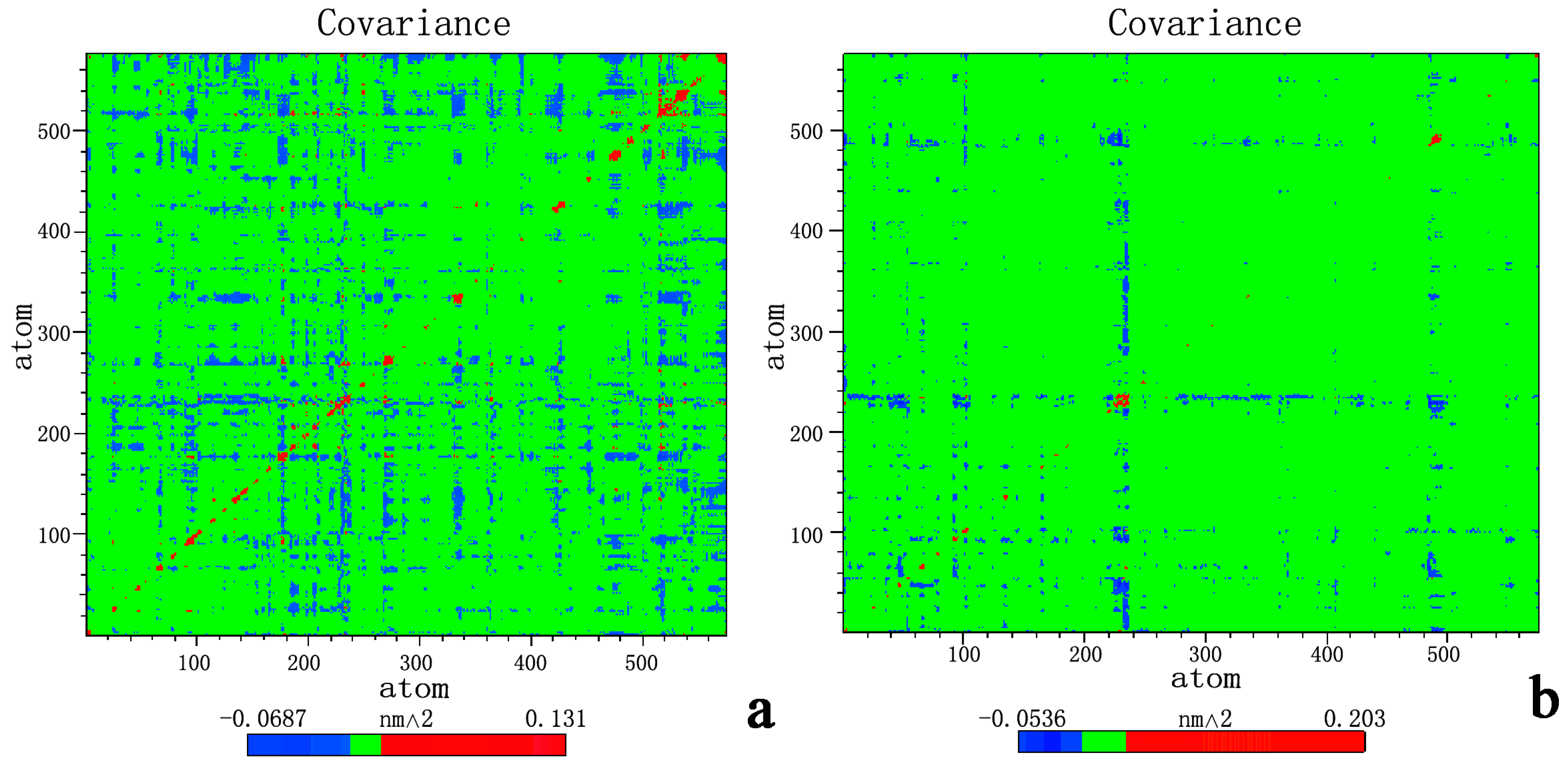
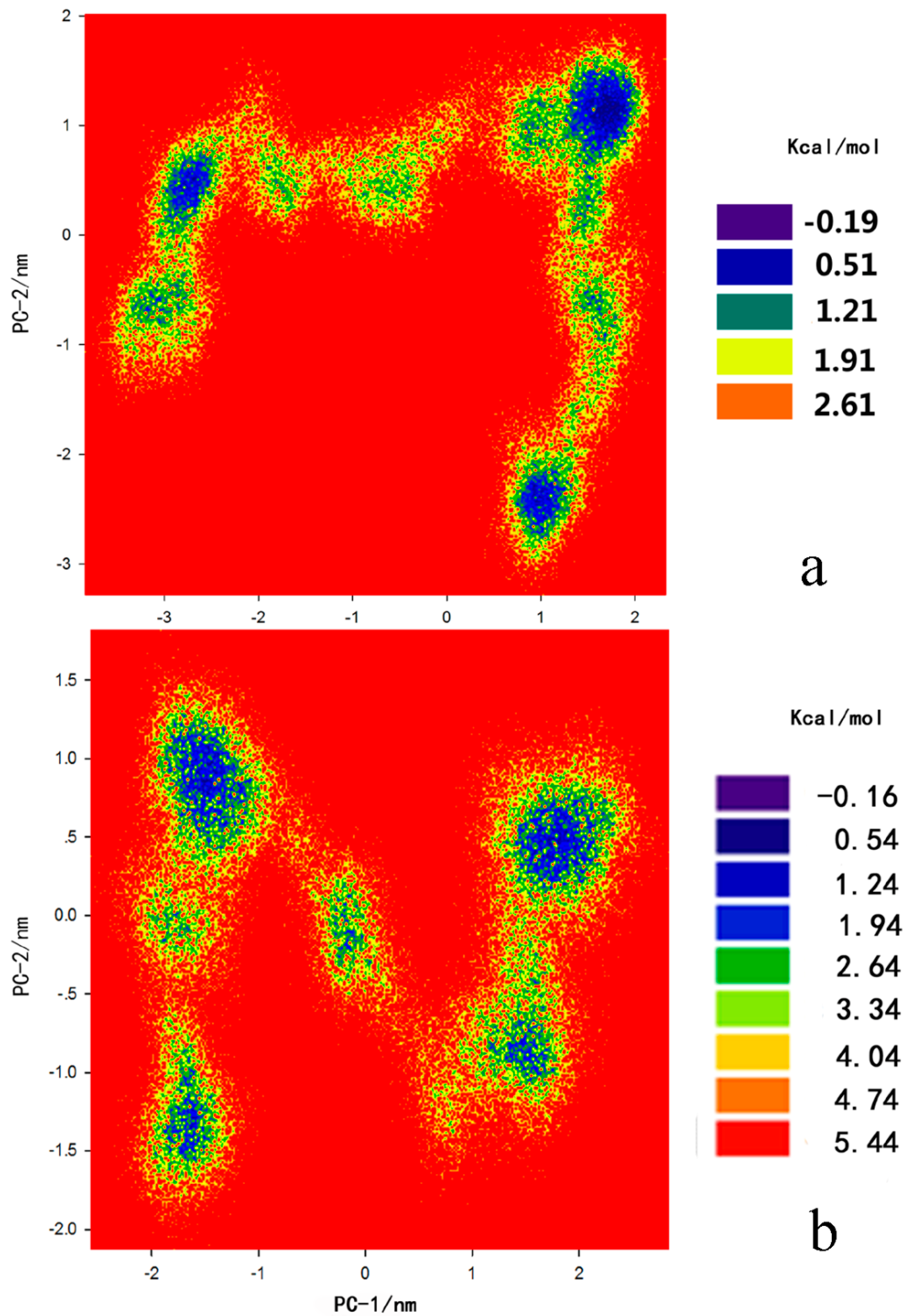
2.3. Principal Component Analysis and Free-Energy Landscape

2.4. MM/PBSA Calculation

| Energy Components (kcal·mol−1) | Dichlorvos | Chlorpyrifosmethyl Oxon |
|---|---|---|
| ∆Eele | −18.28 | −2.63 |
| ∆EvdW | −28.49 | −29.64 |
| ∆GPB a | 31.60 | 15.09 |
| ∆Gnp b | −4.08 | −3.72 |
| Nonpolar | −32.57 | −33.36 |
| Polar | 13.32 | 12.46 |
| ∆Gbind | −19.25 | −30.90 |
3. Experimental Section
3.1. Docking Study
3.2. Conventional Molecular Dynamics Simulations
3.3. Principal Component Analysis and Free-Energy Landscape
3.4. Calculations of MM/PBSA Binding Free Energy
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, H.F.; Zheng, B.S.; Peng, Y.; Lou, Z.Y.; Feng, Y.; Rao, Z.H. Expression, purification and crystal structure of a truncated acylpeptide hydrolase from Aeropyrum pernix K1. Acta Biochim. Biophys. Sin. 2005, 37, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.L.; Wang, H.L.; Zhang, Y.H.; Gao, L.; Feng, Y. Alteration of substrate specificities of thermophilic α/β hydrolases through domain swapping and domain interface optimization. Acta Biochim. Biophys. Sin. 2012, 44, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.M.; Zheng, B.S.; Wang, Y.P.; Chen, Y.Q.; Manco, G.; Feng, Y. The conserved N-terminal helix of acylpeptide hydrolase from archaeon Aeropyrum pernix K1 is important for its hyperthermophilic activity. Biochim. Biophys. Acta 2008, 1784, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Perrier, J.; Durand, A.; Giardina, T.; Puigserver, A. Catabolism of intracellular N-terminal acetylated proteins: Involvement of acylpeptide hydrolase and acylase. Biochimie 2005, 87, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Liu, Y.L.; Feng, Y. Discrimination of esterase and peptidase activities of acylaminoacyl peptidase from hyperthermophilic Aeropyrum pernix K1 by a single mutation. J. Biol. Chem. 2006, 281, 18618–18625. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Fülöp, V. Structure–function properties of prolyl oligopeptidase family enzymes. Cell Biochem. Biophys. 2006, 44, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.G.; Johnson, M.K.; Ray, D.E. Identification of acylpeptide hydrolase as a sensitive site for reaction with organophosphorus compounds and a potential target for cognitive enhancing drugs. Mol. Pharmacol. 2000, 58, 577–583. [Google Scholar] [PubMed]
- Harmat, V.; Domokos, K.; Menyhárd, D.K.; Palló, A.; Szeltner, Z.; Szamosi, I.; Beke-Somfai, T.; Náray-Szabó, G.; Polgár, L. Structure and catalysis of acylaminoacyl peptidase: Closed and open subunits of a dimer oligopeptidase. J. Biol. Chem. 2011, 286, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Bartlam, M.; Wang, G.; Yang, H.; Gao, R.; Zhao, X.; Gao, S.; Feng, Y.; Rao, Z. Crystal structure of an acylpeptide hydrolase/esterase from Aeropyrum pernix K1 in complex with p-nitrophenyl phosphate. Structure 2004, 12, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, G.; Langella, E.; Gogliettino, M.; Saviano, M.; Pocsfalvia, G.; Rossid, M. A novel class of protease targets of phosphatidylethanolamine-binding proteins (PEBP): A study of the acylpeptide hydrolase and the PEBP inhibitor from the archaeon Sulfolobus solfataricus. Mol. Biosyst. 2010, 6, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Scaloni, A.; Jones, WM.; Pospischil, M.; Sassa, S.; Schneewind, O.; Popowicz, A.M.; Bossa, F.; Graziano, S.L.; Manning, J.M. Deficiency of acylpeptide hydrolase in small-cell lung carcinoma cell lines. J. Lab. Clin. Med. 1992, 120, 546–552. [Google Scholar] [PubMed]
- Erlandsson, R.; Boldog, F.; Persson, B.; Zabarousky, E.R.; Allikmets, R.L.; Su megi, J.; Klein, G.; Joernvall, H. The gene from the short arm of chromosome 3, at D3F15S2, frequently deleted in renal cell carcinoma, encodes acylpeptide hydrolase. Oncogene 1991, 6, 1293–1295. [Google Scholar] [PubMed]
- Gass, J.; Khosla, C. Prolyl endopeptidases. Cell. Mol. Life Sci. 2007, 64, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, J.S.; Kozarich, J.W. Prolyl peptidases: A serine protease subfamily with high potential for drug discovery. Curr. Opin. Chem. Biol. 2003, 7, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Gallo, M.A.; Lawryk, N.J. Organic Phosphorus Pesticides; Academic Press: San Diego, CA, USA, 1991; pp. 918–919. [Google Scholar]
- Yang, G.Y.; Bai, A.X.; Gao, L.; Zhang, Z.M.; Zheng, B.S.; Feng, Y. Glu88 in the non-catalytic domain of acylpeptide hydrolase plays dual roles: Charge neutralization for enzymatic activity and formation of salt bridge for thermodynamic stability. Biochim. Biophys. Acta 2009, 1794, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, E.; Renzetti, G. Coupled motions during dynamics reveal a tunnel toward the active site regulated by the N-terminal α-helix in an acylaminoacyl peptidase. J. Mol. Graph. Model. 2012, 38, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, E.; Renzetti, G.; Tiberti, M. Mechanisms of intramolecular communication in a hyperthermophilic acylaminoacyl peptidase: A molecular dynamics investigation. PLoS One 2012, 7, e35686. [Google Scholar] [CrossRef] [PubMed]
- Fuxreiter, M.; Magyar, C.; Juhász, T.; Szeltner, Z.; Polgár, L.; Simon, I. Flexibility of prolyl oligopeptidase: Molecular dynamics and molecular framework analysis of the potential substrate pathways. PROTEINS: Struct. Funct. Bioinform. 2005, 60, 504–512. [Google Scholar] [CrossRef]
- Marzinek, J.K.; Bond, P.J.; Lian, G.; Zhao, Y.; Han, L.; Noro, M.G.; Pistikopoulos, E.N.; Mantalaris, A. Free energy predictions of ligand binding to an α-helix using steered molecular dynamics and umbrella sampling simulations. J. Chem. Inf. Model. 2014, 54, 2093–2104. [Google Scholar] [CrossRef] [PubMed]
- Soares, T.A.; Osman, M.A.; Straatsma, T.P. Molecular dynamics of organophosphorous hydrolases bound to the nerve agent soman. J. Chem. Theory Comput. 2007, 3, 1569–1579. [Google Scholar] [CrossRef]
- Gomes, D.E.B.; Lins, R.D.; Pascutti, P.G.; Lei, C.; Soares, T.A. Conformational variability of organophosphorus hydrolase upon soman and paraoxon binding. J. Phys. Chem. B 2011, 115, 15389–15398. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, D.L.; Dicapua, F.M. Free energy via molecular simulation: Applications to chemical and biomolecular systems. Annu. Rev. Biophys. Chem. 1989, 18, 431–492. [Google Scholar] [CrossRef]
- Deng, Y.Q.; Roux, B. Computations of standard binding free energies with molecular dynamics simulations. J. Phys. Chem. B 2009, 113, 2234–2246. [Google Scholar] [CrossRef] [PubMed]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.H.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W.; et al. Calculating structures and free energies of complex molecules: Combining molecular mechanics and continuum models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, B.; Gerber, P.; Schulz-Gasch, T.; Stahl, M. Validation and use of the MM-PBSA approach for drug discovery. J. Med. Chem. 2005, 48, 4040–4048. [Google Scholar] [CrossRef] [PubMed]
- Hansson, T.; Marelius, J.; Aqvist, J. Ligand binding affinity prediction by linear interaction energy methods. J. Comput. Aided Mol. Des. 1998, 12, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Laio, A.; Parinello, M. Escaping free-energy minima. Proc. Natl. Acad. Sci. USA 2014, 99, 12562–12566. [Google Scholar] [CrossRef]
- Kokubo, H.; Tanaka, T.; Okamoto, Y. Ab initio prediction of protein-ligand binding structures by replica-exchange umbrella sampling simulations. J. Comput. Chem. 2011, 32, 2810–2821. [Google Scholar] [CrossRef]
- Torrien, G.M.; Valleau, J.P. Monte-Carlo free-energy estimates using non-Boltzmann sampling applicatio to subcritical Lennard-Jones fluid. Chem. Phys. Lett. 1974, 28, 578–581. [Google Scholar] [CrossRef]
- Buch, I.; Sadiq, S.K.; de Fabritiis, G. Optimized potential of mean force calculations for standard binding free energies. J. Chem. Theory Comput. 2011, 7, 1765–1772. [Google Scholar] [CrossRef]
- Woo, H.J.; Roux, B. Calculation of absolute protein-ligand binding free energy from computer simulations. Proc. Natl. Acad. Sci. USA 2005, 102, 6825–6830. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Families of serine peptidases. Methods Enzymol. 1994, 244, 19–61. [Google Scholar] [PubMed]
- Mukherjee, S.; Balius, T.E.; Rizzo, R.C. Docking validation resources: Protein family and ligand flexibility experiments. J. Chem. Inf. Model. 2010, 50, 1986–2000. [Google Scholar] [CrossRef] [PubMed]
- Brozell, S.R.; Mukherjee, S.; Balius, T.E.; Roe, D.R.; Case, D.A.; Rizzo, R.C. Evaluation of DOCK 6 as a pose generation and database enrichment tool. J. Comput. Aided Mol. Des. 2012, 26, 749–773. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Chen, L.C.; Li, Q.; Zheng, M.Z.; Liu, J.S. Computational study on substrate specificity of a novel cysteine Protease 1 Precursor from Zea mays. Int. J. Mol. Sci. 2014, 15, 10459–10478. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed]
- Norgan, A.P.; Coffman, P.K.; Kocher, J.P.; Katzmann, D.J.; Sosa, C.P. Multilevel parallelization of AutoDock 4.2. J. Cheminform. 2011, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.S.; Robertson, D.H.; Brooks, C.L., III; Michal, V. Detailed analysis of grid-based molecular docking: A case study of CDOCKER-A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Benlloch, C.; Roldán, E.; Chang, Y.; Shi, T. Regularized linearization for quantum nonlinear optical cavities: Application to degenerate optical parametric oscillators. Opt. Express. 2014, 22, 24010–24023. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Chen, F. Multiwfn: A multifunctional wavefunction analyzer. J. Comput. Chem. 2012, 33, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1996, 8, 127–134. [Google Scholar] [CrossRef]
- Fulop, V.; Szeltner, Z.; Polgar, L. Catalysis of serine oligopeptidasesis controlled by a gating filter mechanism. EMBO Rep. 2000, 1, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, H.; Kyono, K.; Higashiyama, Y.; Fukushima, C.; Shima, H.; Sugiyama, S.; Inaka, K.; Yamamoto, A.; Shimizu, R. The structure and function of human dipeptidyl peptidase IV, possessing a unique eight-bladed b-propeller fold. Biochem. Biophys. Res. Commun. 2003, 302, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Lindorff-Larsen, K.; Maragakis, P.; Piana, S.; Eastwood, M.P.; Dror, R.O.; Shaw, D.E. Systematic validation of protein force fields against experimental data. PLoS One 2012, 7, e32131. [Google Scholar] [CrossRef] [PubMed]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An overview of the Amber biomolecular simulation package. WIREs Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. A linear constraint solver for molecular simulations. J. Comp. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Miyamoto, S.; Kollman, P.A. An analytical version of the SHAKE and RATTLE algorithm for rigid water models. J. Comput. Chem. 1992, 13, 952–962. [Google Scholar] [CrossRef]
- Van Aalten, D.M.; Bywater, R.; Findlay, J.B.; Hendlich, M.; Hooft, R.W.; Vriend, G. PRODRG, a program for generating molecular toppologies and uniquemolecular descriptors from coordinates of small molecules. J. Comput. Aided Mol. Des. 1996, 10, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Piana, S.; Lindorff-Larsen, K.; Shaw, D.E. How robust are protein-folding simulations with respect to force field parameterization? Biophys. J. 2011, 100, L47–L49. [Google Scholar] [CrossRef] [PubMed]
- Laberge, M.; Yonetani, T. Molecular dynamics simulations of hemoglobin A in different states and bound to DPG: Effector-linked perturbation of tertiary conformations and HbA concerted dynamics. Biophys. J. 2008, 94, 2737–2751. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Hu, J.P.; Li, K.S.; Tian, X.H.; Chang, S. Molecular dynamics simulations of DNA-free and DNA-bound TAL effectors. PLoS One 2013, 8, e76045. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, H.X.; Yao, X.J. Understanding the molecular basis of MK2-p38a signaling complex assembly: Insights into protein-protein interaction by molecular dynamics and free energy studies. Mol. Biosyst. 2012, 8, 2106–2118. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.P.; He, H.Q.; Jiao, X.; Chang, S. Understanding the folding and stability of a designed WW domain protein with replica exchange molecular dynamics simulations. Mol. Simul. 2013, 39, 828–836. [Google Scholar] [CrossRef]
- Case, D.A.; Cheatham, T.E., III; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, H.; Zhou, Z.; Wang, D.; Guan, S.; Han, W. Molecular Dynamics Simulations of Acylpeptide Hydrolase Bound to Chlorpyrifosmethyl Oxon and Dichlorvos. Int. J. Mol. Sci. 2015, 16, 6217-6234. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16036217
Jin H, Zhou Z, Wang D, Guan S, Han W. Molecular Dynamics Simulations of Acylpeptide Hydrolase Bound to Chlorpyrifosmethyl Oxon and Dichlorvos. International Journal of Molecular Sciences. 2015; 16(3):6217-6234. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16036217
Chicago/Turabian StyleJin, Hanyong, Zhenhuan Zhou, Dongmei Wang, Shanshan Guan, and Weiwei Han. 2015. "Molecular Dynamics Simulations of Acylpeptide Hydrolase Bound to Chlorpyrifosmethyl Oxon and Dichlorvos" International Journal of Molecular Sciences 16, no. 3: 6217-6234. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16036217