
The WRKY Transcription Factor Genes in Eggplant (Solanum melongena L.) and Turkey Berry (Solanum torvum Sw.)

Abstract
:1. Introduction
2. Results and Discussion
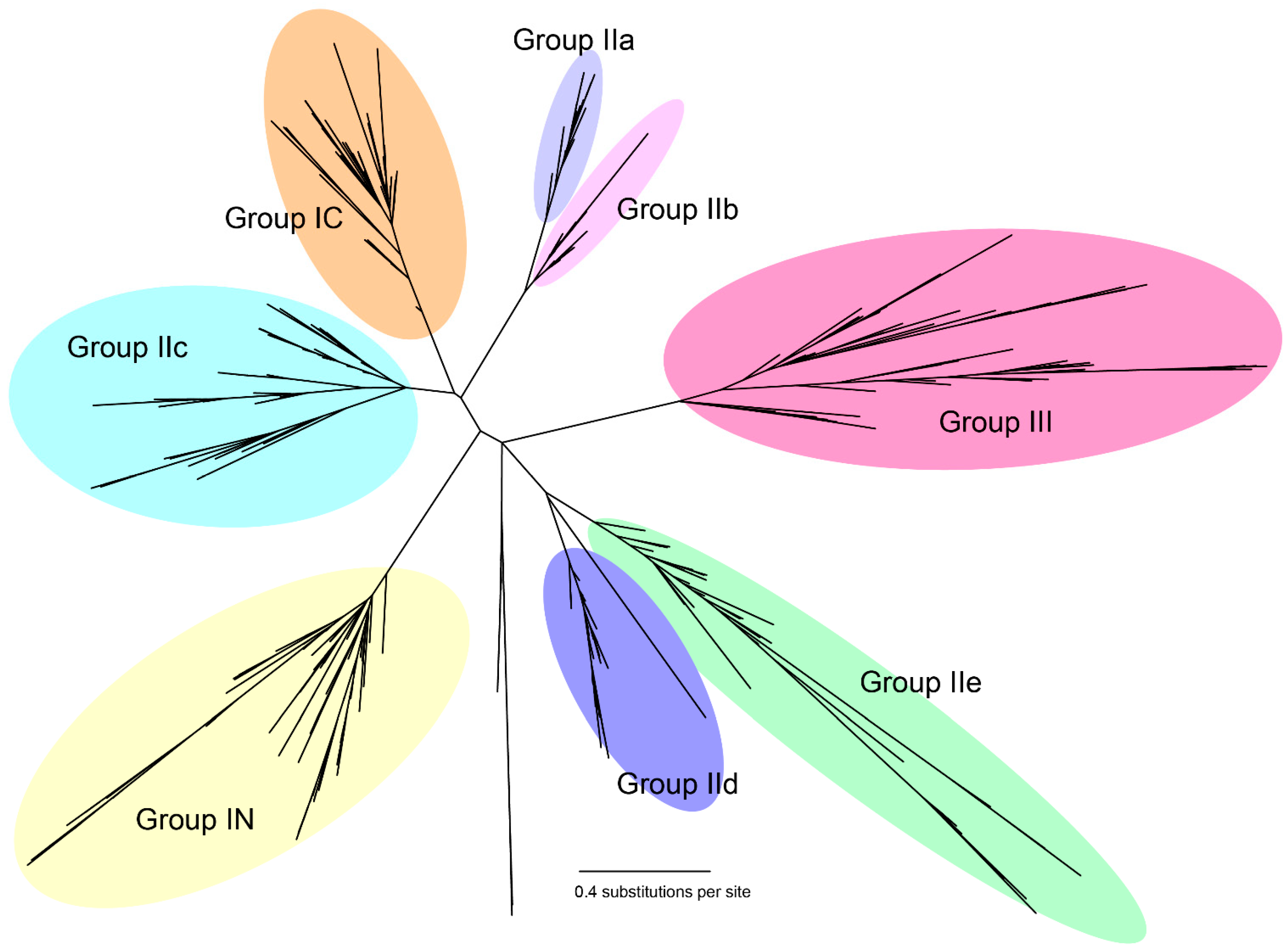
2.1. Identification of the Eggplant and Turkey Berry WRKY Gene Families

{kind=link}
{kind=link}
{kind=link}
| Species | Complete WRKY * | Partial WRKY | I | IIa | IIb | IIc | IId | IIe | III | Unassigned |
|---|---|---|---|---|---|---|---|---|---|---|
| A. trichopoda | 29 | 3 | 6 | 2 | 4 | 4 | 3 | 4 | 4 | 2 |
| S. polyrhiza | 34 | 7 | 9 | 1 | 4 | 6 | 6 | 4 | 4 | 0 |
| A. thaliana | 72 | 0 | 14 | 3 | 8 | 17 | 7 | 9 | 13 | 1 |
| P. trichocarpa | 101 | 1 | 22 | 5 | 9 | 25 | 13 | 14 | 10 | 3 |
| V. vinifera | 57 | 2 | 12 | 3 | 8 | 15 | 6 | 6 | 6 | 1 |
| S. lycopersicum | 80 | 1 | 15 | 5 | 8 | 16 | 6 | 18 | 11 | 1 |
| S. tuberosum | 77 | 8 | 14 | 5 | 6 | 16 | 7 | 15 | 14 | 0 |
| C. annuum | 59 | 3 | 15 | 4 | 6 | 11 | 5 | 8 | 9 | 1 |
| S. melongena | 44 | 6 | 12 | 2 | 3 | 8 | 7 | 5 | 7 | 0 |
| S. torvum | 53 | 9 | 13 | 4 | 3 | 10 | 7 | 6 | 10 | 0 |
2.2. Phylogenetic Analysis and Classification of the WRKY Gene Family

| Species | Total Group I | With Two WRKY Domains | With One WRKY Domain | |||
|---|---|---|---|---|---|---|
| Complete Group I | Partial IC | Partial IN | IN | IC | ||
| A. trichopoda | 6 | – | – | – | – | – |
| S. polyrhiza | 9 | 6 | 1 | – | – | 2 |
| A. thaliana | 14 | 12 | 1 | – | – | 1 |
| P. trichocarpa | 22 | 21 | 1 | – | – | – |
| V. vinifera | 12 | 11 | – | – | – | 1 |
| S. lycopersicum | 15 | 15 | – | – | – | – |
| S. tuberosum | 14 | 12 | 1 | – | 1 | – |
| C. annuum | 15 | 13 | 1 | – | 1 | – |
| S. melongena | 12 | 4 | 1 | – | 3 | 4 |
| S. torvum | 13 | 6 | 1 | – | 4 | 2 |
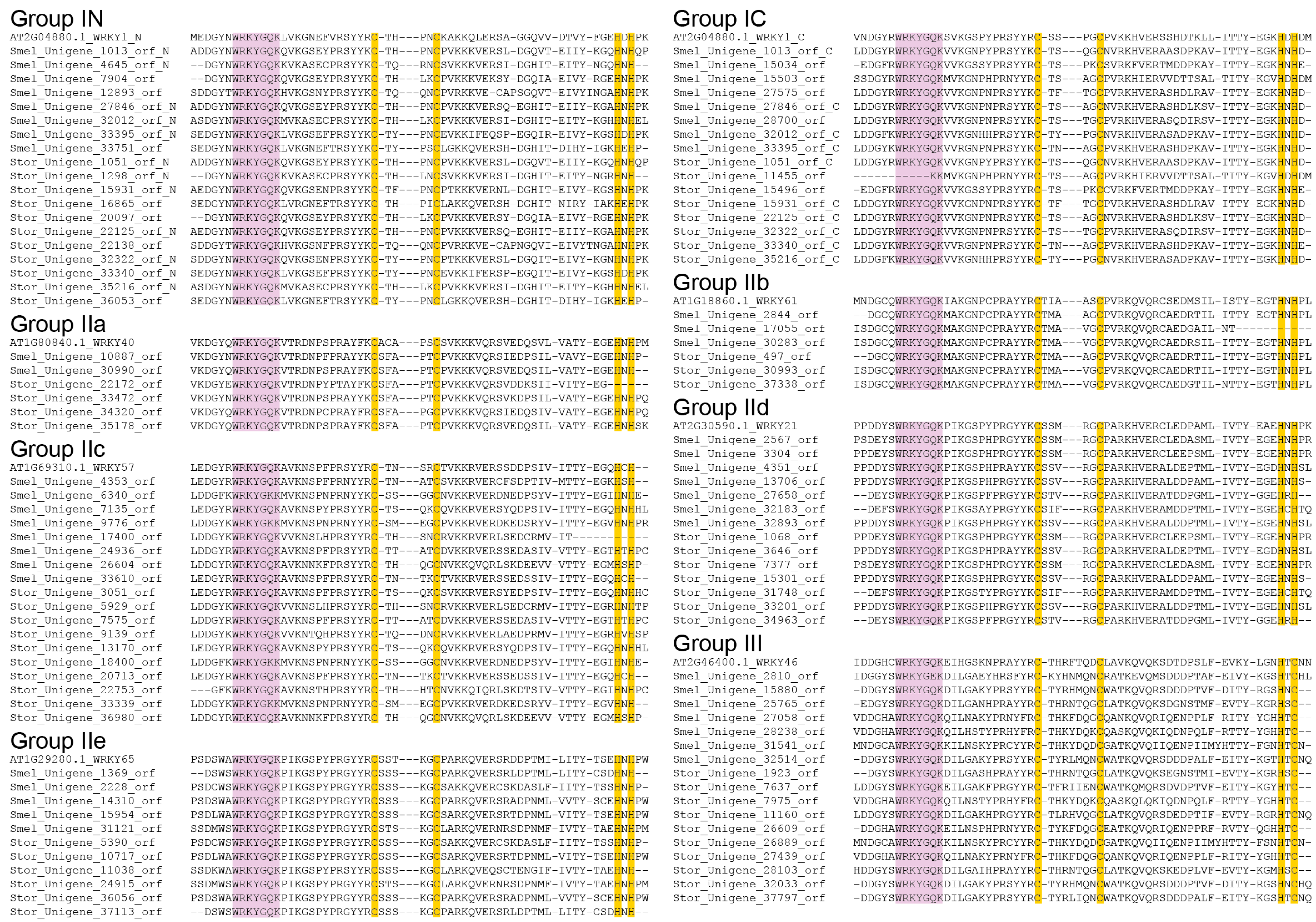
2.3. Multiple Sequence Alignment of SmelWRKYs and StorWRKYs

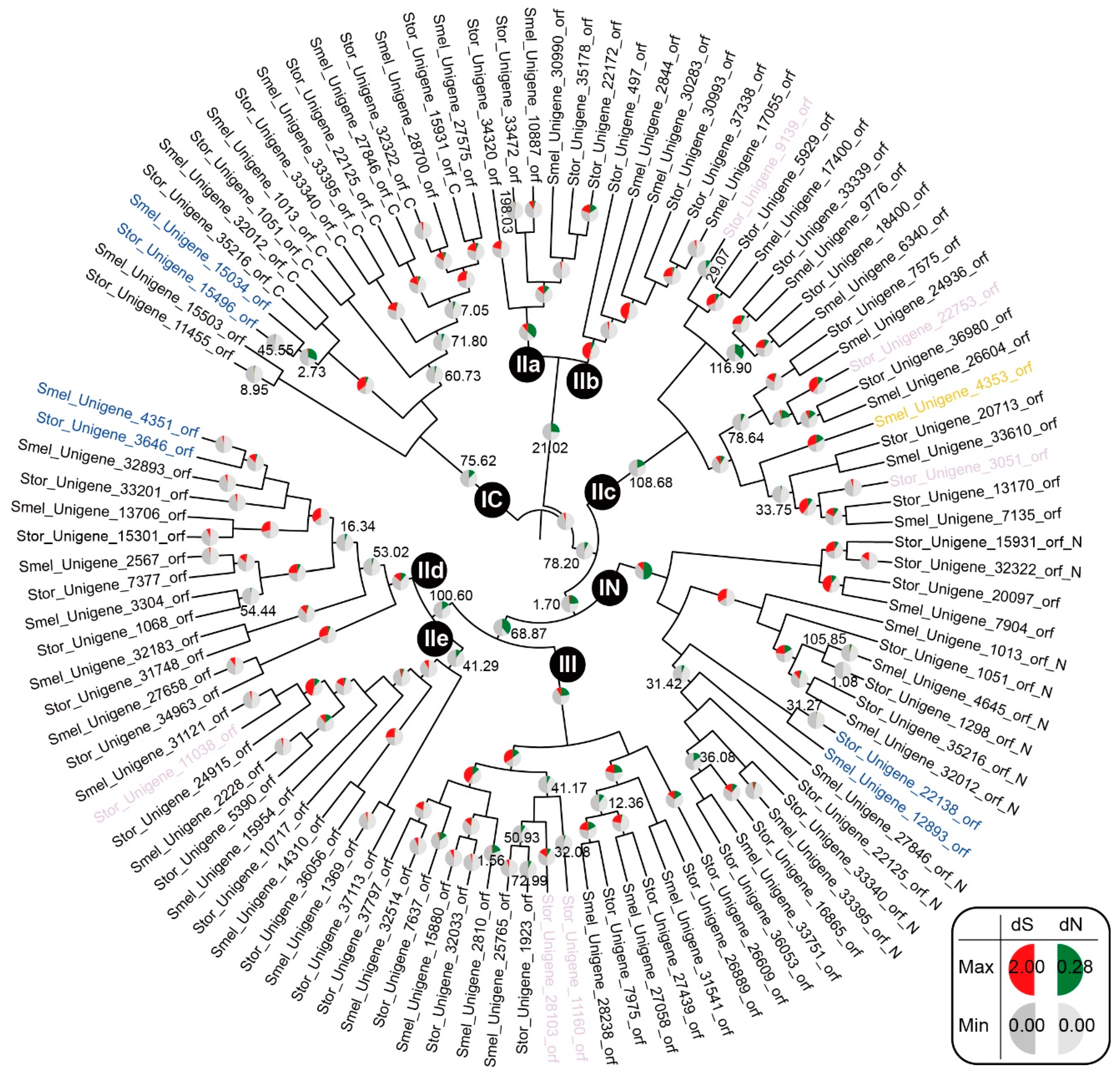
2.4. Evolutionary Selective Pressure in SmelWRKYs and StorWRKYs

| Group | ι (Foreground) | ι (Background) | 2Δι | p-Value a | Positive Selection Sites b |
|---|---|---|---|---|---|
| IN | −4987.059421 | −4987.867786 | 1.61673 | 2.50 × 10−1 | 8E *, 20N *, 34P *, 36F * |
| IC | −4990.205486 | −4990.433633 | 0.45629 | 1.02 × 10−1 | 35A * |
| IIa | −4991.663923 | −4991.663923 | 0.00000 | 5.00 × 10−1 | none |
| IIb | −4991.663924 | −4991.663923 | −2.00 × 10−6 | – | None |
| IIc | −4987.458167 | −4989.190540 | 3.46475 | 3.14 × 10−2 | 6G ** |
| IId | −4991.663923 | −4991.663923 | 0.00000 | 5.00 × 10−1 | none |
| IIe | −4988.849412 | −4990.664638 | 3.63045 | 2.84 × 10−2 | none |
| III | −4985.941961 | −4988.138760 | 4.39360 | 1.80 × 10−2 | 5L * |
2.5. Predicted Roles of SmelWRKY and StorWRKY Orthologs
| Cluster | A. thaliana | S. melongena | S. torvum |
|---|---|---|---|
| Cluster1 | AtWRKY46 a | Smel_Unigene_32514_orf | Stor_Unigene_37797_orf |
| AtWRKY53, AtWRKY41 | Smel_Unigene_15880_orf | Stor_Unigene_32033_orf | |
| Cluster6 | AtWRKY7 b | Smel_Unigene_13706_orf | Stor_Unigene_15301_orf |
| Smel_Unigene_32893_orf | Stor_Unigene_33201_orf | ||
| Cluster7 | AtWRKY20 b | Smel_Unigene_33395_orf | Stor_Unigene_33340_orf |
| Cluster11 | AtWRKY72 b | – | – |
| Cluster15 | AtWRKY75 a | Smel_Unigene_26604_orf | Stor_Unigene_36980_orf |
| Cluster18 | AtWRKY8, AtWRKY28 a | Smel_Unigene_7135_orf | Stor_Unigene_13170_orf |
| Cluster19 | AtWRKY6 a | Smel_Unigene_30283_orf | Stor_Unigene_30993_orf |
| Cluster20 | AtWRKY48 b | Smel_Unigene_24936_orf | Stor_Unigene_7575_orf |
| Cluster21 | AtWRKY23 | Smel_Unigene_4353_orf | – |
| Cluster22 | AtWRKY13 | – | Stor_Unigene_9139_orf |
| Cluster26 | AtWRKY24, AtWRKY43, AtWRKY56 | – | Stor_Unigene_22753_orf |
| Cluster28 | – | – | Stor_Unigene_3051_orf |
| Cluster32 | AtWRKY29 | – | Stor_Unigene_11038_orf |
| Cluster35 | – | Smel_Unigene_10887_orf | Stor_Unigene_33472_orf |
| Cluster39 | AtWRKY30 | – | Stor_Unigene_11160_orf |
| Cluster40 | AtWRKY62, AtWRKY67, AtWRKY38, AtWRKY66, AtWRKY64, AtWRKY63 a | – | – |
| Cluster59 | – | – | Stor_Unigene_28103_orf |
| Cluster71 | – | Smel_Unigene_12893_orf | Stor_Unigene_22138_orf |
| Cluster72 | – | Smel_Unigene_15034_orf | Stor_Unigene_15496_orf |
| Cluster73 | – | Smel_Unigene_4351_orf | Stor_Unigene_3646_orf |
3. Experimental Section
3.1. Dataset Collection
3.2. Structural Annotation of Eggplant and Turkey Berry Unigenes
3.3. WRKY Gene Identification
3.4. WRKY Gene Classification
3.5. Selection Pressure Analyses
3.6. Identification of Gene Orthologous Groups
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Xu, X.; Pan, S.; Cheng, S.; Zhang, B.; Mu, D.; Ni, P.; Zhang, G.; Yang, S.; Li, R.; Wang, J. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Zouine, M.; Latché, A.; Rousseau, C.; Regad, F.; Pech, J.-C.; Philippot, M.; Bouzayen, M.; Delalande, C.; Frasse, P.; Schiex, T. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collonnier, C.; Fock, I.; Kashyap, V.; Rotino, G.; Daunay, M.; Lian, Y.; Mariska, I.; Rajam, M.; Servaes, A.; Ducreux, G. Applications of biotechnology in eggplant. Plant Cell Tissue Organ Cult. 2001, 65, 91–107. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Fukuoka, H.; Arao, T.; Ohyama, A.; Nunome, T.; Miyatake, K.; Negoro, S. Gene expression analysis in cadmium-stressed roots of a low cadmium-accumulating solanaceous plant, solanum torvum. J. Exp. Bot. 2010, 61, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Gousset, C.; Collonnier, C.; Mulya, K.; Mariska, I.; Rotino, G.L.; Besse, P.; Servaes, A.; Sihachakr, D. Solanum torvum, as a useful source of resistance against bacterial and fungal diseases for improvement of eggplant (S. Melongena L.). Plant Sci. 2005, 168, 319–327. [Google Scholar] [CrossRef]
- Garibaldi, A.; Minuto, A.; Gullino, M. Verticillium wilt incited by verticillium dahliae in eggplant grafted on solanum torvum in italy. Plant Dis. 2005, 89, 777. [Google Scholar]
- Daunay, M.; Lester, R.; Laterrot, H. The use of wild species for the genetic improvement of brinjal eggplant (Solanum melongena) and tomato (Lycopersicon esculentum). Solanaceae III Taxon. Chem. Evol. 1991, 27, 389–413. [Google Scholar]
- Guri, A.; Sink, K. Interspecific somatic hybrid plants between eggplant (Solanum melongena) and solanum torvum. Theor. Appl. Genet. 1988, 76, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J.; Snyder, A. Eggplant (Solanum melongena L.). In Agrobacterium Protocols; Humana Press: Clifton, NJ, USA, 2006; pp. 439–448. [Google Scholar]
- Yang, X.; Cheng, Y.-F.; Deng, C.; Ma, Y.; Wang, Z.-W.; Chen, X.-H.; Xue, L.-B. Comparative transcriptome analysis of eggplant (Solanum melongena L.) and turkey berry (Solanum torvum Sw.): Phylogenomics and disease resistance analysis. BMC Genomics 2014, 15, 412. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The wrky superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Ulker, B.; Somssich, I.E. WRKY transcription factors: From DNA binding towards biological function. Curr. Opin. Plant Biol. 2004, 7, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. Transparent testa glabra2, a trichome and seed coat development gene of arabidopsis, encodes a wrky transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef]
- Jiang, W.; Yu, D. Arabidopsis WRKY2 transcription factor mediates seed germination and postgermination arrest of development by abscisic acid. BMC Plant Biol. 2009, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Zhang, Z.L.; Hanzlik, S.; Cook, E.; Shen, Q.J. Salicylic acid inhibits gibberellin-induced alpha-amylase expression and seed germination via a pathway involving an abscisic-acid-inducible wrky gene. Plant Mol. Biol. 2007, 64, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Zhang, Z.; Laux, T. Transcriptional activation of arabidopsis axis patterning genes wox8/9 links zygote polarity to embryo development. Dev. Cell 2011, 20, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Laun, T.; Zimmermann, P.; Zentgraf, U. Targets of the wrky53 transcription factor and its role during leaf senescence in arabidopsis. Plant Mol. Biol. 2004, 55, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta 2012, 1819, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression profiles of 7000 arabidopsis genes under drought, cold and high-salinity stresses using a full-length cdna microarray. Plant J. 2002, 31, 279–292. [Google Scholar] [PubMed]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; D’Angelo, C.; Bornberg-Bauer, E.; Kudla, J.; Harter, K. The atgenexpress global stress expression data set: Protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Chen, Z.; Liu, Y.; Zhang, H.; Zhang, M.; Liu, Q.; Hong, X.; Zhu, J.K.; Gong, Z. Abo3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in arabidopsis. Plant J. 2010, 63, 417–429. [Google Scholar] [CrossRef]
- Ramamoorthy, R.; Jiang, S.Y.; Kumar, N.; Venkatesh, P.N.; Ramachandran, S. A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments. Plant Cell Physiol. 2008, 49, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Schmelzer, E.; Hahlbrock, K.; Somssich, I.E. Early nuclear events in plant defence signalling: Rapid gene activation by wrky transcription factors. EMBO J. 1999, 18, 4689–4699. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.P.; Somssich, I.E. The role of wrky transcription factors in plant immunity. Plant Physiol. 2009, 150, 1648–1655. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, N.; Yoshioka, H. Post-translational regulation of WRKY transcription factors in plant immunity. Curr. Opin. Plant Biol. 2012, 15, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2013, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.H.; Fischer, N.M.; Harter, K.; Kohlbacher, O.; Wanke, D. Elucidating the evolutionary conserved DNA-binding specificities of wrky transcription factors by molecular dynamics and in vitro binding assays. Nucleic Acids Res. 2013, 41, 9764–9778. [Google Scholar] [CrossRef] [PubMed]
- Machens, F.; Becker, M.; Umrath, F.; Hehl, R. Identification of a novel type of WRKY transcription factor binding site in elicitor-responsive cis-sequences from arabidopsis thaliana. Plant Mol. Biol. 2014, 84, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. Interproscan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z.; Cao, S.; Liu, Y. Genome-wide analysis of WRKY transcription factors in solanum lycopersicum. Mol. Genet. Genomics 2012, 287, 495–513. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Zhang, L.B.; Guo, D.; Li, C.Z.; Peng, S.Q. Identification and expression profiles of the WRKY transcription factor family in ricinus communis. Gene 2012, 503, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Jiang, W.; Zhang, Y.; Yu, H.; Mao, Z.; Gu, X.; Huang, S.; Xie, B. Genome-wide analysis of wrky gene family in cucumis sativus. BMC Genomics 2011, 12, 471. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Schluttenhofer, C.; Pattanaik, S.; Patra, B.; Yuan, L. Analyses of catharanthus roseus and arabidopsis thaliana wrky transcription factors reveal involvement in jasmonate signaling. BMC Genomics 2014, 15, 502. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Xu, H.; Xiao, S.; Qin, Y.; Li, Y.; Yan, Y.; Hu, Y. The large soybean (Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups. BMC Plant Biol. 2013, 13, 148. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, P.; Nan, Z. The WRKY transcription factor genes in lotus japonicus. Int. J. Genomics 2014, 2014. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.R.; Nan, J.; Liang, Y.H.; Mao, P.; Lu, L.; Li, L.; Wei, C.; Lai, L.; Li, Y.; Su, X.D. DNA binding mechanism revealed by high resolution crystal structure of arabidopsis thaliana wrky1 protein. Nucleic Acids Res. 2007, 35, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.K.; Baek, K.H.; Park, J.M.; Yi, S.Y.; Yu, S.H.; Kamoun, S.; Choi, D. Capsicum annuum WRKY protein cawrky1 is a negative regulator of pathogen defense. New Phytol. 2008, 177, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Brunk, B.P.; Chen, F.; Gao, X.; Harb, O.S.; Iodice, J.B.; Shanmugam, D.; Roos, D.S.; Stoeckert, C.J. Using OrthoMCL to assign proteins to OrthoMCL-DB groups or to cluster proteomes into new ortholog groups. Curr. Protoc. Bioinform. 2011. [Google Scholar] [CrossRef]
- Scarpeci, T.E.; Zanor, M.I.; Mueller-Roeber, B.; Valle, E.M. Overexpression of atwrky30 enhances abiotic stress tolerance during early growth stages in arabidopsis thaliana. Plant Mol. Biol. 2013, 83, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, W.; Karimi, M.; Wieczorek, K.; Van de Cappelle, E.; Wischnitzki, E.; Grundler, F.; Inze, D.; Beeckman, T.; Gheysen, G. A role for atwrky23 in feeding site establishment of plant-parasitic nematodes. Plant Physiol. 2008, 148, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagnaresi, P.; Sala, T.; Irdani, T.; Scotto, C.; Lamontanara, A.; Beretta, M.; Rotino, G.; Sestili, S.; Cattivelli, L.; Sabatini, E. Solanum torvum responses to the root-knot nematode meloidogyne incognita. BMC Genomics 2013, 14, 540. [Google Scholar] [CrossRef] [PubMed]
- Tacconi, R. Reproduction of heterodera schachtii on selected crops. Nematol. Mediterr. 1993, 21, 9–12. [Google Scholar]
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell Online 2002, 14, S153–S164. [Google Scholar]
- Park, C.Y.; Lee, J.H.; Yoo, J.H.; Moon, B.C.; Choi, M.S.; Kang, Y.H.; Lee, S.M.; Kim, H.S.; Kang, K.Y.; Chung, W.S.; et al. WRKY group IId transcription factors interact with calmodulin. FEBS Lett. 2005, 579, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, K.K.; Atamian, H.S.; Kaloshian, I.; Eulgem, T. WRKY72-type transcription factors contribute to basal immunity in tomato and arabidopsis as well as gene-for-gene resistance mediated by the tomato R gene Mi-1. Plant J. 2010, 63, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, R.; Fonstein, M.; D’Souza, M.; Pusch, G.D.; Maltsev, N. The use of gene clusters to infer functional coupling. Proc. Natl. Acad. Sci. USA 1999, 96, 2896–2901. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, J.; Lin, G.; Wang, A.; Wang, Z.; Lu, G. Overexpression of AtWRKY28 and AtWRKY75 in Arabidopsis enhances resistance to oxalic acid and Sclerotinia sclerotiorum. Plant Cell Rep. 2013, 32, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.P.; Sarma, B.K.; Singh, D.P.; Maurya, S.; Mishra, P.K.; Singh, H.B. Effects of exudate depletion on sclerotial development of sclerotinia sclerotiorum and of oxalic acid on the synthesis of phenolic acids in egg plant (Solanum melongena L.). J. Plant Dis. Prot. 2004, 111, 371–379. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Boeckmann, B.; Bairoch, A.; Apweiler, R.; Blatter, M.-C.; Estreicher, A.; Gasteiger, E.; Martin, M.J.; Michoud, K.; O’Donovan, C.; Phan, I. The swiss-prot protein knowledgebase and its supplement trembl in 2003. Nucleic Acids Res. 2003, 31, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Iseli, C.; Jongeneel, C.V.; Bucher, P. Estscan: A Program for Detecting, Evaluating, and Reconstructing Potential Coding Regions in Est Sequences; ISMB: San Diego, CA, USA, 1999; pp. 138–148. [Google Scholar]
- Birney, E.; Clamp, M.; Durbin, R. Genewise and genomewise. Genome Res. 2004, 14, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of phyml 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Deng, C.; Zhang, Y.; Cheng, Y.; Huo, Q.; Xue, L. The WRKY Transcription Factor Genes in Eggplant (Solanum melongena L.) and Turkey Berry (Solanum torvum Sw.). Int. J. Mol. Sci. 2015, 16, 7608-7626. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16047608
Yang X, Deng C, Zhang Y, Cheng Y, Huo Q, Xue L. The WRKY Transcription Factor Genes in Eggplant (Solanum melongena L.) and Turkey Berry (Solanum torvum Sw.). International Journal of Molecular Sciences. 2015; 16(4):7608-7626. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16047608
Chicago/Turabian StyleYang, Xu, Cao Deng, Yu Zhang, Yufu Cheng, Qiuyue Huo, and Linbao Xue. 2015. "The WRKY Transcription Factor Genes in Eggplant (Solanum melongena L.) and Turkey Berry (Solanum torvum Sw.)" International Journal of Molecular Sciences 16, no. 4: 7608-7626. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16047608