
Exposure of Tumor-Associated Macrophages to Apoptotic MCF-7 Cells Promotes Breast Cancer Growth and Metastasis

Abstract
:1. Introduction
2. Results
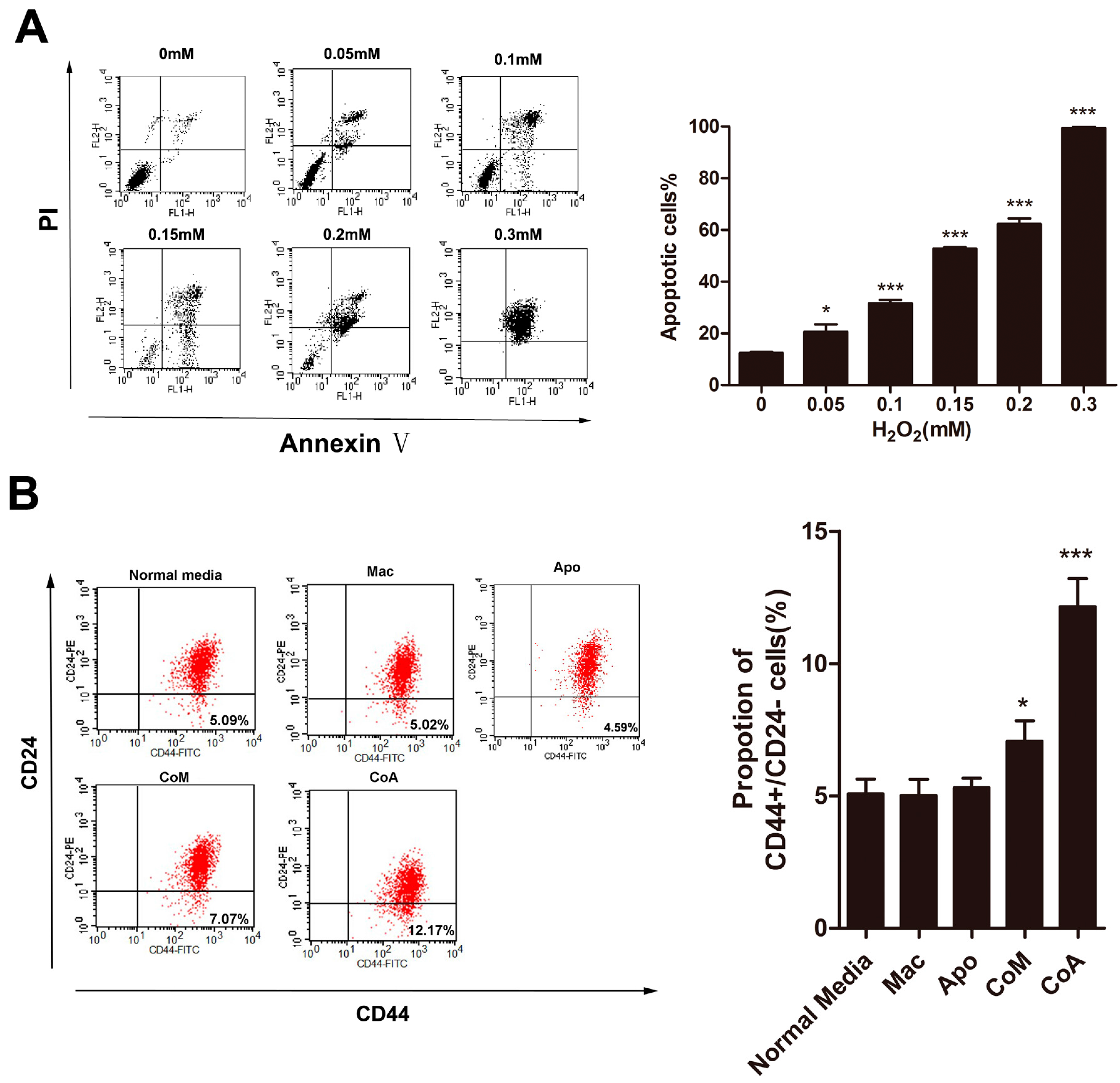
2.1. Macrophages Could Increase the Proportion of CD44+/CD24− Cancer Stem Cells after Co-Culture with Apoptotic MCF-7 Cells

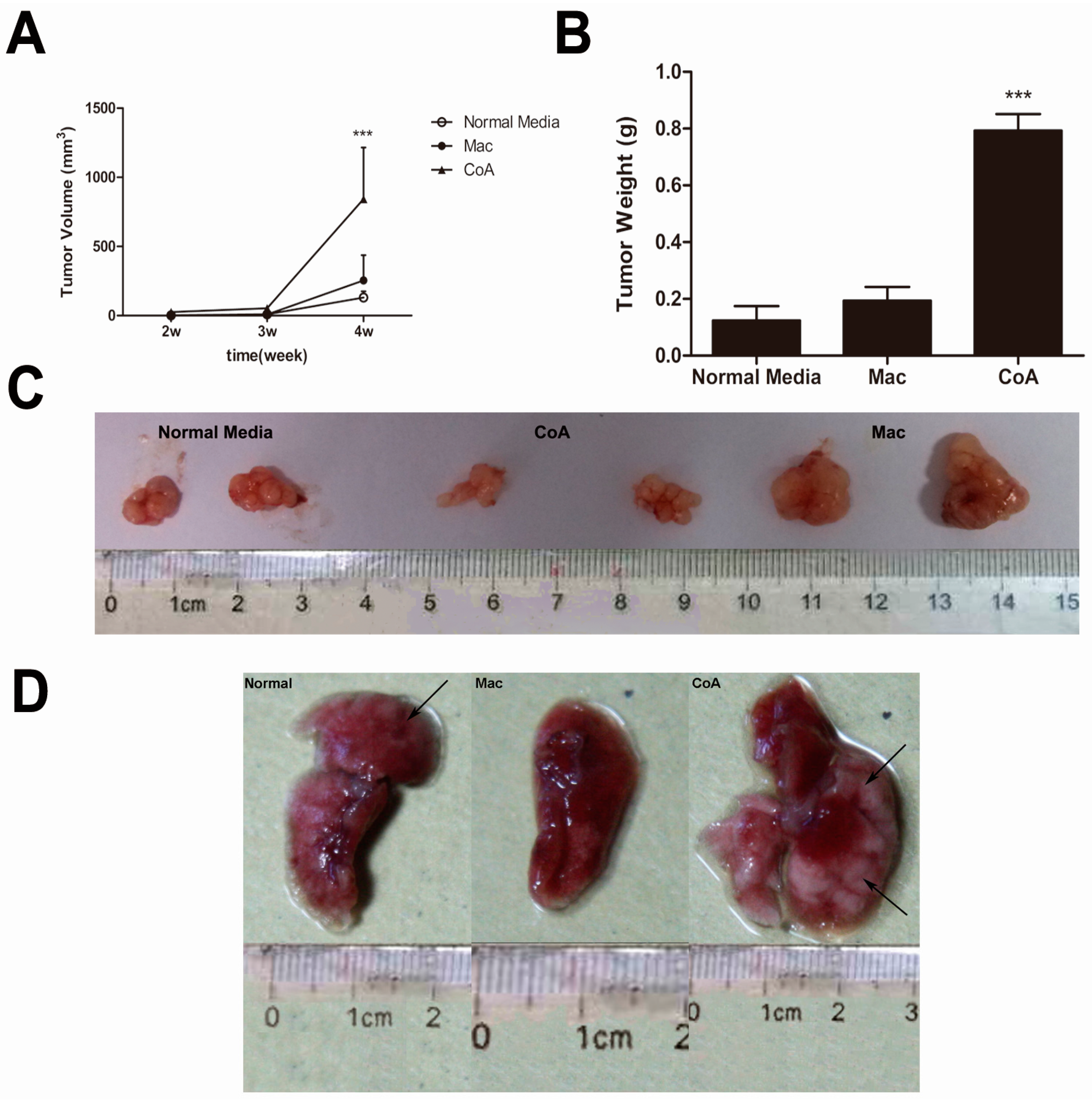
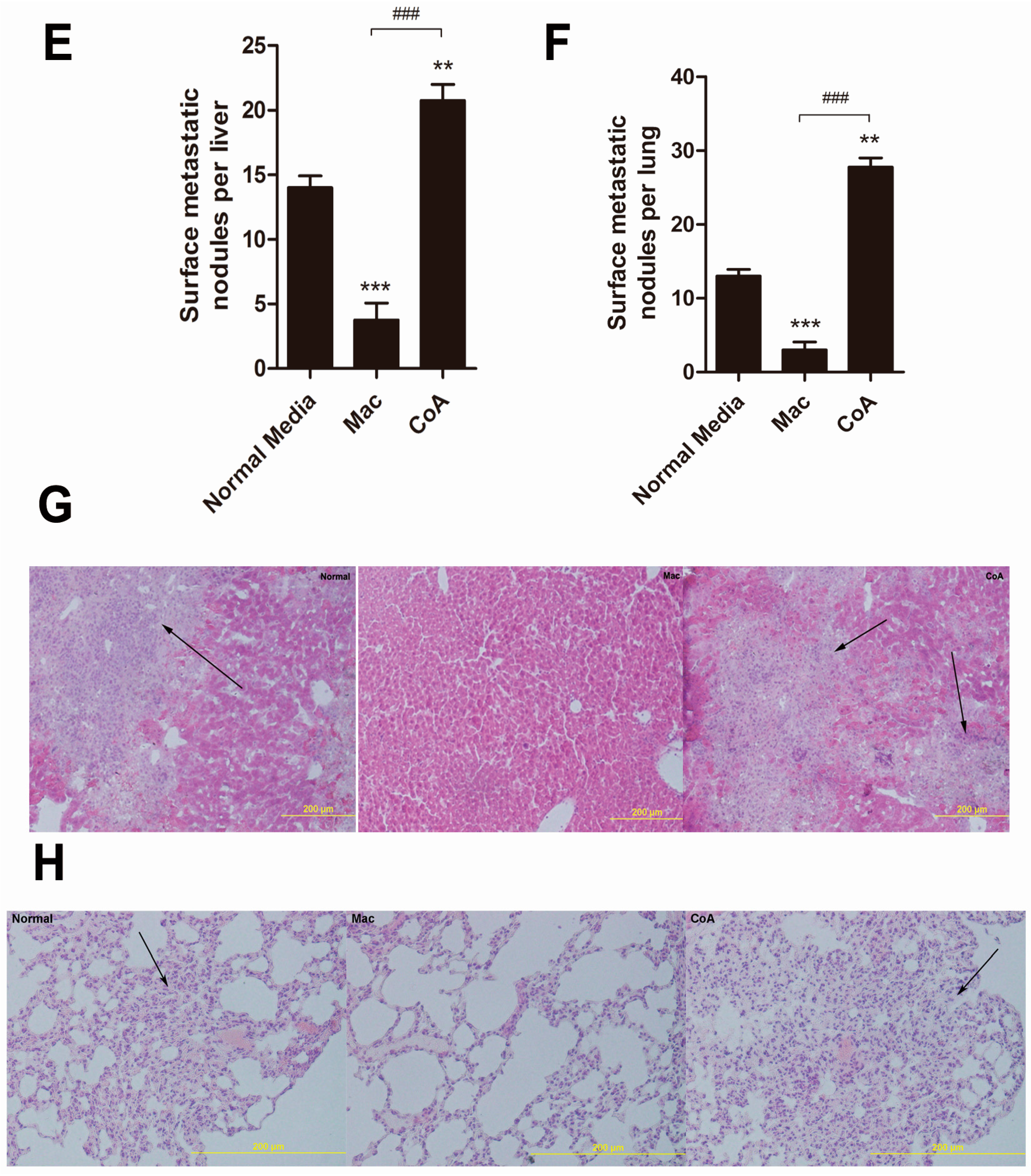
2.2. MCF-7 Cells Cultured in Conditioned Media from a Co-Culture of Macrophages and Apoptotic MCF-7 Show Increased Tumor Growth and Metastatic Ability in Vivo


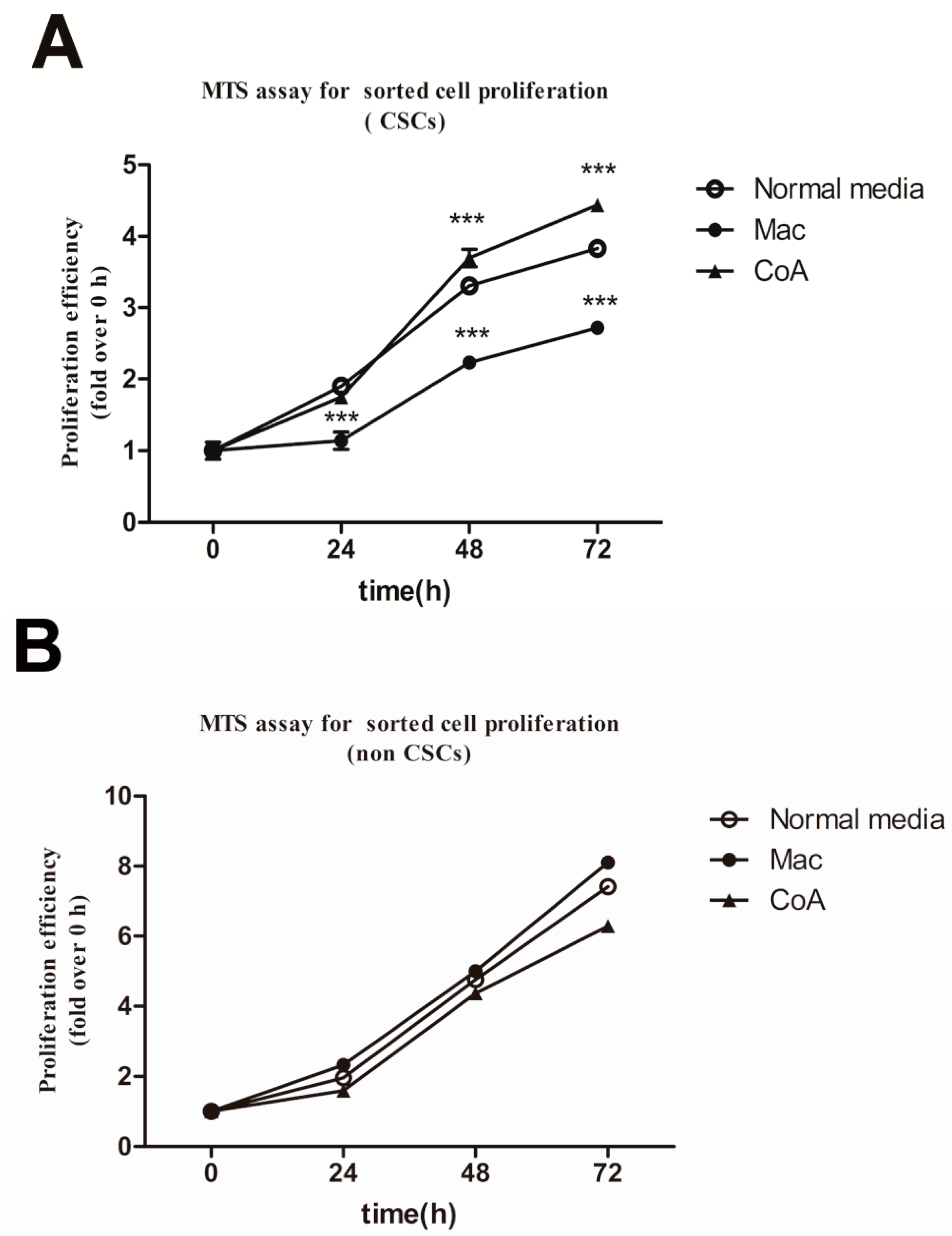
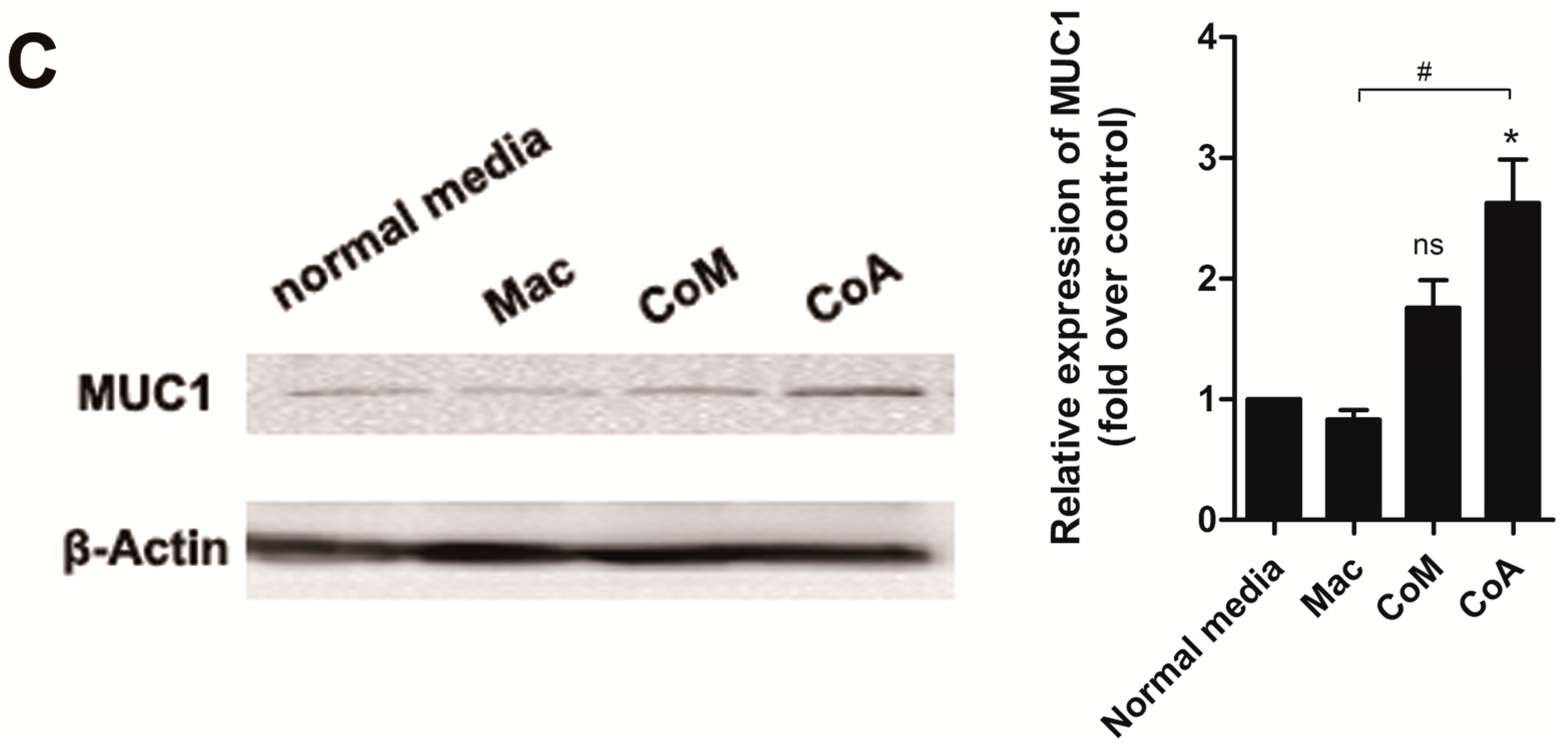
2.3. Macrophage Co-Culture with Apoptotic MCF-7 Cells Promotes the Proliferative Ability of CD44+/CD24− Cancer Stem-Like Cells and Up-Regulates the Expression of Mucin 1


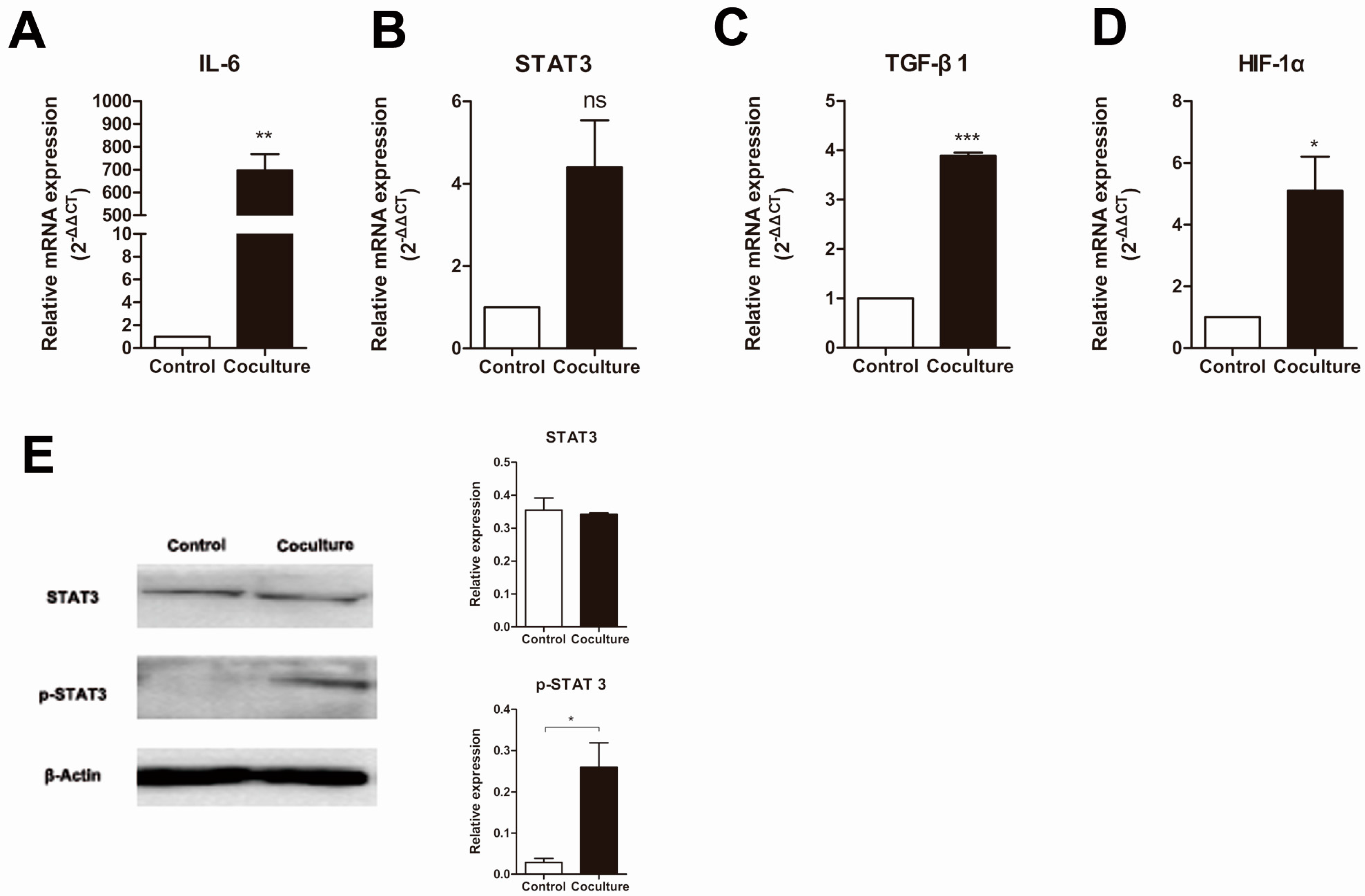
2.4. Macrophage Interleukin 6 (IL-6)/Signal Transducers and Activators of Transcription 3 (STAT3) Signaling Pathway Is Activated after Co-Culture with Apoptotic Cancer Cells

3. Discussion
4. Experimental Section
4.1. Mice
4.2. Mice Peritoneal Macrophages Isolation and Culture
4.3. Generation of Apoptotic MCF-7
4.4. Co-Culture Experiment and the Preparation of Conditioned Media
4.5. Flow Cytometric Analysis and Fluorescence-Activated Cell Sorting (FACS)
4.6. Cell Proliferation Assay
4.7. Nude Mice Xenografts Assay and Tumor Metastasis Assay
4.8. Isolation of Total RNA and Quantitative RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Tm (°C) |
|---|---|---|
| IL-6 | F-CAACGATGATGCACTTGCAGA R-CTCCAGGTAGCTATGGTACTCCAGA | 64 |
| STAT3 | F-TGCACCTGATCACCTTCGAGAC R-CCCAAGCATTTGGCATCTGAC | 68 |
| TGF-β1 | F-TACGGCAGTGGCTGAACCAA R-CGGTTCATGTCATGGATGGTG | 68 |
| HIF-1α | F-GGACGATGAACATCAAGTCAGCA R-AGGAATGGGTTCACAAATCAGCA | 68 |
| β-actin | F-CATCCGTAAAGACCTCTATGCCAAC R-ATGGAGCCACCGATCCACA | 60 |
4.9. Western Blot Analysis
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Carey, L.A.; Metzger, R.; Dees, E.C.; Collichio, F.; Sartor, C.I.; Ollila, D.W.; Klauber-DeMore, N.; Halle, J.; Sawyer, L.; Moore, D.T.; et al. American Joint Committee on cancer tumor-node-metastasis stage after neoadjuvant chemotherapy and breast cancer outcome. J. Natl. Cancer Inst. 2005, 97, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Kerbel, R.S. Reappraising antiangiogenic therapy for breast cancer. Breast 2011, 20, S56–S60. [Google Scholar] [CrossRef]
- Steeg, P.S. Tumor metastasis: Mechanistic insights and clinical challenges. Nat. Med. 2006, 12, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Allavena, P.; Mantovani, A. Cancer related inflammation: The macrophage connection. Cancer Lett. 2008, 267, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.Y.; Pollard, J.W. Macrophages: Modulators of breast cancer progression. Novartis Found. Symp. 2004, 256, 158–168, discussion 168–172, 259–269. [Google Scholar] [PubMed]
- Lin, E.Y.; Nguyen, A.V.; Russell, R.G.; Pollard, J.W. Colony-stimulating factor 1 promotes progression of mammary tumors to malignancy. J. Exp. Med. 2001, 193, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.Y.; Gouon-Evans, V.; Nguyen, A.V.; Pollard, J.W. The macrophage growth factor CSF-1 in mammary gland development and tumor progression. J. Mammary Gland Biol. Neoplasia 2002, 7, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.; Deng, Y.; Im, J.H.; Muschel, R.J.; Zou, Y.; Li, J.; Lang, R.A.; Pollard, J.W. A distinct macrophage population mediates metastatic breast cancer cell extravasation, establishment and growth. PLoS ONE 2009, 4, e6562. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liao, D.; Chen, C.; Liu, Y.; Chuang, T.H.; Xiang, R.; Markowitz, D.; Reisfeld, R.A.; Luo, Y. Tumor-associated macrophages regulate murine breast cancer stem cells through a novel paracrine EGFR/Stat3/Sox-2 signaling pathway. Stem Cells 2013, 31, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Leek, R.D.; Harris, A.L. Tumor-associated macrophages in breast cancer. J. Mammary Gland Biol. Neoplasia 2002, 7, 177–189. [Google Scholar] [PubMed]
- Wyckoff, J.; Wang, W.; Lin, E.Y.; Wang, Y.; Pixley, F.; Stanley, E.R.; Graf, T.; Pollard, J.W.; Segall, J.; Condeelis, J. A paracrine loop between tumor cells and macrophages is required for tumor cell migration in mammary tumors. Cancer Res. 2004, 64, 7022–7029. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Jia, P.; Zhao, H.; Hu, C.; Yang, X. MicroRNA-26a overexpression protects RGC-5 cells against H2O2-induced apoptosis. Biochem. Biophys. Res. Commun. 2015, 460, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.; Malone, C.; Chumsri, S.; Burger, A.M.; McDonnell, S. Characterisation of breast cancer cell lines and establishment of a novel isogenic subclone to study migration, invasion and tumourigenicity. Clin. Exp. Metastasis 2008, 25, 549–557. [Google Scholar] [PubMed]
- Dontu, G.; Liu, S.; Wicha, M.S. Stem cells in mammary development and carcinogenesis: Implications for prevention and treatment. Stem Cell Rev. 2005, 1, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Zhang, Y.; Zhang, X.; Lei, Z.; Hu, R.; Li, H.; Mao, Y.; Wang, X.; Irwin, D.M.; Niu, G.; et al. Chemotherapy Induced Apoptosis May Activate Cancer Stem-Like Cells (CD44+/CD24−) in MCF-7 by Upregulating Mucin 1 and EpCAM. Peking University: Beijing, China, Unpublished work. 2015. [Google Scholar]
- Da Silva, S.D.; Hier, M.; Mlynarek, A.; Kowalski, L.P.; Alaoui-Jamali, M.A. Recurrent oral cancer: Current and emerging therapeutic approaches. Front. Pharmacol. 2012, 3, 149. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.A.; Zafarana, G.; Ishkanian, A.S.; Milosevic, M.; Thoms, J.; Have, C.L.; Malloff, C.A.; Lam, W.L.; Squire, J.A.; Pintilie, M.; et al. NKX3.1 haploinsufficiency is prognostic for prostate cancer relapse following surgery or image-guided radiotherapy. Clin. Cancer Res. 2012, 18, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.A.; Ndabahaliye, A.; Lim, P.K.; Milton, R.; Rameshwar, P. Challenges in the development of future treatments for breast cancer stem cells. Breast Cancer 2010, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Swann, J.B.; Vesely, M.D.; Silva, A.; Sharkey, J.; Akira, S.; Schreiber, R.D.; Smyth, M.J. Demonstration of inflammation-induced cancer and cancer immunoediting during primary tumorigenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.W. Macrophages define the invasive microenvironment in breast cancer. J. Leukoc. Biol. 2008, 84, 623–630. [Google Scholar] [PubMed]
- Condeelis, J.; Pollard, J.W. Macrophages: Obligate partners for tumor cell migration, invasion, and metastasis. Cell 2006, 124, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.H.; Yu, C.T.; Wu, C.Y.; Lee, M.J.; Lee, W.H.; Wang, L.S.; Lin, S.M.; Fu, J.F.; Lee, K.Y.; Yen, T.H. Tumor-associated macrophages in stage IIIA pN2 non-small cell lung cancer after neoadjuvant chemotherapy and surgery. Am. J. Transl. Res. 2014, 6, 593–603. [Google Scholar] [PubMed]
- Lanciotti, M.; Masieri, L.; Raspollini, M.R.; Minervini, A.; Mari, A.; Comito, G.; Giannoni, E.; Carini, M.; Chiarugi, P.; Serni, S. The role of M1 and M2 macrophages in prostate cancer in relation to extracapsular tumor extension and biochemical recurrence after radical prostatectomy. Biomed. Res. Int. 2014, 2014, 486798. [Google Scholar] [CrossRef] [PubMed]
- Tham, M.; Khoo, K.; Yeo, K.P.; Kato, M.; Prevost-Blondel, A.; Angeli, V.; Abastado, J.P. Macrophage depletion reduces postsurgical tumor recurrence and metastatic growth in a spontaneous murine model of melanoma. Oncotarget 2015. [Google Scholar] [PubMed]
- Hikita, S.T.; Kosik, K.S.; Clegg, D.O.; Bamdad, C. MUC1* mediates the growth of human pluripotent stem cells. PLoS ONE 2008, 3, e3312. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, K.; Shen, H.; Finn, O.J. MCF7 side population cells with characteristics of cancer stem/progenitor cells express the tumor antigen MUC1. Cancer Res. 2008, 68, 2419–2426. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.X.; Cui, Y.X.; Long, Z.J.; Dai, Z.M.; Lin, J.Y.; Liang, Y.; Zheng, F.M.; Zeng, Y.X.; Liu, Q. Pluripotency-associated genes in human nasopharyngeal carcinoma CNE-2 cells are reactivated by a unique epigenetic sub-microenvironment. BMC Cancer 2010, 10, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinushi, M.; Chiba, S.; Yoshiyama, H.; Masutomi, K.; Kinoshita, I.; Dosaka-Akita, H.; Yagita, H.; Takaoka, A.; Tahara, H. Tumor-associated macrophages regulate tumorigenicity and anticancer drug responses of cancer stem/initiating cells. Proc. Natl. Acad. Sci. USA 2011, 108, 12425–12430. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Wei, J.; Kong, L.Y.; Wang, Y.; Priebe, W.; Qiao, W.; Sawaya, R.; Heimberger, A.B. Glioma cancer stem cells induce immunosuppressive macrophages/microglia. Neuro-oncology 2010, 12, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Naugler, W.E.; Karin, M. The wolf in sheep’s clothing: The role of interleukin-6 in immunity, inflammation and cancer. Trends Mol. Med. 2008, 14, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Wu, T.; Zhang, H.W.; Lu, N.; Hu, R.; Wang, Y.J.; Zhao, L.; Chen, F.H.; Wang, X.T.; You, Q.D.; et al. HIF-1α is critical for hypoxia-mediated maintenance of glioblastoma stem cells by activating Notch signaling pathway. Cell Death Differ. 2012, 19, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Philip, B.; Ito, K.; Moreno-Sanchez, R.; Ralph, S.J. HIF expression and the role of hypoxic microenvironments within primary tumours as protective sites driving cancer stem cell renewal and metastatic progression. Carcinogenesis 2013, 34, 1699–1707. [Google Scholar] [CrossRef] [PubMed]
- Muraoka-Cook, R.S.; Kurokawa, H.; Koh, Y.; Forbes, J.T.; Roebuck, L.R.; Barcellos-Hoff, M.H.; Moody, S.E.; Chodosh, L.A.; Arteaga, C.L. Conditional overexpression of active transforming growth factor β1 in vivo accelerates metastases of transgenic mammary tumors. Cancer Res. 2004, 64, 9002–9011. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.M.; Jing, Y.Y.; Yu, G.F.; Kou, X.R.; Ye, F.; Gao, L.; Li, R.; Zhao, Q.D.; Yang, Y.; Lu, Z.H.; et al. Tumor-associated macrophages promote cancer stem cell-like properties via transforming growth factor-β1-induced epithelial-mesenchymal transition in hepatocellular carcinoma. Cancer Lett. 2014, 352, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Zhao, E.; Kryczek, I.; Vatan, L.; Sadovskaya, A.; Ludema, G.; Simeone, D.M.; Zou, W.; Welling, T.H. Tumor-associated macrophages produce interleukin 6 and signal via STAT3 to promote expansion of human hepatocellular carcinoma stem cells. Gastroenterology 2014, 147, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Mano, Y.; Aishima, S.; Fujita, N.; Tanaka, Y.; Kubo, Y.; Motomura, T.; Taketomi, A.; Shirabe, K.; Maehara, Y.; Oda, Y. Tumor-associated macrophage promotes tumor progression via STAT3 signaling in hepatocellular carcinoma. Pathobiology 2013, 80, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Li, Y.; Wang, Y.; Zhao, H.; Zhang, J.; Chai, H.; Tang, T.; Yue, J.; Guo, A.M.; Yang, J. Downregulation of COX-2 and CYP 4A signaling by isoliquiritigenin inhibits human breast cancer metastasis through preventing anoikis resistance, migration and invasion. Toxicol. Appl. Pharmacol. 2014, 280, 10–20. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, N.; Zhang, Y.; Zhang, X.; Lei, Z.; Hu, R.; Li, H.; Mao, Y.; Wang, X.; Irwin, D.M.; Niu, G.; et al. Exposure of Tumor-Associated Macrophages to Apoptotic MCF-7 Cells Promotes Breast Cancer Growth and Metastasis. Int. J. Mol. Sci. 2015, 16, 11966-11982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160611966
Zhou N, Zhang Y, Zhang X, Lei Z, Hu R, Li H, Mao Y, Wang X, Irwin DM, Niu G, et al. Exposure of Tumor-Associated Macrophages to Apoptotic MCF-7 Cells Promotes Breast Cancer Growth and Metastasis. International Journal of Molecular Sciences. 2015; 16(6):11966-11982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160611966
Chicago/Turabian StyleZhou, Na, Yizhuang Zhang, Xuehui Zhang, Zhen Lei, Ruobi Hu, Hui Li, Yiqing Mao, Xi Wang, David M. Irwin, Gang Niu, and et al. 2015. "Exposure of Tumor-Associated Macrophages to Apoptotic MCF-7 Cells Promotes Breast Cancer Growth and Metastasis" International Journal of Molecular Sciences 16, no. 6: 11966-11982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160611966