
Transcriptional Profiling of Hydrogen Production Metabolism of Rhodobacter capsulatus under Temperature Stress by Microarray Analysis

Abstract
:1. Introduction
2. Results and Discussion
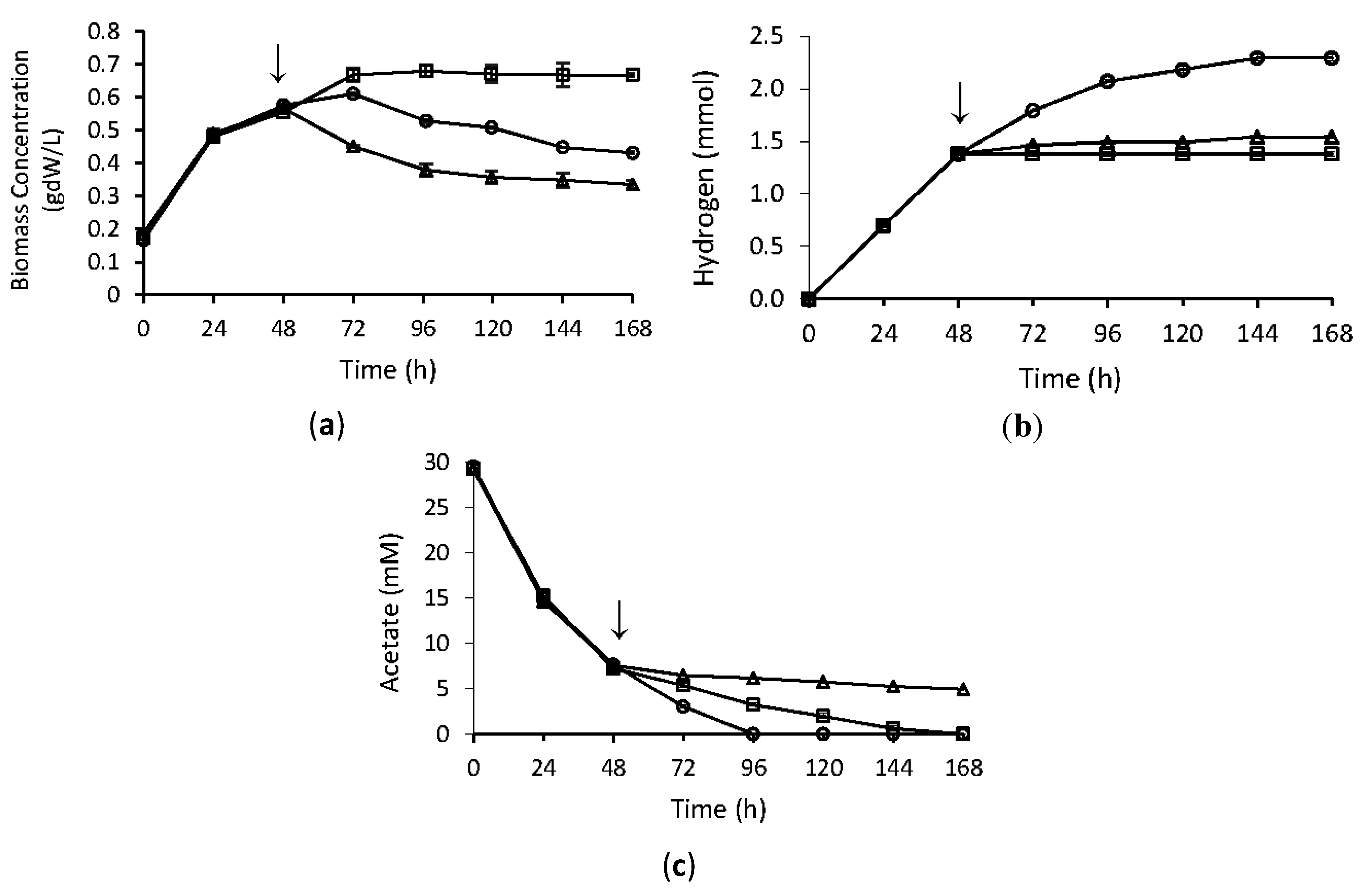
2.1. Physiological Analyses

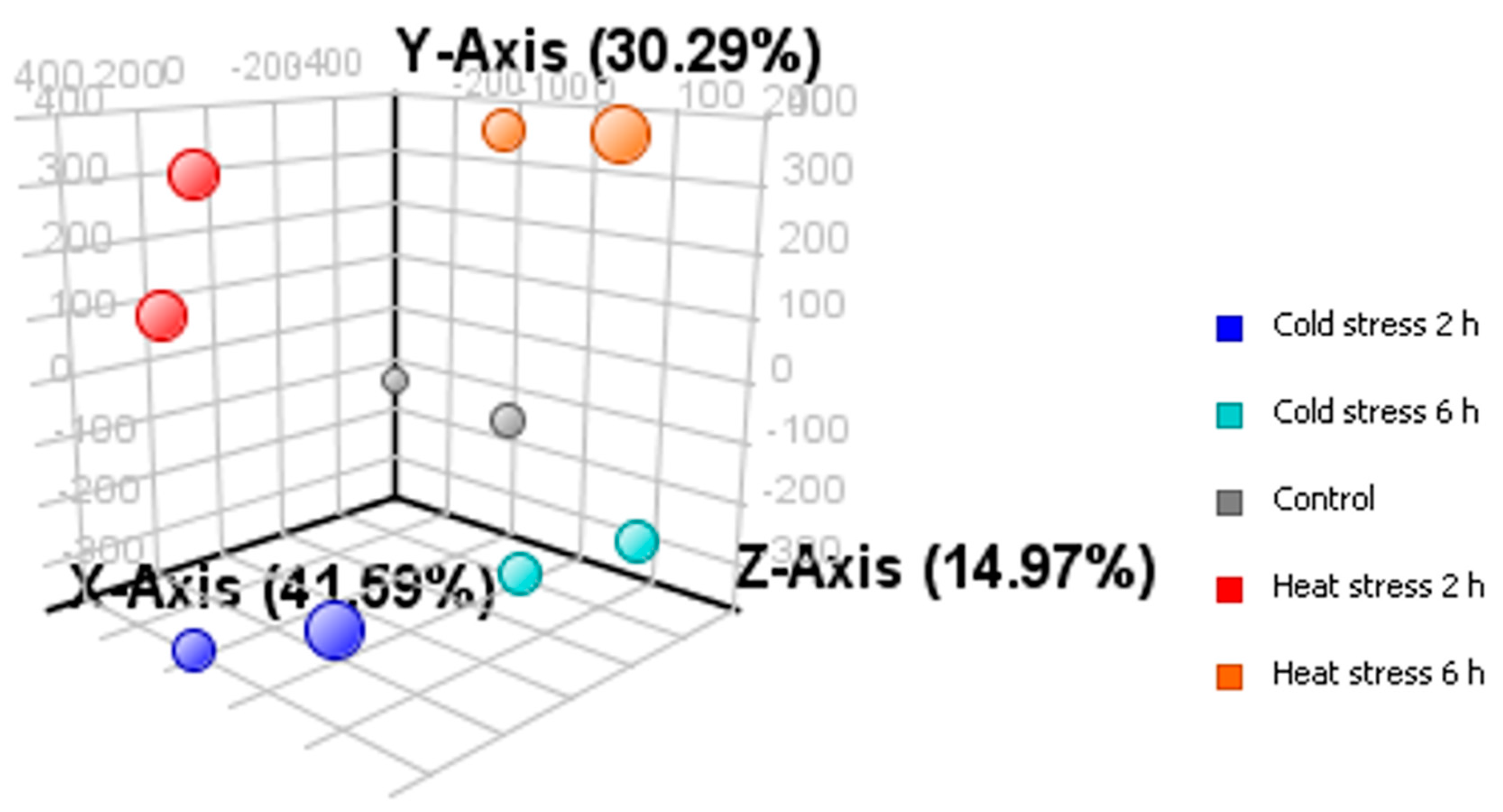
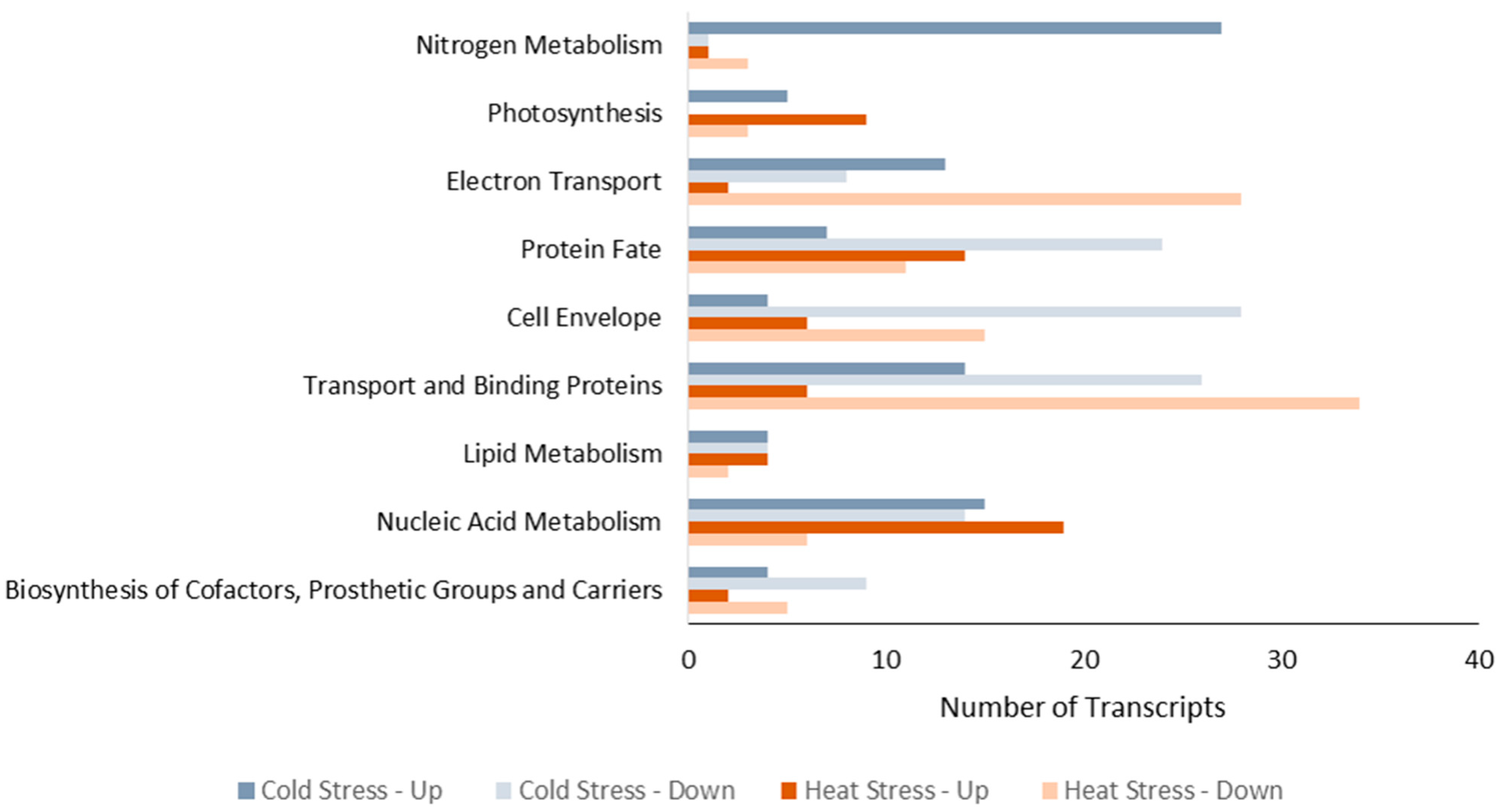
2.2. Microarray Analysis


{kind=link}
{kind=link}
{kind=link}
| Samples | Correlation Coefficients between Duplicates |
|---|---|
| Control | 0.975 |
| Cold stress 2 h | 0.959 |
| Cold stress 6 h | 0.985 |
| Heat stress 2 h | 0.967 |
| Heat stress 6 h | 0.987 |
| Gene | Transcript Change with Real-Time qPCR | Transcript Level Change with Microarray | Forward Primer (5′>3′) | Reverse Primer (3′>5′) |
|---|---|---|---|---|
| 16S rRNA | 1.0 | 1.0 | GCTAGTAATCGCGTAACAGCA | CAGTCACTGAGCCTACCGT |
| atpF | +2.50 ± 0.23 | +3.57 | ACGTTCCTGCTTGTTGCTCT | TCGAGGGAACCTTGAACTTG |
| pufM | −1.43 ± 0.02 | −3.10 | CACCATCGGTGTGTGGTACT | AGACACCACCCTGTTTCAGC |
| oppA | −2.58 ± 0.12 | −3.20 | AGGAACTGCTCAAACCGATG | GTCCTTGTAGTCCGCAAACC |

| Functional Group | Probe Set ID | Gene Symbol | Description | Fold Change | p Value | |
|---|---|---|---|---|---|---|
| 2 h | 6 h | |||||
| Nitrogen Metabolism | RCAP_rcc00566_s_at | nifB | Nitrogenase cofactor biosynthesis protein NifB | +6.317 | +2.487 | 0.0204 |
| RCAP_rcc00571_at | nifD | Nitrogenase molybdenum-iron protein α chain | +59.112 | +38.480 | 0.0176 | |
| RCAP_rcc03280_at | nifE | Nitrogenase molybdenum-iron cofactor biosynts protein NifE | +3.069 | +1.223 | 0.0089 | |
| RCAP_rcc00572_at | nifH | Nitrogenase iron protein | +99.953 | +44.111 | 0.006 | |
| RCAP_rcc00570_at | nifK | Nitrogenase molybdenum-iron protein β chain | +29.376 | +16.530 | 0.003 | |
| RCAP_rcc03279_at | nifN | Nitrogenase molybdenum-iron cofactor biosynthesis protein NifN | +6.598 | +1.957 | 0.0038 | |
| RCAP_rcc00586_at | anfD | Nitrogenase iron-iron protein, α subunit | +23.486 | +13.126 | 0.0301 | |
| RCAP_rcc00587_at | anfG | Nitrogenase iron-iron protein, delta subunit | +37.180 | +22.194 | 0.0379 | |
| RCAP_rcc00585_at | anfH | Nitrogenase iron protein | +33.189 | +20.210 | 0.0873 | |
| RCAP_rcc01674_at | glnA | Glutamine synthetase | +2.249 | +2.212 | 0.0027 | |
| RCAP_rcc01673_at | glnB | Nitrogen regulatory protein P-II | +2.756 | +2.093 | 0.0355 | |
| Photosynthesis | RCAP_rcc01830_at | fba | Fructose-bisphosphate aldolase | +2.056 | +3.441 | 0.0137 |
| RCAP_rcc01834_at | fbp | Fructose-bisphosphatase | +2.748 | +2.564 | 0.0988 | |
| RCAP_rcc00744_at | atpF | ATP synthase F0, B subunit | +3.575 | +1.640 | 0.0535 | |
| Electron Transport | RCAP_rcc03284_at | fdxN | Ferredoxin I | +23.668 | +10.214 | 0.0093 |
| RCAP_rcc03285_at | fdxC | Ferredoxin IV | +13.582 | +2.455 | 0.0000 | |
| RCAP_rcc03287_at | rnfA | Electron transport complex protein RnfA | +20.417 | +5.874 | 0.0033 | |
| RCAP_rcc03288_at | rnfB | Electron transport complex protein RnfB | +9.373 | +1.972 | 0.0018 | |
| RCAP_rcc03289_at | rnfC | Electron transport complex protein RnfC | +17.068 | +7.526 | 0.0036 | |
| Electron Transport | RCAP_rcc03290_at | rnfD | Electron transport complex protein RnfD | +7.219 | +4.282 | 0.0042 |
| RCAP_rcc03292_at | rnfE | Electron transport complex protein RnfE | +7.124 | +5.601 | 0.0039 | |
| RCAP_rcc03291_at | rnfG | Electron transport complex protein RnfG | +8.091 | +3.526 | 0.0028 | |
| RCAP_rcc00768_at | hupB | Hydrogenase, large subunit | +1.442 | +3.407 | 0.0749 | |
| Protein Fate | RCAP_rcc00223_at | dnaJ | Chaperone DnaJ | −4.573 | −2.022 | 0.0422 |
| RCAP_rcc00224_at | dnaK | Chaperone DnaK | −5.482 | −1.307 | 0.0031 | |
| RCAP_rcc02977_at | clpA | ATP-dependent Clp protease, ATP-binding subunit ClpA | −3.889 | −1.271 | 0.0850 | |
| RCAP_rcc03406_at | clpB | Chaperone ClpB | −12.622 | −1.735 | 0.0035 | |
| RCAP_rcc02609_at | clpP | ATP-dependent Clp protease, ATP-binding subunit ClpX | −2.995 | −1.054 | 0.0292 | |
| RCAP_rcc01167_at | clpS | ATP-dependent Clp protease adaptor protein ClpS | −3.602 | −1.333 | 0.0108 | |
| RCAP_rcc02478_at | groL | Chaperonin GroL | −8.799 | −1.019 | 0.0037 | |
| RCAP_rcc02477_at | groS | Chaperonin GroS | −6.357 | −1.041 | 0.0021 | |
| RCAP_rcc02818_at | ibpA | Small heat shock protein IbpA | −13.62 | −1.762 | 0.0023 | |
| RCAP_rcc02583_at | lon | ATP-dependent protease La | −5.656 | −1.884 | 0.0193 | |
| RCAP_rcc00034_s_at | hslU | ATP-dependent hsl protease ATP-binding subunit hslU | −5.356 | −3.532 | 0.0979 | |
| RCAP_rcc00035_s_at | hslV | ATP-dependent protease HslV | −5.836 | −2.666 | 0.0585 | |
| RCAP_rcc00480_at | rpsU | 30S ribosomal protein S21 | +5.586 | +2.147 | 0.0835 | |
| RCAP_rcc00327_at | rplQ | 50S ribosomal protein L17 | +4.036 | +2.120 | 0.0982 | |
| Cell Envelope Biogenesis | RCAP_rcc00373_at | mdoH | Glucans biosynthesis glucosyltransferase H | −4.527 | −1.843 | 0.0506 |
| RCAP_rcc01872_at | lpxD | UDP-3-O-[3-hydroxymyristoyl] glucosamine N-acyltransferase | −2.652 | −1.499 | 0.0668 | |
| RCAP_rcc03179_at | phbB | Acetoacetyl-CoA reductase | +2.664 | +1.002 | 0.0081 | |
| Nucleic Acid Metabolism | RCAP_rcc00561_at | mopA | Molybdenum transport operon repressor MopA | +2.673 | +1.150 | 0.0027 |
| RCAP_rcc00568_at | rpoN | RNA polymerase σ-54 factor | +6.031 | +2.602 | 0.0269 | |
| RCAP_rcc01801_at | hfq | RNA chaperone Hfq | +2.452 | −1.020 | 0.0017 | |
| RCAP_rcc02165_at | rne | Ribonuclease E | +3.679 | −1.004 | 0.0014 | |
| RCAP_rcc01384_at | uvrB | UvrABC system protein B | −4.826 | −1.998 | 0.0136 | |
| RCAP_rcc00201_at | - | site specific DNA methyltransferase | +2.578 | +1.070 | 0.0041 | |
| Transporter and Binding Proteins | RCAP_rcc00563_at | modB | Molybdenum ABC transporter, permease protein ModB | +3.113 | +1.062 | 0.0033 |
| Functional Group | Probe Set ID | Gene Symbol | Description | Fold Change | p Value | ||
|---|---|---|---|---|---|---|---|
| 2 h | 6 h | ||||||
| Nitrogen Metabolism | RCAP_rcc01674_at | glnA | Glutamine synthetase | −5.476 | −9.266 | 0.0013 | |
| RCAP_rcc03387_at | glnB | Nitrogen regulatory protein P-II | −2.532 | −7.445 | 0.0912 | ||
| Photosynthesis | RCAP_rcc00733_at | sdhA | Succinate dehydrogenase, flavoprotein subunit | −1.982 | −2.147 | 0.0089 | |
| RCAP_rcc00736_at | sdhB | Succinate dehydrogenase, iron-sulfur subunit | −2.558 | −3.508 | 0.0324 | ||
| RCAP_rcc02531_at | pucA | Light-harvesting protein B-800/850, α chain | +1.070 | −2.053 | 0.0405 | ||
| RCAP_rcc00660_at | pucC | PucC protein | −2.221 | −1.728 | 0.0855 | ||
| RCAP_rcc00693_at | pufL | Photosynthetic reaction center, L subunit | −1.894 | −3.864 | 0.0825 | ||
| RCAP_rcc00694_at | pufM | Photosynthetic reaction center, M subunit | −2.156 | −3.102 | 0.0691 | ||
| RCAP_rcc02970_at | atpC | ATP synthase F1, epsilon subunit | −1.981 | −2.559 | 0.0380 | ||
| RCAP_rcc02971_at | atpD | ATP synthase F1, β subunit | −6.198 | −9.121 | 0.0211 | ||
| RCAP_rcc02150_at | acpA | Aconitate hydratase | −2.482 | −2.801 | 0.0930 | ||
| Electron Transport | RCAP_rcc00573_at | fdxD | Ferredoxin V | −3.345 | −2.949 | 0.0075 | |
| RCAP_rcc00767_at | hupA | Hydrogenase, small subunit | −2.436 | −3.333 | 0.0778 | ||
| RCAP_rcc00768_at | hupB | Hydrogenase, large subunit | −3.744 | −5.254 | 0.0249 | ||
| RCAP_rcc00769_at | hupC | Hydrogenase, cytochrome b subunit | −2.401 | −2.354 | 0.0387 | ||
| RCAP_rcc01517_at | nuoA | NADH-quinone oxidoreductase, A subunit | −2.363 | −2.214 | 0.0795 | ||
| RCAP_rcc01518_at | nuoB | NADH-quinone oxidoreductase, B subunit | −2.401 | −2.791 | 0.0719 | ||
| RCAP_rcc01519_at | nuoC | NADH-quinone oxidoreductase, C subunit | −2.884 | −2.758 | 0.0153 | ||
| RCAP_rcc01520_at | nuoD | NADH-quinone oxidoreductase, D subunit | −3,101 | −4.132 | 0.0043 | ||
| RCAP_rcc01521_at | nuoE | NADH-quinone oxidoreductase, E subunit | −2,700 | −2.869 | 0.0289 | ||
| RCAP_rcc01527_at | nuoG | NADH-quinone oxidoreductase, G subunit | −1,720 | −2.262 | 0.0553 | ||
| RCAP_rcc01529_at | nuoH | NADH-quinone oxidoreductase, H subunit | −2.118 | −1,663 | 0.0749 | ||
| Protein Fate | RCAP_rcc03406_at | clpB | Chaperone ClpB | −2.502 | −2.580 | 0.0120 | |
| RCAP_rcc02609_at | clpP | ATP-dependent Clp protease, ATP-binding subunit ClpX | −2,358 | −3.859 | 0.0676 | ||
| RCAP_rcc01167_at | clpS | ATP-dependent Clp protease adaptor protein ClpS | +1291 | +2.704 | 0.0839 | ||
| RCAP_rcc02583_at | Lon | ATP-dependent protease La | −4.331 | −2.544 | 0.0235 | ||
| RCAP_rcc00480_at | rpsU | 30S ribosomal protein S21 | +12.697 | +4.962 | 0.0026 | ||
| RCAP_rcc00321_at | rplO | 50S ribosomal protein L15 | +8.431 | +8.222 | 5.4674 × 10−4 | ||
| RCAP_rcc00361_at | rpmE | 50S ribosomal protein L31 | +3.899 | +2.197 | 0.0983 | ||
| Cell Envelope Biogenesis | RCAP_rcc00350_at | rpmH | 50S ribosomal protein L34 | +4.232 | +3.551 | 0.0134 | |
| RCAP_rcc02380_at | ftsI | Peptidoglycan synthetase FtsI | −2.279 | −2.546 | 0.0851 | ||
| RCAP_rcc01678_at | acpP | Acyl carrier protein | +4.132 | +1.487 | 0.0863 | ||
| RCAP_rcc00559_at | pmtA | Phosphatidylethanolamine N-methyltransferase | +3.117 | +2.273 | 0.0809 | ||
| RCAP_rcc00028_at | Idi | Isopentenyl-diphosphate delta-isomerase | +4.085 | +2.981 | 0.0973 | ||
| Nucleic Acid Metabolism | RCAP_rcc00349_at | rnpA | Ribonuclease P | +2.065 | +1.919 | 0.0393 | |
| RCAP_rcc00286_at | nusG | Transcription antitermination protein NusG | +3.113 | +2.461 | 0.0678 | ||
| RCAP_rcc03054_at | rpoD | RNA polymerase σ factor RpoD | −2.919 | −2.406 | 0.0329 | ||
| RCAP_rcc00458_at | rpoH | RNA polymerase σ-32 factor | −3.361 | −1.667 | 0.0032 | ||
| RCAP_rcc01751_at | recA | RecA protein | +1.895 | +2.357 | 0.0845 | ||
| RCAP_rcc00201_at | - | Site specific DNA methyltransferase | +2.231 | −1.070 | 0.0013 | ||
| Transporter and Binding Proteins | RCAP_rcc01243_at | potA | Polyamine ABC transporter, ATP binding protein PotA | −8.289 | −10.273 | 0.0073 | |
| RCAP_rcc01245_at | potB | Polyamine ABC transporter, permease protein PotB | −4.566 | −7.072 | 0.0040 | ||
| RCAP_rcc01244_at | potD | Polyamine ABC transporter, periplasmic polyamine-binding protein PotD | −14.108 | −15.842 | 0.0014 | ||
| RCAP_rcc02186_at | potF | Spermidine/putrescine ABC transporter, periplasmic spermidine/putrescine-binding protein PotF | −4.010 | −4.508 | 0.0193 | ||
| RCAP_rcc02183_at | potG | Spermidine/putrescine ABC transporter, ATP-binding protein PotG | −3.135 | −2.833 | 0.0203 | ||
| RCAP_rcc01895_at | potH | Spermidine/putrescine ABC transporter, permease protein | −2.976 | −2.301 | 0.0588 | ||
| RCAP_rcc01246_at | potI | Polyamine ABC transporter, permease protein PotI | −3.940 | −4.112 | 0.0304 | ||
| RCAP_rcc01879_at | sufC | FeS assembly ATPase SufC | −6.598 | −6.351 | 2.1582 × 10−4 | ||
| RCAP_rcc00092_at | feoB | Ferrous iron transport protein B | −2.362 | −1.524 | 0.0338 | ||
| RCAP_rcc01878_at | sufD | FeS assembly protein SufD | −4.363 | −4.224 | 0.0306 | ||
| RCAP_rcc01376_at | dctP | TRAP dicarboxylate transporter, DctP subunit | −1.538 | −2.436 | 0.0186 | ||
| RCAP_rcc00706_at | oppA | Oligopeptide ABC transporter, Periplasmic oligopeptide-binding protein OppA | −3.203 | −3.085 | 0.0069 | ||
2.3. Effect of Temperature Stress on Hydrogen Production Metabolism of R. capsulatus
2.4. Effect of Temperature Stress on Protein Metabolism of R. capsulatus
2.5. Effect of Temperature Stress on Lipid, Cell Envelope, and Trasporter Metabolisms of R. capsulatus
2.6. Effect of Temperature Stress on Nucleic Acid Metabolism of R. capsulatus
3. Experimental Section
3.1. Growth of Bacteria and Stress Treatments
3.2. Microarray Analysis
3.3. Real Time PCR Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koku, H.; Eroglu, I.; Gündüz, U.; Yücel, M.; Türker, M. Aspects of the metabolism of hydrogen production by Rhodobacter sphaeroides. Int. J. Hydrog. Energy 2002, 27, 1315–1329. [Google Scholar] [CrossRef]
- Hallenbeck, P.; Benemann, J. Biological hydrogen production; fundamentals and limiting processes. Int. J. Hydrog. Energy 2002, 27, 1185–1193. [Google Scholar] [CrossRef]
- Weaver, P.F.; Wall, J.D.; Gest, H. Characterization of Rhodopseudomonas capsulata. Arch. Microbiol. 1975, 105, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Basak, N.; Das, D. The prospect of purple non-sulfur (PNS) photosynthetic bacteria for hydrogen production: The present state of the art. World J. Microbiol. Biotechnol. 2007, 23, 31–42. [Google Scholar] [CrossRef]
- Androga, D.D.; Sevinc, P.; Koku, H.; Yücel, M.; Gündüz, U.; Eroglu, I. Optimization of temperature and light intensity for improved photofermentative hydrogen production using Rhodobacter capsulatus DSM1710. Int. J. Hydrog. Energy 2014, 39, 2472–2480. [Google Scholar]
- Ünlü, G.; Sevinç, P.; Gündüz, U. Expression analysis of nif A and nif H genes in Rhodobacter sphaeroides O.U.001 at different temperatures. In International Symposium on Biotechnology: Developments and Trends, BTEC 2009; Middle East Technical University: Ankara, Turkey, 2009; p. 91. [Google Scholar]
- Özgür, E.; Uyar, B.; Öztürk, Y.; Yücel, M.; Gündüz, U.; Eroglu, I. Biohydrogen production by Rhodobacter capsulatus on acetate at fluctuating temperatures. Resour. Conserv. Recycl. 2010, 54, 310–314. [Google Scholar] [CrossRef]
- Androga, D.D.; Özgür, E.; Gündüz, U.; Yücel, M.; Eroglu, I. Factors affecting the longterm stability of biomass and hydrogen productivity in outdoor photofermentation. Int. J. Hydrog. Energy 2011, 36, 11369–11378. [Google Scholar] [CrossRef]
- Avcioglu, S.G.; Özgür, E.; Eroglu, I.; Yücel, M.; Gündüz, U. Biohydrogen production in an outdoor panel photobioreactor on dark fermentation effluent of molasses. Int. J. Hydrog. Energy 2011, 36, 11360–11368. [Google Scholar] [CrossRef]
- Ray, B. Impact of bacterial injury and repair in food microbiology: Its past, present and future. J. Food Prot. 1986, 49, 651–655. [Google Scholar]
- Shi, L.; Perkins, R.G.; Tong, W. The current status of DNA microarrays. In Microarrays: Preparation, Microfluidics, Detection Methods, and Biological Applications, 1st ed.; Dill, K., Liu, R., Grodzinsky, P., Eds.; Springer: New York, NY, USA, 2009; Volume 1, pp. 3–24. [Google Scholar]
- Yilmaz, R.; Yücel, M.; Öktem, H.A. Quality assessment of gene expression data for an Affymetrix platform with the two sample t-tests statistical analysis. Int. J. Biotechnol. Biochem. 2008, 4, 101–108. [Google Scholar]
- Shi, L.; Jones, W.D.; Jensen, V.R.; Harris, S.C.; Perkins, R.G.; Goodsaid, F.M.; Guo, L.; Croner, L.J.; Boysen, C.; Fang, H.; et al. The balance of reproducibility, sensitivity, and specificity of lists of differentially expressed genes in microarray studies. BMC Bioinform. 2008, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Onder, O.; Aygun-Sunar, S.; Selamoglu, N.; Daldal, F. A glimpse into the proteome of phototrophic bacterium Rhodobacter capsulatus. Adv. Exp. Med. Biol. 2010, 675, 179–209. [Google Scholar] [PubMed]
- Hillmer, P.; Gest, H. H2 Metabolism in the photosynthetic bacterium Rhodopseudomonas capsulata: H2 production by growing cultures. J. Bacteriol. 1977, 129, 724–731. [Google Scholar] [PubMed]
- McEwan, A.G. Photosynthetic electron transport and anaerobic metabolism in purple non-sulfur bacteria. Antonie van Leeuwenhoek 1994, 66, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.J.; Jackson, J.B.; McEwan, A.G. Anaerobic respiration in the Rhodospirillaceae: characterisation of pathways and evaluation of the roles in the redox balancing during photosynthesis. FEMS Microbiol. Rev. 1987, 46, 117–143. [Google Scholar] [CrossRef]
- Biegel, E.; Schmidt, S.; Gonzáles, J.M.; Müller, V. Biochemistry, evolution and physiological function of the Rnf complex, a novel ion-motive electron transport complex in prokaryotes. Cell. Mol. Life Sci. 2011, 68, 613–634. [Google Scholar] [CrossRef] [PubMed]
- Cullen, P.J.; Foster-Hartnett, D.; Gabbert, K.K.; Kranz, R.G. Structure and expression of the alternative sigmafactor, RpoN, in Rhodobacter capsulatus; physiological relevance of an autoactivated nifU2-rpoN superoperon. Mol. Biol. 1994, 11, 51–65. [Google Scholar]
- Leif, H.; Sled, V.D.; Ohnishi, T.; Weiss, H.; Friedrich, T. Isolation and characterization of the proton-translocating NADH: Ubiquinone oxidoreductase from Escherichia coli. Eur. J. Biochem. 1995, 230, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Macario, A.J.L.; Macario, E.J. Heat shock response, overview. In The Encyclopedia of Stress, 2nd ed.; Fink, G., Ed.; Academic Press: California, CA, USA, 2000; pp. 350–357. [Google Scholar]
- Gomes, S.L.; Simão, R.C.G. Stress responses: Heat. Encycl. Microbiol. 2009, 1, 464–476. [Google Scholar]
- Yura, T.; Nagai, H.; Mori, H. Regulation of the heat shock response in bacteria. Annu. Rev. Microbiol. 1994, 47, 321–250. [Google Scholar] [CrossRef] [PubMed]
- Beckering, C.L.; Steil, L.; Weber, M.H.W.; Völker, U.; Maraheil, M.A. Genomewide transcriptional analysis of the cold shock response in Bacillus subtilis. J. Bacteriol. 2002, 184, 6395–6402. [Google Scholar] [CrossRef] [PubMed]
- Methe, B.A.; Nelson, K.E.; Deming, J.W.; Momen, B.; Melamud, E.; Zhang, X.; Moult, J.; Madupu, R.; Nelson, W.C.; Dodson, R.J.; et al. The psyrophilic lifestyle as revealed by the genome sequence of Colwellia psycherythraea 34H through genomic and proteomic analyses. Proc. Natl. Acad. Sci. USA 2005, 102, 10913–10918. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Kim, M.S.; Lee, J.K. Phosphatidycholine is required for the efficient formation of photosynthetic membrane and B800-850 light harvesting complex in Rhodobacter sphaeroides. J. Microbiol. Bacteriol. 2007, 17, 373–377. [Google Scholar]
- Hahn, F.M.; Baker, J.A.; Poulter, C.D. Open reading frame 176 in the photosynthesis gene cluster of Rhodobacter capsulatus encods idi, a gene for isopenthyl diphosphate isomerase. J. Bacteriol. 1996, 178, 619–624. [Google Scholar] [PubMed]
- Majerus, P.W.; Alberts, A.W.; Vagelos, P.R. Acyl carrier protein. III An enoyl hydrase specific for acyl carrier protein thioesters. J. Biol. Chem. 1964, 240, 618–621. [Google Scholar]
- Raina, A.; Jänne, J. Physiology of the natural polyamines putrescine, spermidine and spermine. Med. Biol. 1975, 53, 121–147. [Google Scholar] [PubMed]
- Shen, G.; Golbeck, J.H. Assembly of the bound iron–sulfur clusters in Photosystem I. Adv. Photosynth. Respir. 2006, 24, 529–548. [Google Scholar]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Hess, W.R.; Berghoff, B.A.; Wilde, A.; Steglich, C.; Klug, G. Riboregulators and the role of Hfq in photosynthetic bacteria. RNA Biol. 2014, 11, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Biebl, H.; Pfennig, N. Isolation of member of the family Rhodosprillaceae. In The Prokaryotes; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: New York, NY, USA, 1981; pp. 267–273. [Google Scholar]
- Uyar, B.; Eroglu, I.; Yücel, M.; Gündüz, U.; Türker, L. Effect of light intensity, wavelength and illumination protocol on hydrogen production in photobioreactors. Int. J. Hydrog. Energy 2007, 32, 4670–4677. [Google Scholar] [CrossRef]
- Strnad, H.; Lapidus, A.; Paces, J.; Ulbrich, P.; Vlcek, C.; Paces, V.; Haselkorn, R. Complete genome sequence of the photosynthetic purple nonsulfur bacterium Rhodobacter capsulatus SB 1003. J. Bacteriol. 2010, 192, 3545–3546. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gürgan, M.; Erkal, N.A.; Özgür, E.; Gündüz, U.; Eroglu, I.; Yücel, M. Transcriptional Profiling of Hydrogen Production Metabolism of Rhodobacter capsulatus under Temperature Stress by Microarray Analysis. Int. J. Mol. Sci. 2015, 16, 13781-13797. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160613781
Gürgan M, Erkal NA, Özgür E, Gündüz U, Eroglu I, Yücel M. Transcriptional Profiling of Hydrogen Production Metabolism of Rhodobacter capsulatus under Temperature Stress by Microarray Analysis. International Journal of Molecular Sciences. 2015; 16(6):13781-13797. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160613781
Chicago/Turabian StyleGürgan, Muazzez, Nilüfer Afşar Erkal, Ebru Özgür, Ufuk Gündüz, Inci Eroglu, and Meral Yücel. 2015. "Transcriptional Profiling of Hydrogen Production Metabolism of Rhodobacter capsulatus under Temperature Stress by Microarray Analysis" International Journal of Molecular Sciences 16, no. 6: 13781-13797. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160613781