
Evidence for P-Glycoprotein Involvement in Cell Volume Regulation Using Coulter Sizing in Flow Cytometry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
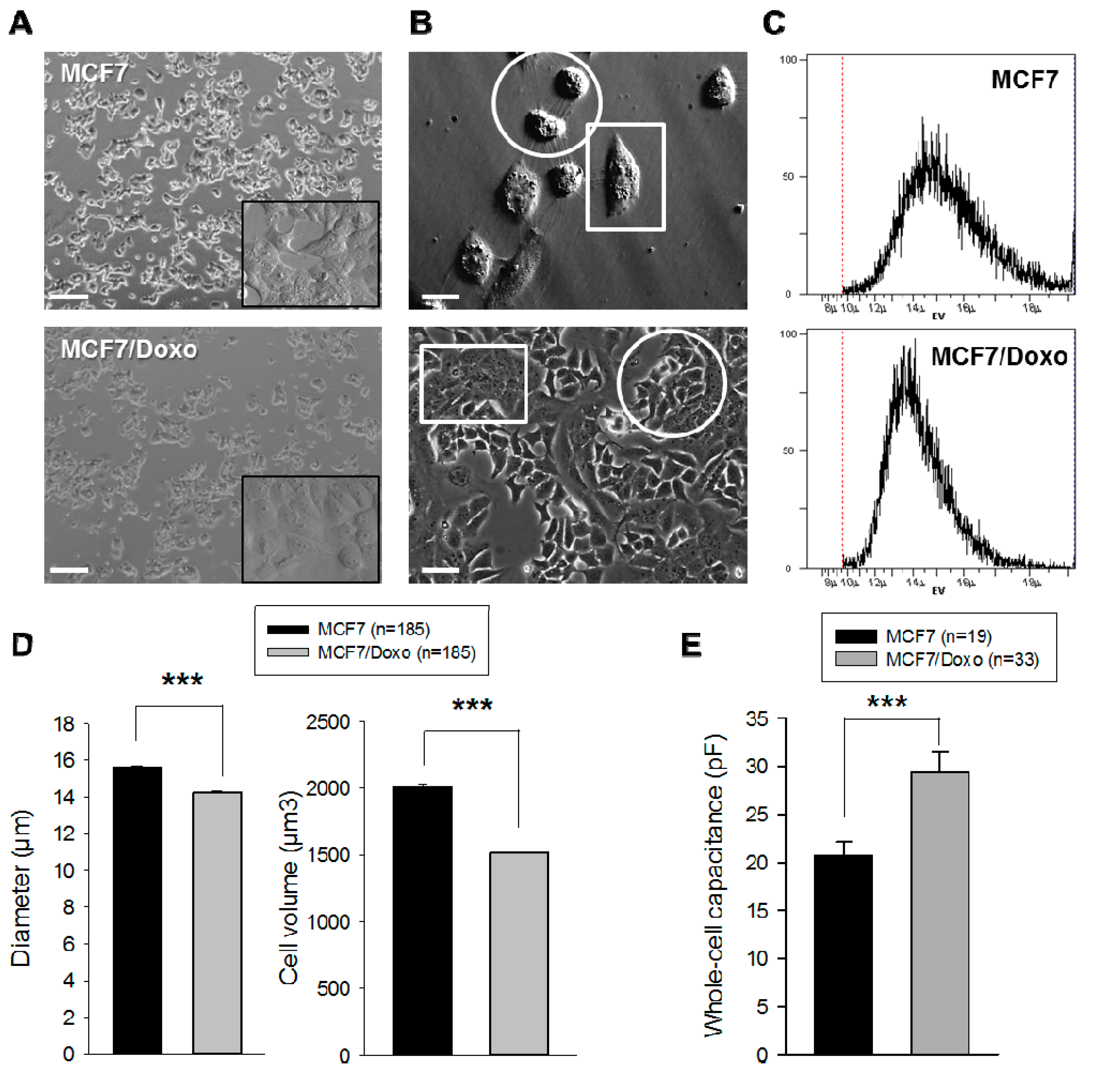
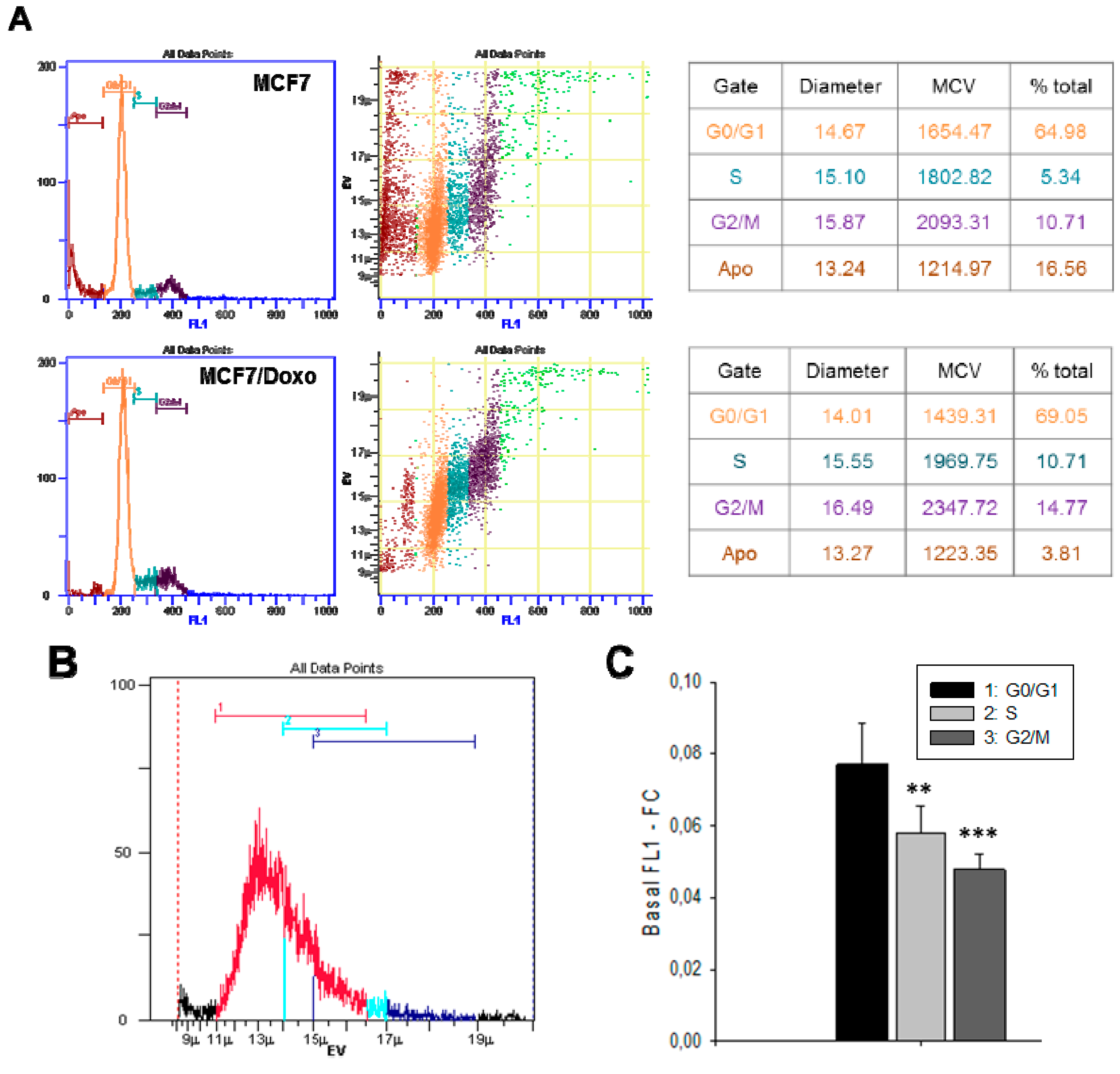
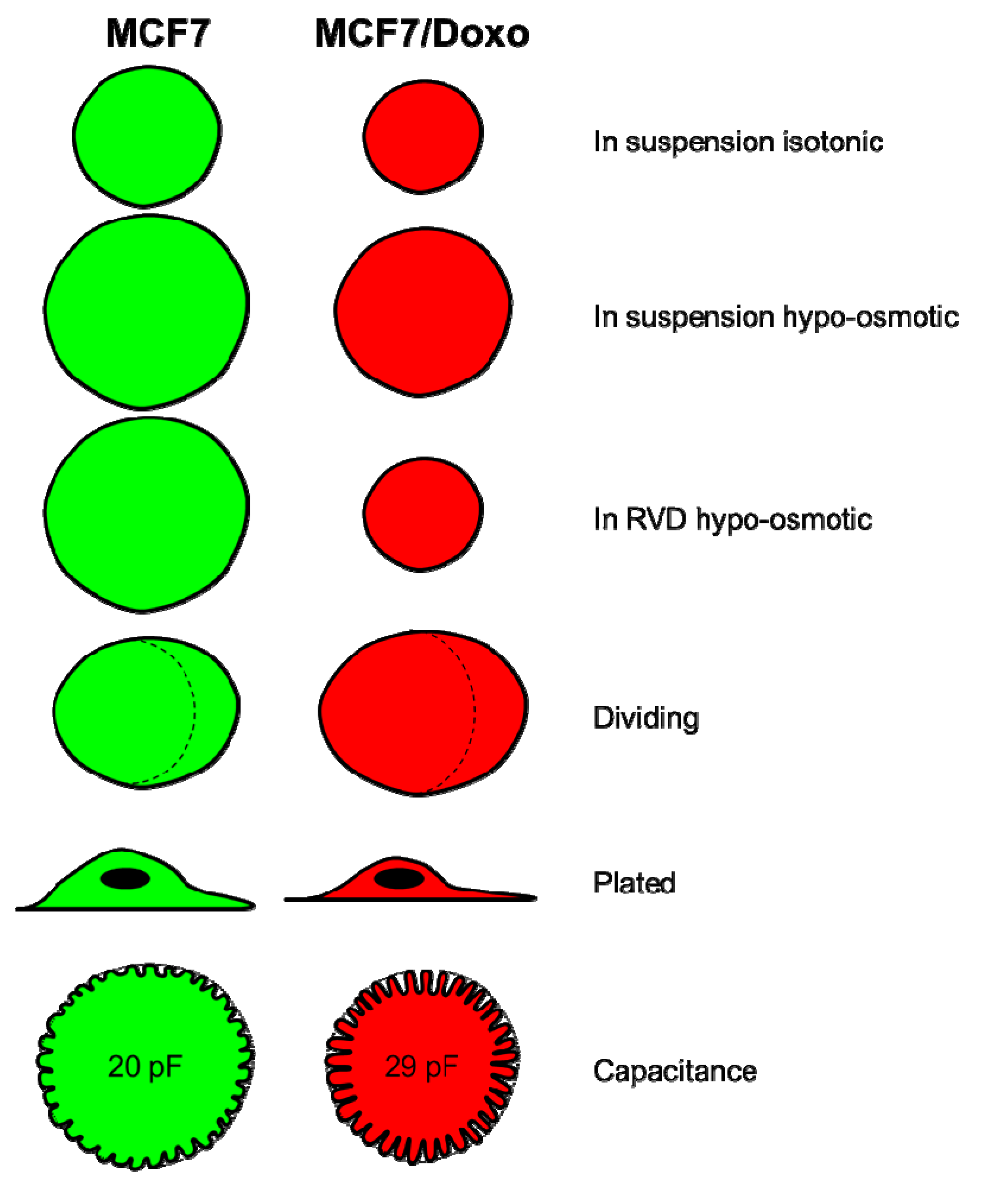
2.1. Resistant and Sensitive Variants Display Different Cell Volumes, Shapes and Membrane Capacitance


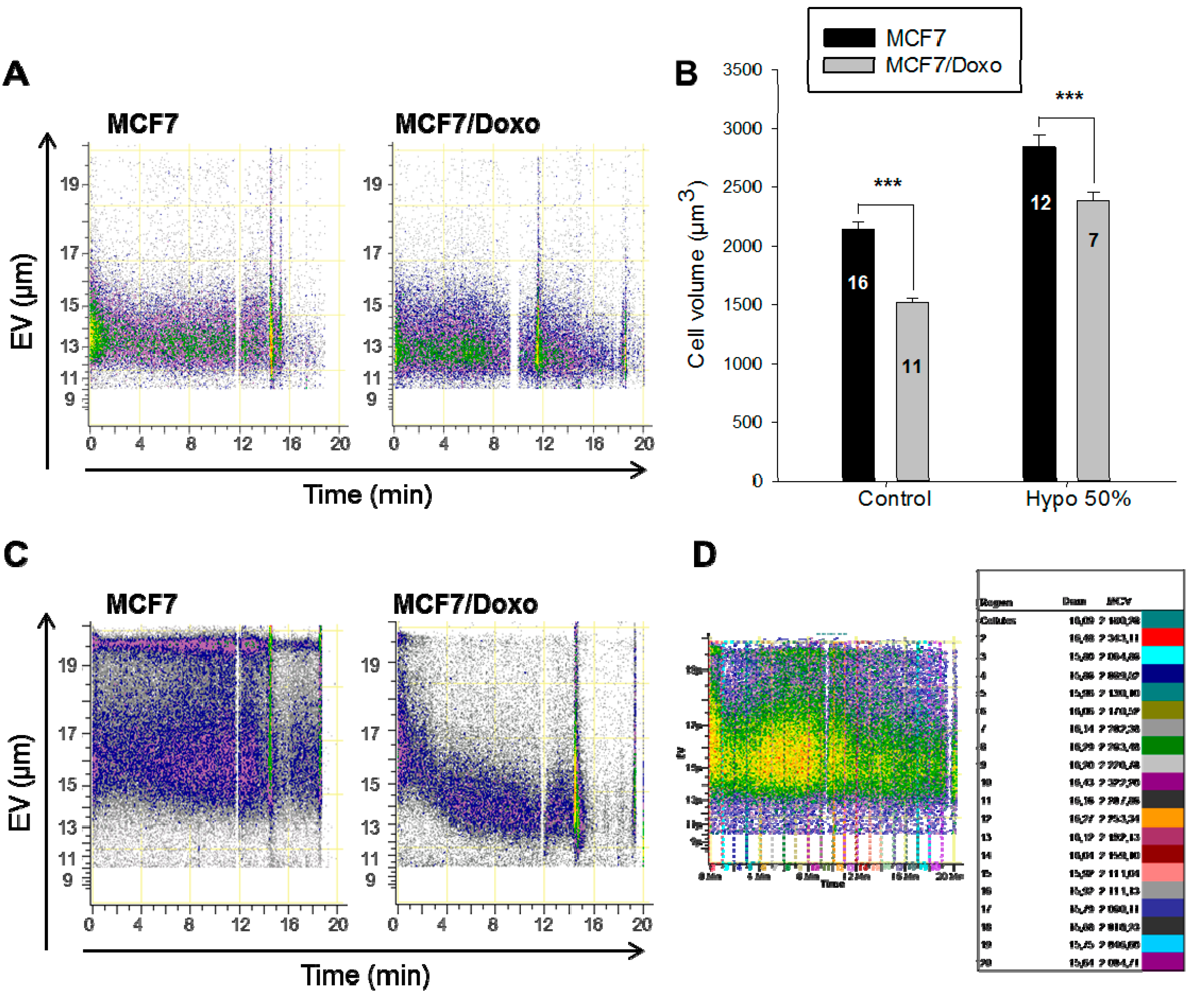
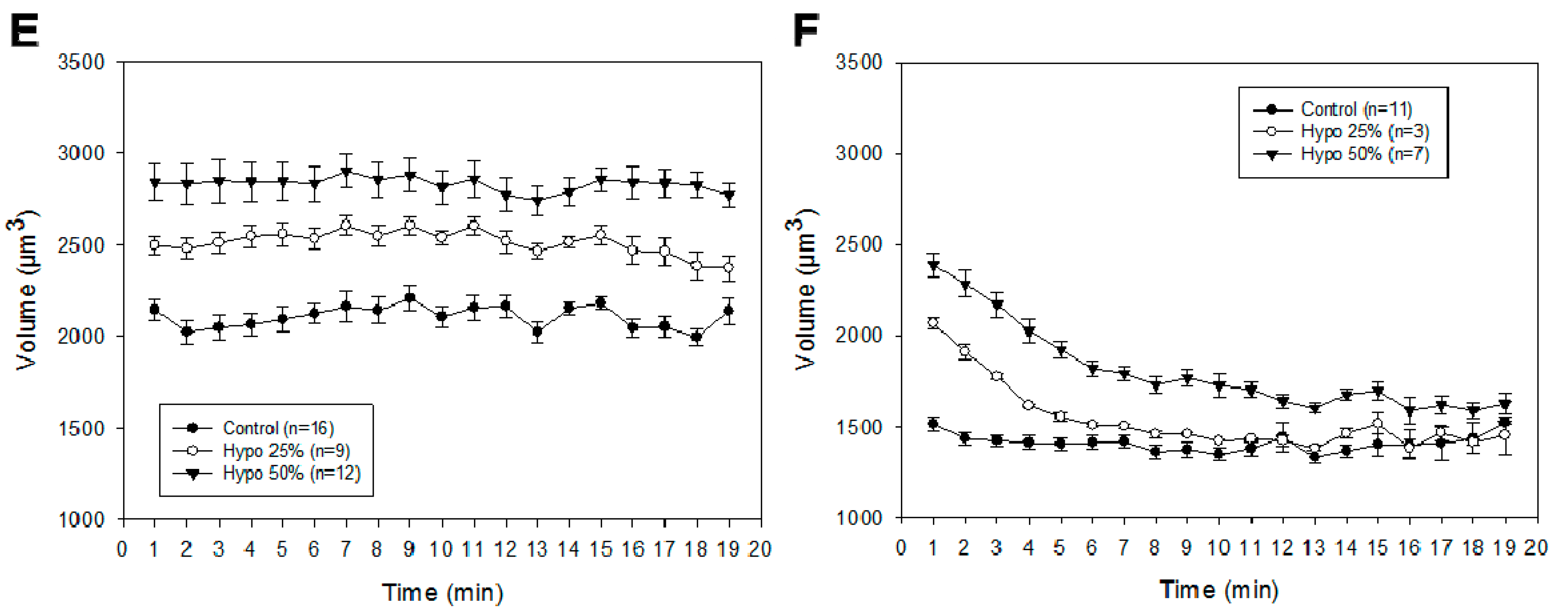
2.2. Cell Volume Monitoring during Hypo-Osmotic Shocks


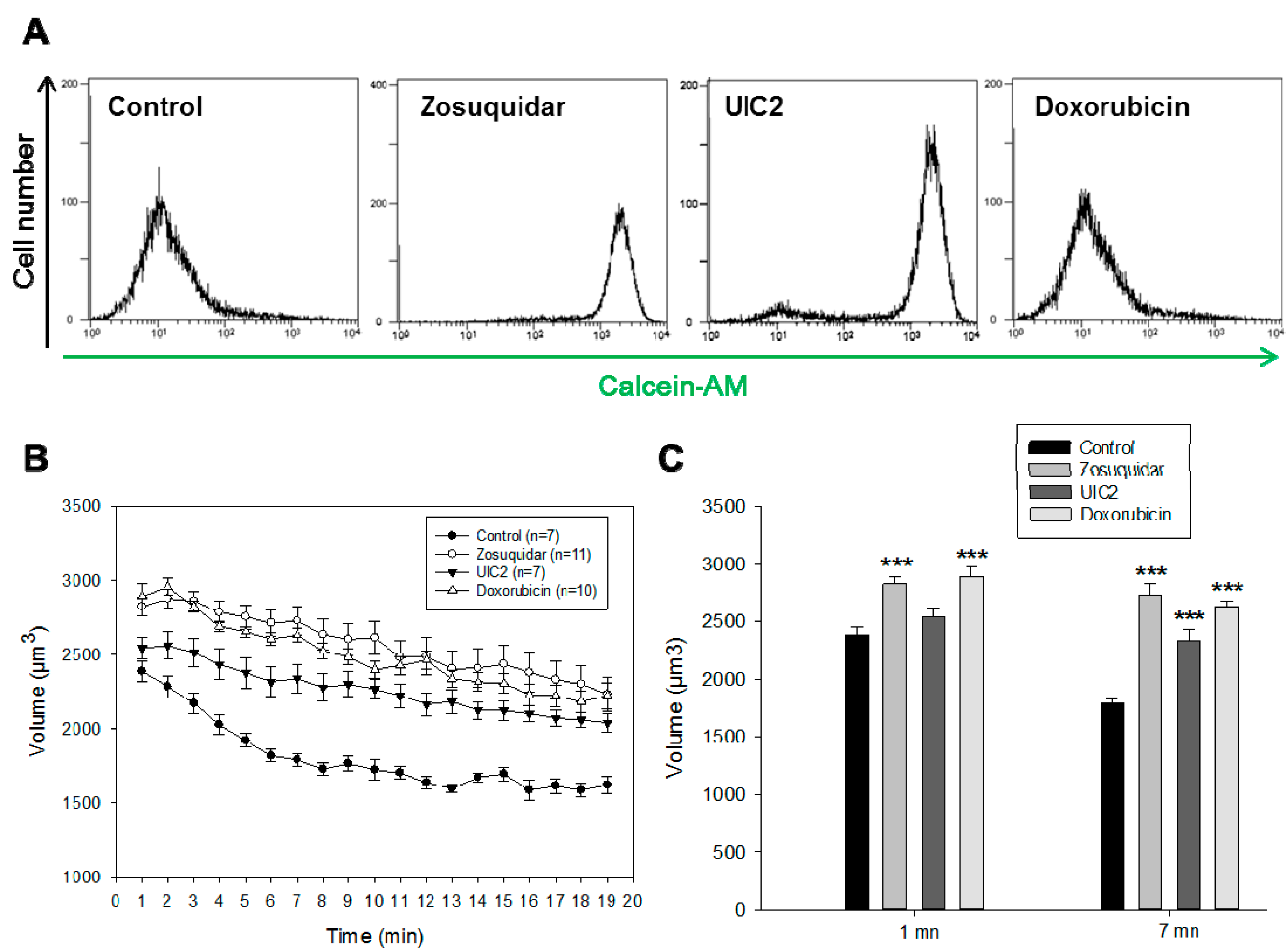
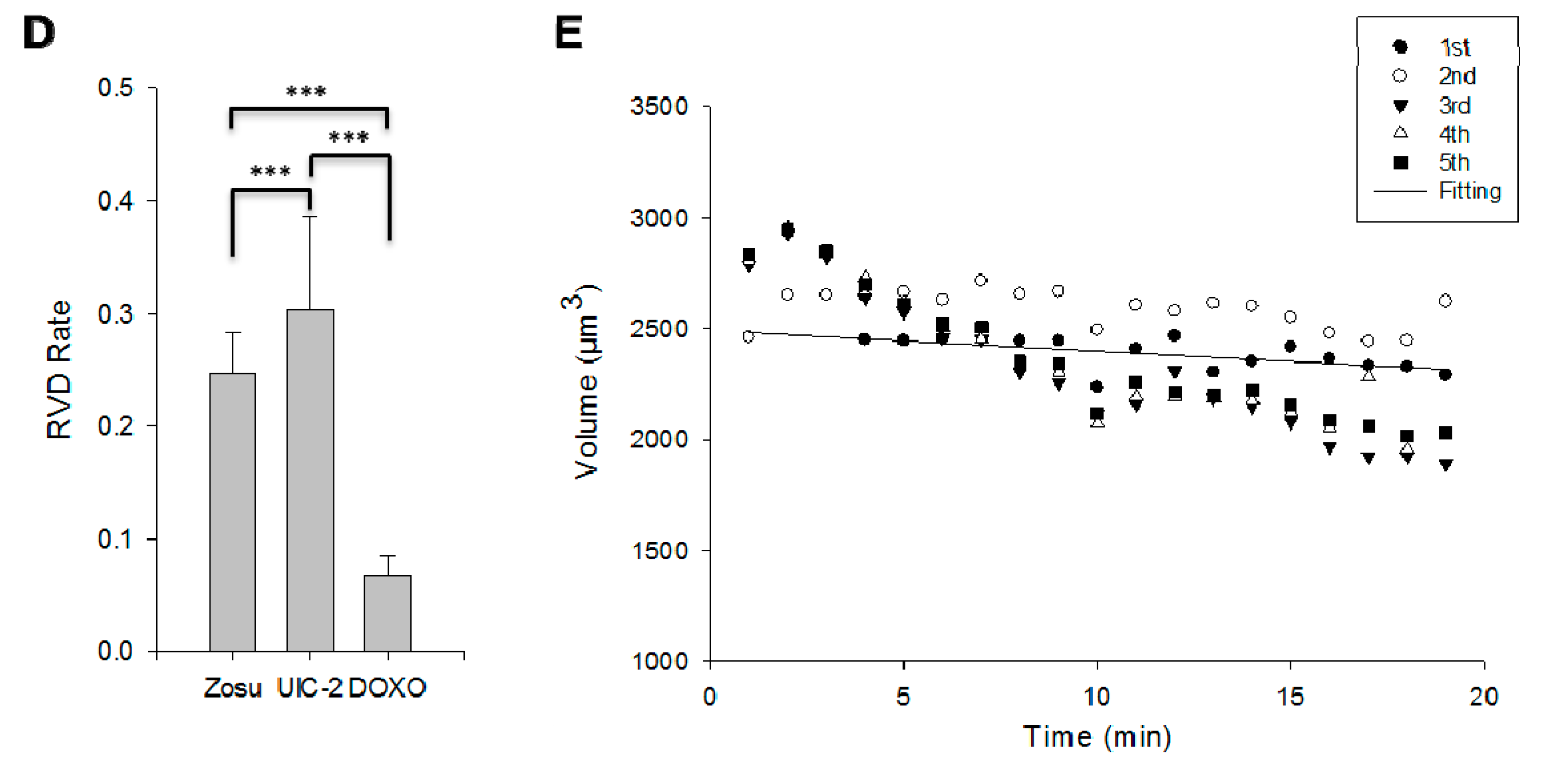
2.3. RVD in MCF7/Doxo Cells Is Dependent on P-gp Activity


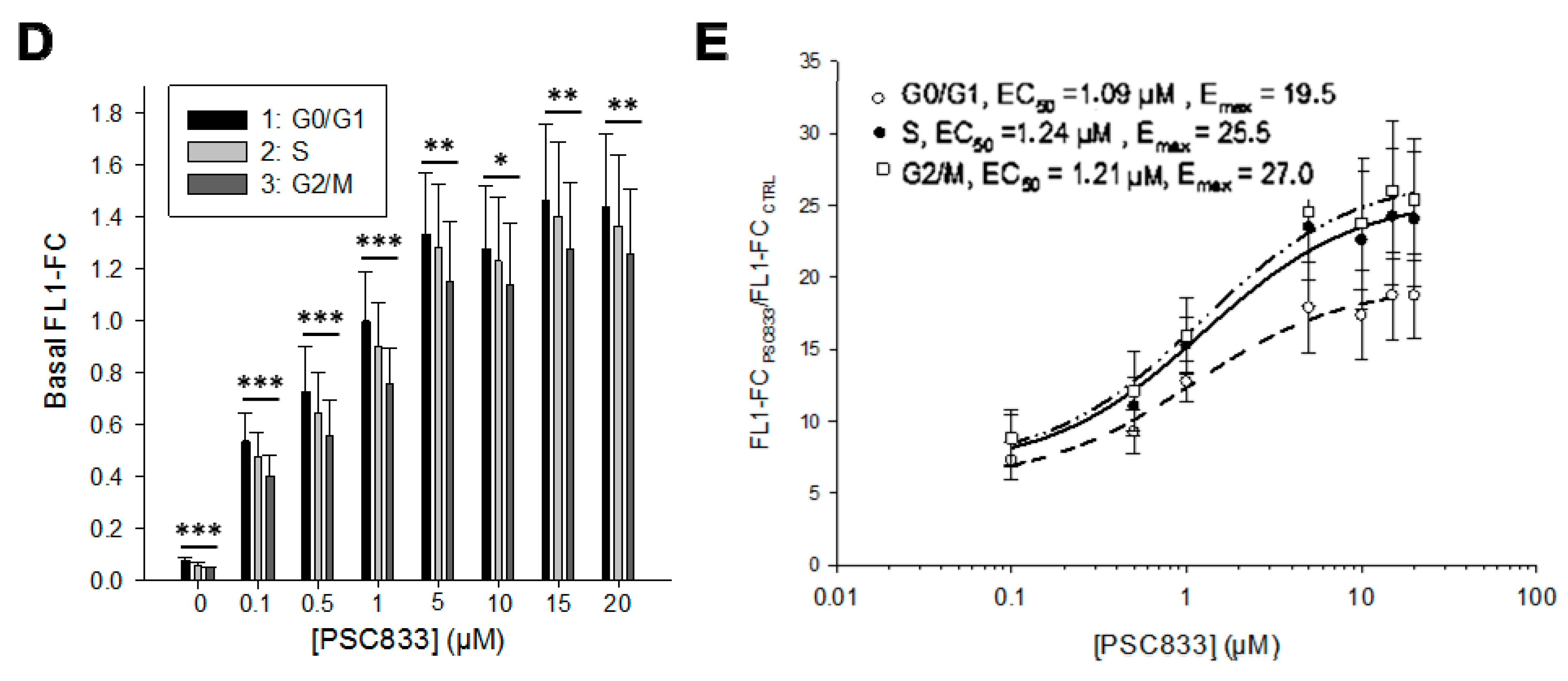
2.4. P-gp Activity and Cell Cycle


3. Discussion

4. Material and Methods
4.1. Cell Cultures
4.2. Reagents
4.3. Flow Cytometry
4.4. Osmostic Challenges and RVD Rate Calculation
4.5. P-gp Activity
4.6. Cell Cycle
4.7. Electrophysiology
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| AVD | apoptotic volume decrease |
| EV | electronic volume |
| MDR | MultiDrug Resistance |
| P-gp | P-glycoprotein |
| RVD | regulatory volume decrease |
| RVI | regulatory volume increase |
References
- Okada, Y. Volume expansion-sensing outward-rectifier Cl− channel: Fresh start to the molecular identity and volume sensor. Am. J. Physiol. 1997, 273, C755–C789. [Google Scholar] [PubMed]
- Okada, Y.; Maeno, E. Apoptosis, cell volume regulation and volume-regulatory chloride channels. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 377–783. [Google Scholar] [CrossRef]
- Agre, P.; King, L.S.; Yasui, M.; Guggino, W.B.; Ottersen, O.P.; Fujiyoshi, Y.; Engel, A.; Nielsen, S. Aquaporin water channels—From atomic structure to clinical medicine. J. Physiol. 2002, 542 Pt 1, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Sardini, A.; Amey, J.S.; Weylandt, K.H.; Nobles, M.; Valverde, M.A.; Higgins, C.F. Cell volume regulation and swelling-activated chloride channels. Biochim. Biophys. Acta 2003, 1618, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Busch, G.L.; Volkl, H. The diversity of volume regulatory mechanisms. Cell. Physiol. Biochem. 1998, 8, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Peak, M.; al-Habori, M.; Agius, L. Regulation of glycogen synthesis and glycolysis by insulin, pH and cell volume. Interactions between swelling and alkalinization in mediating the effects of insulin. Biochem. J. 1992, 282, 797–805. [Google Scholar] [PubMed]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death: Recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ. 2009, 16, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Maeno, E.; Shimizu, T.; Dezaki, K.; Wang, J.; Morishima, S. Receptor-mediated control of regulatory volume decrease (RVD) and apoptotic volume decrease (AVD). J. Physiol. 2001, 532, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Bosch, I.; Croop, J. P-glycoprotein multidrug resistance and cancer. Biochim. Biophys. Acta 1996, 1288, F37–F54. [Google Scholar] [CrossRef]
- Endicott, J.A.; Ling, V. The biochemistry of P-glycoprotein-mediated multidrug resistance. Annu. Rev. Biochem. 1989, 58, 137–171. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, T.J.; Gunther, W. Chloride channels: An emerging molecular picture. Bioessays 1997, 19, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Eggermont, J.; Voets, T.; Droogmans, G. Volume-activated Cl− channels. Gen. Pharmacol. 1996, 27, 1131–1140. [Google Scholar] [CrossRef]
- Valverde, M.A.; Diaz, M.; Sepulveda, F.V.; Gill, D.R.; Hyde, S.C.; Higgins, C.F. Volume-regulated chloride channels associated with the human multidrug-resistance P-glycoprotein. Nature 1992, 355, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.K.; Vanoye, C.G.; Reuss, L. Swelling-activated chloride currents in a drug-sensitive cell line and a P-glycoprotein-expressing derivative are underlied by channels with the same pharmacological properties. Cell. Physiol. Biochem. 1998, 8, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Hardy, S.P.; Goodfellow, H.R.; Valverde, M.A.; Gill, D.R.; Sepulveda, V.; Higgins, C.F. Protein kinase C-mediated phosphorylation of the human multidrug resistance P-glycoprotein regulates cell volume-activated chloride channels. EMBO J. 1995, 14, 68–75. [Google Scholar] [PubMed]
- Bond, T.D.; Valverde, M.A.; Higgins, C.F. Protein kinase C phosphorylation disengages human and mouse-1a P-glycoproteins from influencing the rate of activation of swelling-activated chloride currents. J. Physiol. 1998, 508 Pt 2, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Ehring, G.R.; Osipchuk, Y.V.; Cahalan, M.D. Swelling-activated chloride channels in multidrug-sensitive and -resistant cells. J. Gen. Physiol. 1994, 104, 1129–1161. [Google Scholar] [CrossRef] [PubMed]
- Miwa, A.; Ueda, K.; Okada, Y. Protein kinase C-independent correlation between P-glycoprotein expression and volume sensitivity of Cl− channel. J. Membr. Biol. 1997, 157, 63–69. [Google Scholar] [CrossRef] [PubMed]
- De Greef, C.; Sehrer, J.; Viana, F.; van Acker, K.; Eggermont, J.; Mertens, L.; Raeymaekers, L.; Droogmans, G.; Nilius, B. Volume-activated chloride currents are not correlated with P-glycoprotein expression. Biochem. J. 1995, 307, 713–718. [Google Scholar] [PubMed]
- De Greef, C.; van der Heyden, S.; Viana, F.; Eggermont, J.; de Bruijn, E.A.; Raeymaekers, L.; Droogmans, G.; Nilius, B. Lack of correlation between MDR-1 expression and volume-activation of cloride-currents in rat colon cancer cells. Pflugers Arch. 1995, 430, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.; Poret, A.; Maillet, G.; Leboulenger, F.; le Foll, F. Regulation of volume-sensitive Cl− channels in multi-drug resistant MCF7 cells. Biochem. Biophys. Res. Commun. 2005, 334, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, K.A.; Andersen, E.C.; Hansen, C.F.; Klausen, T.K.; Hougaard, C.; Lambert, I.H.; Hoffmann, E.K. Deregulation of apoptotic volume decrease and ionic movements in multidrug-resistant tumor cells: Role of chloride channels. Am. J. Physiol. Cell Physiol. 2010, 298, C14–C25. [Google Scholar] [CrossRef] [PubMed]
- Torre, A.; Trischitta, F.; Faggio, C. Purinergic receptors and regulatory volume decrease in seabream (Sparus aurata) hepatocytes: A videometric study. Fish Physiol. Biochem. 2012, 38, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, J.; Galas, L.; Boulange-Lecomte, C.; Rioult, D.; Bultelle, F.; Magal, P.; Webb, G.; le Foll, F. Different modalities of intercellular membrane exchanges mediate cell-to-cell p-glycoprotein transfers in MCF-7 breast cancer cells. J. Biol. Chem. 2012, 287, 7374–7387. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, J.; Magal, P.; Boulange-Lecomte, C.; Webb, G.; Le Foll, F. Consequences of cell-to-cell P-glycoprotein transfer on acquired multidrug resistance in breast cancer: A cell population dynamics model. Biol. Direct. 2011, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, J.; Rioult, D.; Abu-Kaoud, N.; Marie, S.; Rafii, A.; Guerrouahen, B.S.; le Foll, F. P-glycoprotein-activity measurements in multidrug resistant cell lines: Single-cell vs. single-well population fluorescence methods. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer associates Inc.: Sunderland, MA, USA, 2001; p. 814. [Google Scholar]
- Gill, D.R.; Hyde, S.C.; Higgins, C.F.; Valverde, M.A.; Mintenig, G.M.; Sepulveda, F.V. Separation of drug transport and chloride channel functions of the human multidrug resistance P-glycoprotein. Cell 1992, 71, 23–32. [Google Scholar] [CrossRef]
- Sardini, A.; Mintenig, G.M.; Valverde, M.A.; Sepulveda, F.V.; Gill, D.R.; Hyde, S.C.; Higgins, C.F.; McNaughton, P.A. Drug efflux mediated by the human multidrug resistance P-glycoprotein is inhibited by cell swelling. J. Cell Sci. 1994, 107 Pt 12, 3281–3290. [Google Scholar] [PubMed]
- Weaver, J.L.; Aszalos, A.; McKinney, L. MDR1/P-glycoprotein function. II. Effect of hypotonicity and inhibitors on Cl− efflux and volume regulation. Am. J. Physiol. 1996, 270, C1453–C1460. [Google Scholar] [PubMed]
- Aleu, J.; Ivorra, I.; Lejarreta, M.; Gonzalez-Ros, J.M.; Morales, A.; Ferragut, J.A. Functional incorporation of P-glycoprotein into Xenopus oocyte plasma membrane fails to elicit a swelling-evoked conductance. Biochem. Biophys. Res. Commun. 1997, 237, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.X.; Wang, L.W.; Jacob, T. The role of MDR1 gene in volume-activated chloride currents in pigmented ciliary epithelial cells. Sheng Li Xue Bao 2002, 54, 1–6. [Google Scholar] [PubMed]
- Shi, X.; Wu, S.; Yang, Y.; Tang, L.; Wang, Y.; Dong, J.; Lu, B.; Jiang, G.; Zhao, W. AQP5 silencing suppresses p38 MAPK signaling and improves drug resistance in colon cancer cells. Tumour Biol. 2014, 35, 7035–7045. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, E.; Webb, A.; Hughes, F.M., Jr. Water movement during apoptosis: A role for aquaporins in the apoptotic volume decrease (AVD). Adv. Exp. Med. Biol. 2004, 559, 179–188. [Google Scholar] [PubMed]
- Jablonski, E.M.; Mattocks, M.A.; Sokolov, E.; Koniaris, L.G.; Hughes, F.M., Jr.; Fausto, N.; Pierce, R.H.; McKillop, I.H. Decreased aquaporin expression leads to increased resistance to apoptosis in hepatocellular carcinoma. Cancer Lett. 2007, 250, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Johnstone, R.W.; Ruefli, A.A.; Tainton, K.M.; Smyth, M.J. A role for P-glycoprotein in regulating cell death. Leuk. Lymphoma 2000, 38, 1–11. [Google Scholar] [PubMed]
- Yang, X.; Page, M. P-glycoprotein expression in ovarian cancer cell line following treatment with cisplatin. Oncol. Res. 1995, 7, 619–624. [Google Scholar] [PubMed]
- Litman, T.; Nielsen, D.; Skovsgaard, T.; Zeuthen, T.; Stein, W.D. ATPase activity of P-glycoprotein related to emergence of drug resistance in Ehrlich ascites tumor cell lines. Biochim. Biophys. Acta 1997, 1361, 147–158. [Google Scholar] [CrossRef]
- Barakat, S.; Demeule, M.; Pilorget, A.; Regina, A.; Gingras, D.; Baggetto, L.G.; Beliveau, R. Modulation of P-glycoprotein function by caveolin-1 phosphorylation. J. Neurochem. 2007, 101, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Barakat, S.; Turcotte, S.; Demeule, M.; Lachambre, M.P.; Regina, A.; Baggetto, L.G.; Beliveau, R. Regulation of brain endothelial cells migration and angiogenesis by P-glycoprotein/caveolin-1 interaction. Biochem. Biophys. Res. Commun. 2008, 372, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Demeule, M.; Jodoin, J.; Gingras, D.; Beliveau, R. P-glycoprotein is localized in caveolae in resistant cells and in brain capillaries. FEBS Lett. 2000, 466, 219–224. [Google Scholar] [CrossRef]
- Pedersen, S.F.; Hoffmann, E.K.; Novak, I. Cell volume regulation in epithelial physiology and cancer. Front. Physiol. 2013, 4, 233. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Arai, S. Two distinct controls of mitotic cdk1/cyclin B1 activity requisite for cell growth prior to cell division. Cell Cycle 2007, 6, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Sveiczer, A.; Novak, B.; Mitchison, J.M. The size control of fission yeast revisited. J. Cell Sci. 1996, 109, 2947–2957. [Google Scholar] [PubMed]
- Wells, W.A. Does size matter? J. Cell Biol. 2002, 158, 1156–1159. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S. Control and maintenance of mammalian cell size. BMC Cell Biol. 2004, 5. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.; Tyers, M. How cells coordinate growth and division. Curr. Biol. 2004, 14, R1014–R1027. [Google Scholar] [CrossRef] [PubMed]
- Saucedo, L.J.; Edgar, B.A. Why size matters: Altering cell size. Curr. Opin. Genet. Dev. 2002, 12, 565–571. [Google Scholar] [CrossRef]
- Conlon, I.; Raff, M. Control and maintenance of mammalian cell size: Response. BMC Cell Biol. 2004, 5. [Google Scholar] [CrossRef] [PubMed]
- Umen, J.G. The elusive sizer. Curr. Opin. Cell Biol. 2005, 17, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Ernest, N.J.; Habela, C.W.; Sontheimer, H. Cytoplasmic condensation is both necessary and sufficient to induce apoptotic cell death. J. Cell Sci. 2008, 121 Pt 3, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Habela, C.W.; Sontheimer, H. Cytoplasmic volume condensation is an integral part of mitosis. Cell Cycle 2007, 6, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Vo, T.; Hajar, A.; Li, S.; Chen, X.; Parissenti, A.M.; Brindley, D.N.; Wang, Z. Multiple mechanisms underlying acquired resistance to taxanes in selected docetaxel-resistant MCF-7 breast cancer cells. BMC Cancer 2014, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Briasoulis, E.; Linardou, H.; Bafaloukos, D.; Papadimitriou, C. Taxane resistance in breast cancer: Mechanisms, predictive biomarkers and circumvention strategies. Cancer Treat. Rev. 2012, 38, 890–903. [Google Scholar] [CrossRef] [PubMed]
- Maulard, C.; Marie, J.P.; Delanian, S.; Housset, M. Clinical effects on CA 15-3 level of cyclosporin A as a chemosensitiser in chemoresistant metastatic breast cancer. Eur. J. Cancer 1993, 29A, 480. [Google Scholar] [CrossRef]
- Le Foll, F.; Rioult, D.; Boussa, S.; Pasquier, J.; Dagher, Z.; Leboulenger, F. Characterisation of Mytilus edulis hemocyte subpopulations by single cell time-lapse motility imaging. Fish Shellfish Immunol. 2010, 28, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Rioult, D.; Pasquier, J.; Boulange-Lecomte, C.; Poret, A.; Abbas, I.; Marin, M.; Minier, C.; le Foll, F. The multi-xenobiotic resistance (MXR) efflux activity in hemocytes of Mytilus edulis is mediated by an ATP binding cassette transporter of class C (ABCC) principally inducible in eosinophilic granulocytes. Aquat. Toxicol. 2014, 153, 98–109. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasquier, J.; Rioult, D.; Abu-Kaoud, N.; Hoarau-Véchot, J.; Marin, M.; Foll, F.L. Evidence for P-Glycoprotein Involvement in Cell Volume Regulation Using Coulter Sizing in Flow Cytometry. Int. J. Mol. Sci. 2015, 16, 14318-14337. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160714318
Pasquier J, Rioult D, Abu-Kaoud N, Hoarau-Véchot J, Marin M, Foll FL. Evidence for P-Glycoprotein Involvement in Cell Volume Regulation Using Coulter Sizing in Flow Cytometry. International Journal of Molecular Sciences. 2015; 16(7):14318-14337. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160714318
Chicago/Turabian StylePasquier, Jennifer, Damien Rioult, Nadine Abu-Kaoud, Jessica Hoarau-Véchot, Matthieu Marin, and Frank Le Foll. 2015. "Evidence for P-Glycoprotein Involvement in Cell Volume Regulation Using Coulter Sizing in Flow Cytometry" International Journal of Molecular Sciences 16, no. 7: 14318-14337. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160714318