
Identification of Rhopalosiphum Padi Virus 5′ Untranslated Region Sequences Required for Cryptic Promoter Activity and Internal Ribosome Entry

Abstract
:1. Introduction
2. Results and Discussion
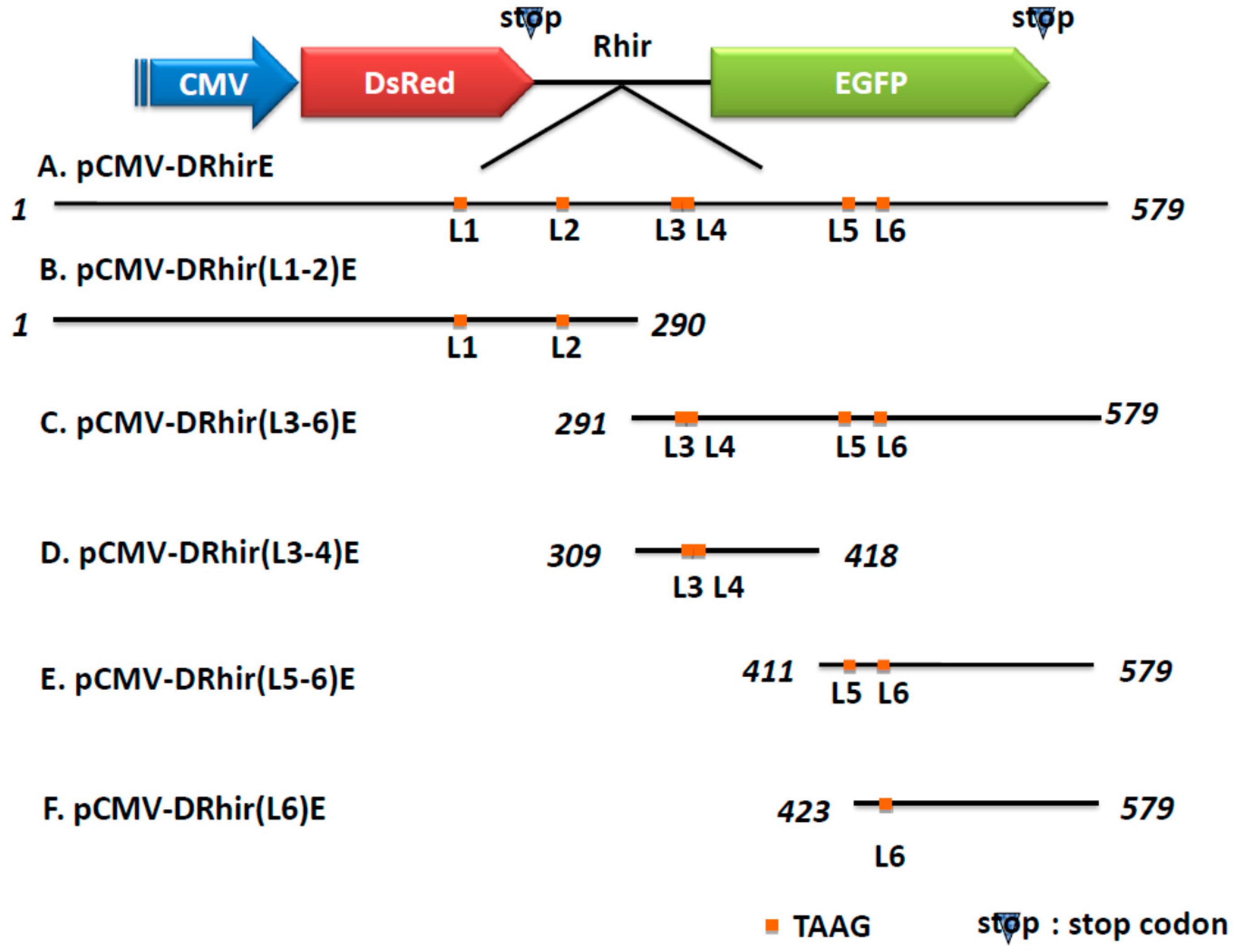
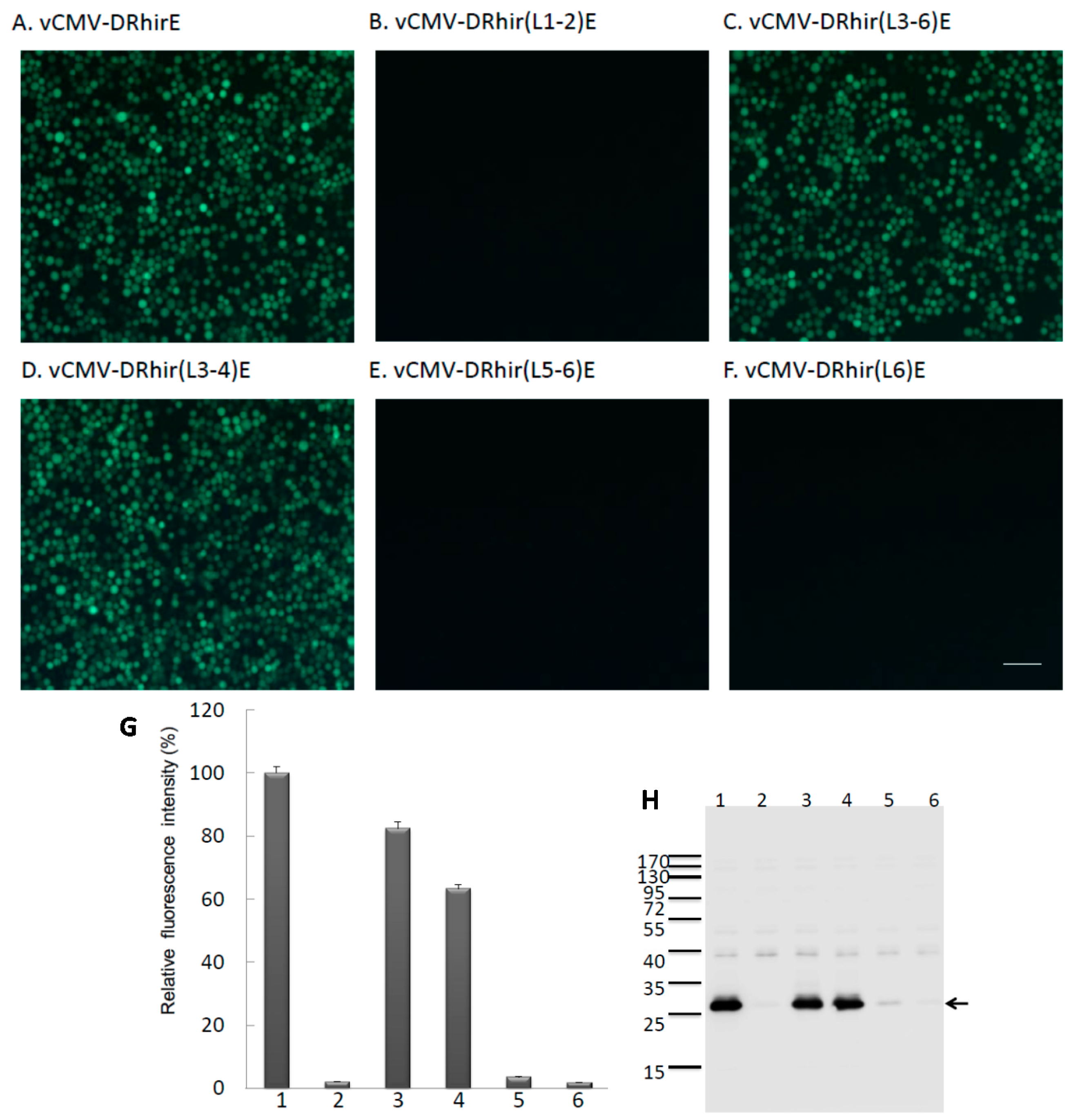
2.1. Tandem Repeated TAAG Motifs Are Responsible for the Promoter Activity of RhPV IRES in Baculovirus Infected Sf21 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Nucleotide * Position |
|---|---|---|
| RH001-290F | GGCGAATTCGATAAAAGAACCTATAATC | 1–19 |
| RH001-290R | GGCCGGGGATCCGAATAAAATATAATAAAATAG | 270–290 |
| RH291-579F | GGAGAATTCACCCCCCACATTAATCCC | 291–308 |
| RH291-579R | GGCGGATCCCGGGTATAAATAGATAAAG | 564–579 |
| RH309-418F | GGCCGGGAATTCAGTTAAAGCTTTATAAC | 309–325 |
| RH309-418R | GGCGGATCCACTAAAAATTGTGAAAAATA | 399–418 |
| RH411-579F | GGCCGGGAATTCAATTTTTAGTTAAGATTTTAGC | 411–432 |
| RH291-579R | GGCGGATCCCGGGTATAAATAGATAAAG | 564–579 |
| RH423-579F | GGCGAATTCATTTTAGCTTGCCTTAAG | 423–440 |
| RH291-579R | GGCGGATCCCGGGTATAAATAGATAAAG | 564–579 |



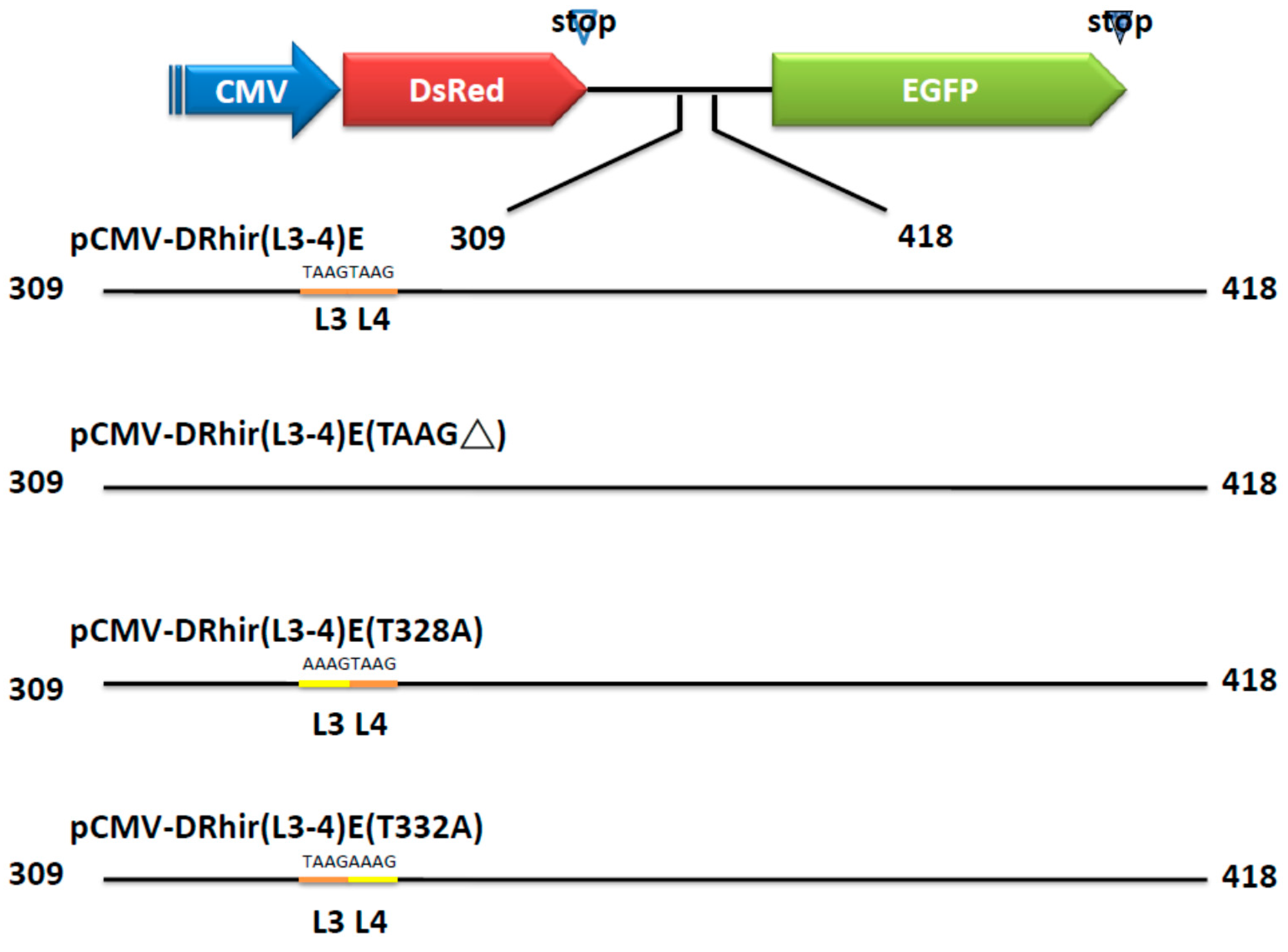
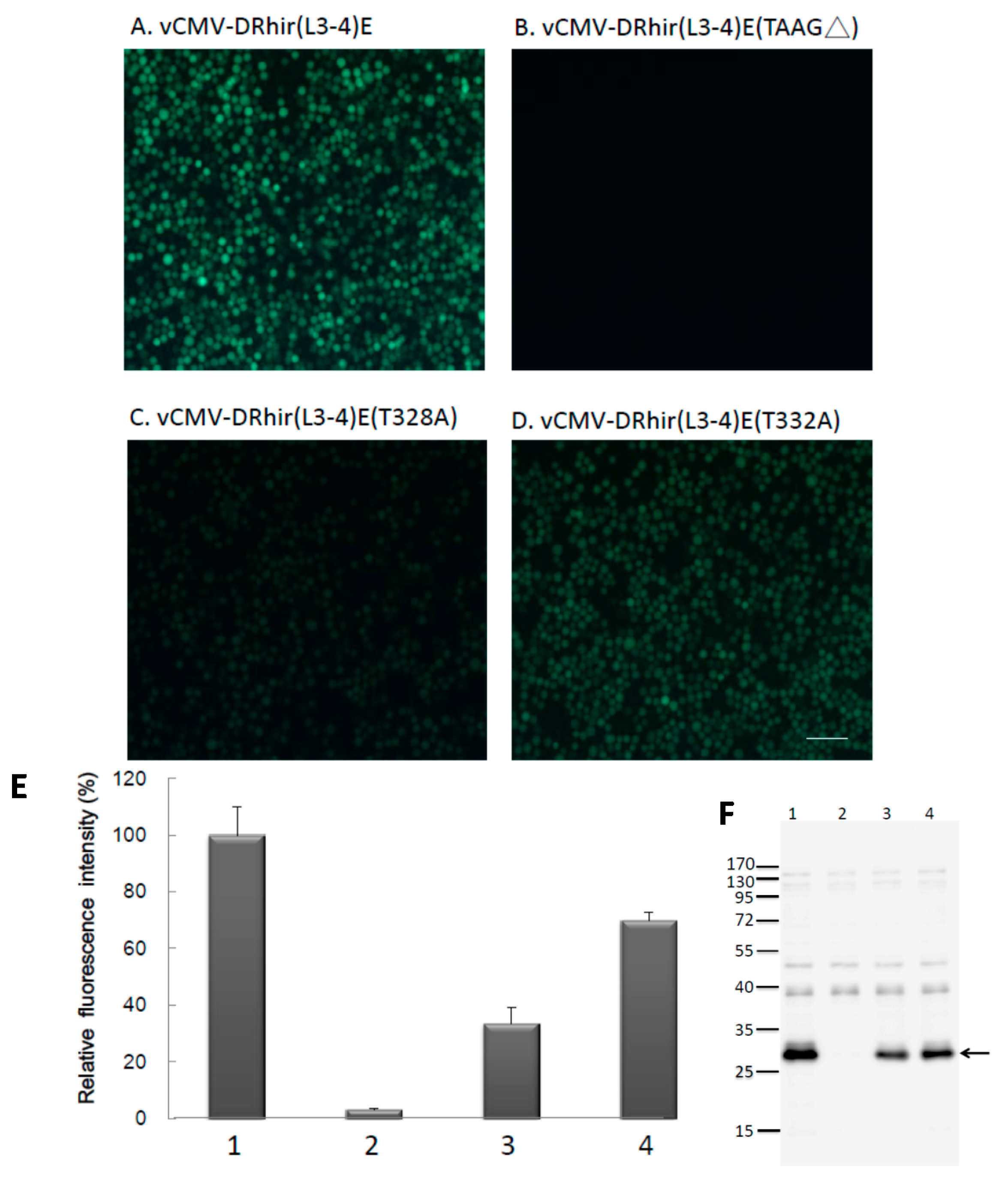
2.2. TAAG Motif in L3 Is More Critical for RP110 Promoter Activity in the Baculoviruses Infected Sf21 Cells

2.3. RP110 Acts as an IRES in CHO Cells

3. Experimental Section
3.1. Cells
3.2. Construction of Plasmids
3.3. Recombinant Virus Production and Titer Determination
3.4. Western Blot Analysis
3.5. Transfection of CHO-K1 Cells
3.6. EGFP Reporter Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.K.; Pestova, T.V.; Hellen, C.U.; Witherell, G.W.; Wimmer, E. Cap-independent translation of picornavirus RNAs: Structure and function of the internal ribosomal entry site. Enzyme 1990, 44, 292–309. [Google Scholar] [PubMed]
- Pelletier, J.; Sonenberg, N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature 1988, 334, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Hellen, C.U.; Sarnow, P. Internal ribosome entry sites in eukaryotic mRNA molecules. Genes Dev. 2001, 15, 1593–1612. [Google Scholar] [CrossRef] [PubMed]
- Dorokhov, Y.L.; Skulachev, M.V.; Ivanov, P.A.; Zvereva, S.D.; Tjulkina, L.G.; Merits, A.; Gleba, Y.Y.; Hohn, T.; Atabekov, J.G. Polypurine (A)-rich sequences promote cross-kingdom conservation of internal ribosome entry. Proc. Natl. Acad. Sci. USA 2002, 99, 5301–5306. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Lefebvre, C.; Yeh, C.; Chow, T.; Korneluk, R.G. A new internal-ribosome-entry-site motif potentiates XIAP-mediated cytoprotection. Nat. Cell Biol. 1999, 1, 190–192. [Google Scholar] [PubMed]
- Sachs, A.B. Cell cycle-dependent translation initiation: IRES elements prevail. Cell 2000, 101, 243–245. [Google Scholar] [CrossRef]
- Werner, R. IRES elements in connexin genes: A hypothesis explaining the need for connexins to be regulated at the translational level. IUBMB Life 2000, 50, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.A. A summary of taxonomic changes recently approved by ICTV. Arch. Virol. 2002, 147, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Domier, L.L.; McCoppin, N.K. In vivo activity of Rhopalosiphum padi virus internal ribosome entry sites. J. Gen. Virol. 2003, 84, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Jan, E. Divergent IRES elements in invertebrates. Virus Res. 2006, 119, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Domier, L.L.; McCoppin, N.K.; D’Arcy, C.J. Sequence requirements for translation initiation of Rhopalosiphum padi virus ORF2. Virology 2000, 268, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, J.; Nakashima, N. Translation initiation at the CUU codon is mediated by the internal ribosome entry site of an insect picorna-like virus in vitro. J. Virol. 1999, 73, 1219–1226. [Google Scholar] [PubMed]
- Wilson, J.E.; Powell, M.J.; Hoover, S.E.; Sarnow, P. Naturally occurring dicistronic cricket paralysis virus RNA is regulated by two internal ribosome entry sites. Mol. Cell. Biol. 2000, 20, 4990–4999. [Google Scholar] [CrossRef] [PubMed]
- Woolaway, K.E.; Lazaridis, K.; Belsham, G.J.; Carter, M.J.; Roberts, L.O. The 5′ untranslated region of Rhopalosiphum padi virus contains an internal ribosome entry site which functions efficiently in mammalian, plant, and insect translation systems. J. Virol. 2001, 75, 10244–10249. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.O.; Groppelli, E. An atypical IRES within the 5′UTR of a dicistrovirus genome. Virus Res. 2009, 139, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Terenin, I.M.; Dmitriev, S.E.; Andreev, D.E.; Royall, E.; Belsham, G.J.; Roberts, L.O.; Shatsky, I.N. A cross-kingdom internal ribosome entry site reveals a simplified mode of internal ribosome entry. Mol. Cell. Biol. 2005, 25, 7879–7888. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, C.J.; Burnett, P.A.; Hewings, A.D. Detection, biological effects, and transmission of a virus of the aphid Rhopalosiphum padi. Virology 1981, 114, 268–272. [Google Scholar] [CrossRef]
- Wu, Y.; Teng, C.; Chen, Y.; Chen, S.; Chen, Y.; Lin, Y.; Wu, T. Internal ribosome entry site of Rhopalosiphum padi virus is functional in mammalian cells and has cryptic promoter activity in baculovirus-infected Sf21 cells. Acta Pharmacol. Sin. 2008, 29, 965–974. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, D.R.; Miller, L.K.; Luckow, V.A. Baculovirus Expression Vectors: A Laboratory Manual; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Liu, Z.; Dong, Z.; Han, B.; Yang, Y.; Liu, Y.; Zhang, J.-T. Regulation of expression by promoters versus internal ribosome entry site in the 5′-untranslated sequence of the human cyclin-dependent kinase inhibitor p27kip1. Nucleic Acids Res. 2005, 33, 3763–3771. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Weaver, M.; Magnuson, N.S. Cryptic promoter activity in the DNA sequence corresponding to the pim-1 5′UTR. Nucleic Acids Res. 2005, 33, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Dumas, E.; Staedel, C.; Colombat, M.; Reigadas, S.; Chabas, S.; Astier-Gin, T.; Cahour, A.; Litvak, S.; Ventura, M. A promoter activity is present in the DNA sequence corresponding to the hepatitis C virus 5′UTR. Nucleic Acids Res. 2003, 31, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Groppelli, E.; Belsham, G.J.; Roberts, L.O. Identification of minimal sequences of the Rhopalosiphum padi virus 5′ untranslated region required for internal initiation of protein synthesis in mammalian, plant and insect translation systems. J. Gen. Virol. 2007, 88, 1583–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiem, S.M.; Miller, L.K. Differential gene expression mediated by late, very late and hybrid baculovirus promoters. Gene 1990, 91, 87–94. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Chen, W.-S.; Wu, T.-Y. Development of a bi-cistronic baculovirus expression vector by the Rhopalosiphum padi virus 5′ internal ribosome entry site. Biochem. Biophys. Res. Commun. 2005, 335, 616–623. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.-K.; Lin, J.-Z.; Jinn, T.-R.; Chan, H.-L.; Wu, T.-Y. Identification of Rhopalosiphum Padi Virus 5′ Untranslated Region Sequences Required for Cryptic Promoter Activity and Internal Ribosome Entry. Int. J. Mol. Sci. 2015, 16, 16053-16066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160716053
Liu M-K, Lin J-Z, Jinn T-R, Chan H-L, Wu T-Y. Identification of Rhopalosiphum Padi Virus 5′ Untranslated Region Sequences Required for Cryptic Promoter Activity and Internal Ribosome Entry. International Journal of Molecular Sciences. 2015; 16(7):16053-16066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160716053
Chicago/Turabian StyleLiu, Ming-Kun, Jie-Zue Lin, Tzyy-Rong Jinn, Hong-Lin Chan, and Tzong-Yuan Wu. 2015. "Identification of Rhopalosiphum Padi Virus 5′ Untranslated Region Sequences Required for Cryptic Promoter Activity and Internal Ribosome Entry" International Journal of Molecular Sciences 16, no. 7: 16053-16066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160716053