
Inhibitory Effects of Amorphigenin on the Mitochondrial Complex I of Culex pipiens pallens Coquillett (Diptera: Culicidae)

Abstract
:1. Introduction

2. Results and Discussion
2.1. Results
2.1.1. In Vivo and in Vitro Inhibition of the Mitochondrial Complex I Activity in Cx. pipiens pallens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Delivery | Compounds | Regression Equation | IC50 (μmol/L) | R Value |
|---|---|---|---|---|
| In vivo | Amorphigenin | y = 4.0495 + 2.4616x | 2.4329 | 0.9046 |
| Rotenone | y = 3.9764 + 2.5466x | 2.5232 | 0.9152 | |
| In vitro | Amorphigenin | y = 3.0076 + 4.3668x | 2.8592 | 0.9884 |
| Rotenone | y = 3.2495 + 3.5250x | 3.1375 | 0.9982 |
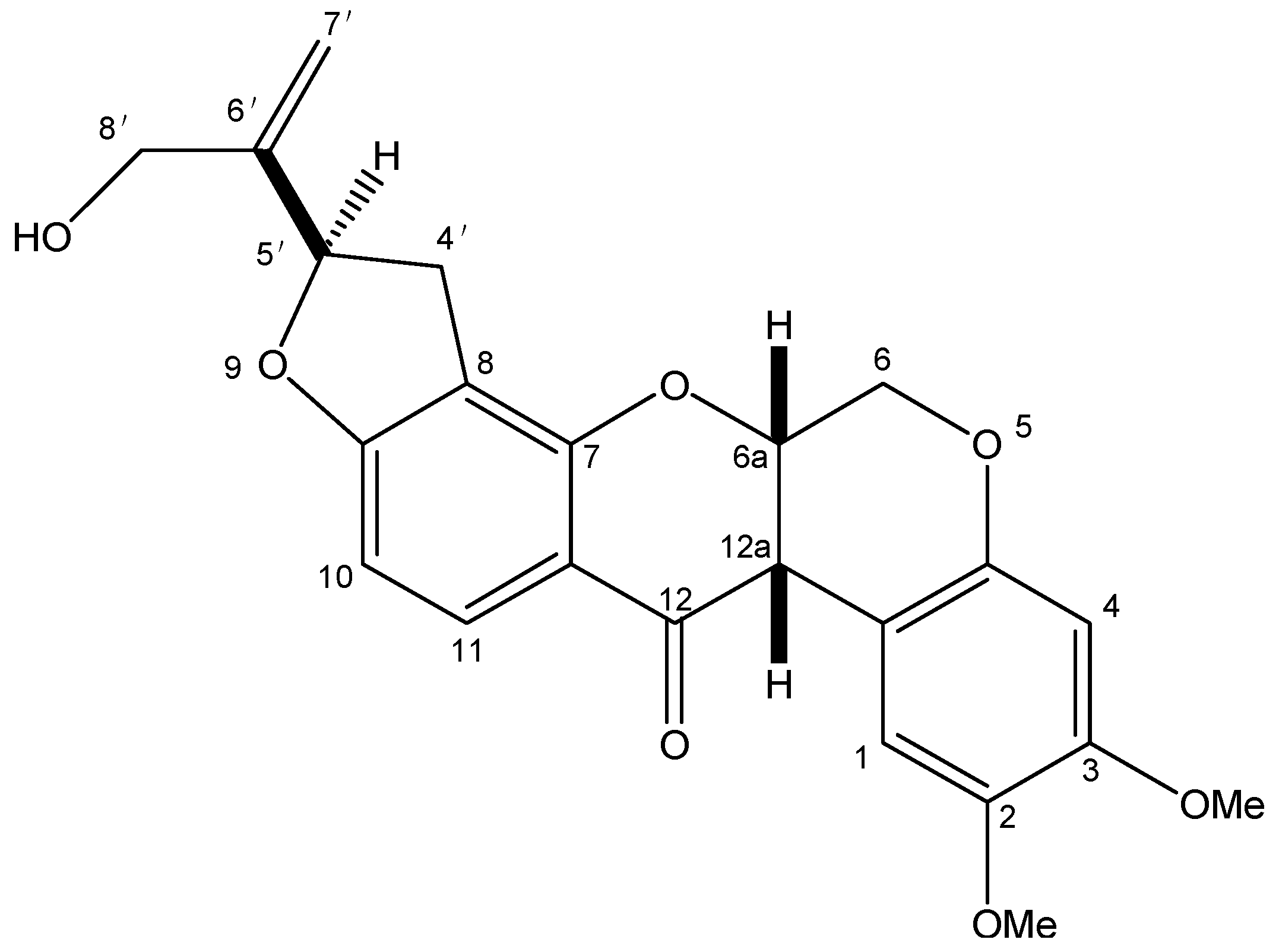
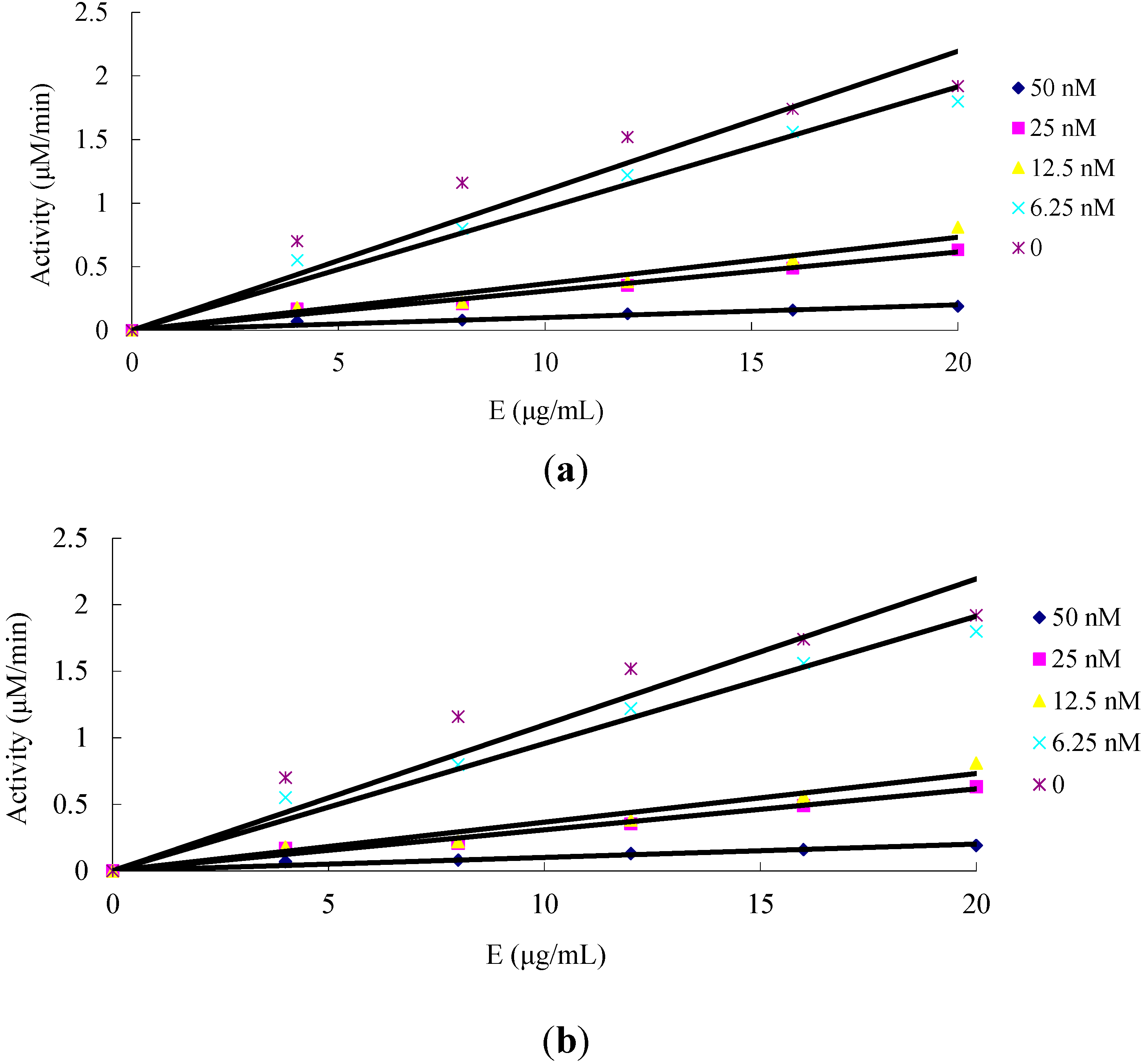
2.1.2. The Effect of Amorphigenin and Rotenone on the Mitochondrial Complex I

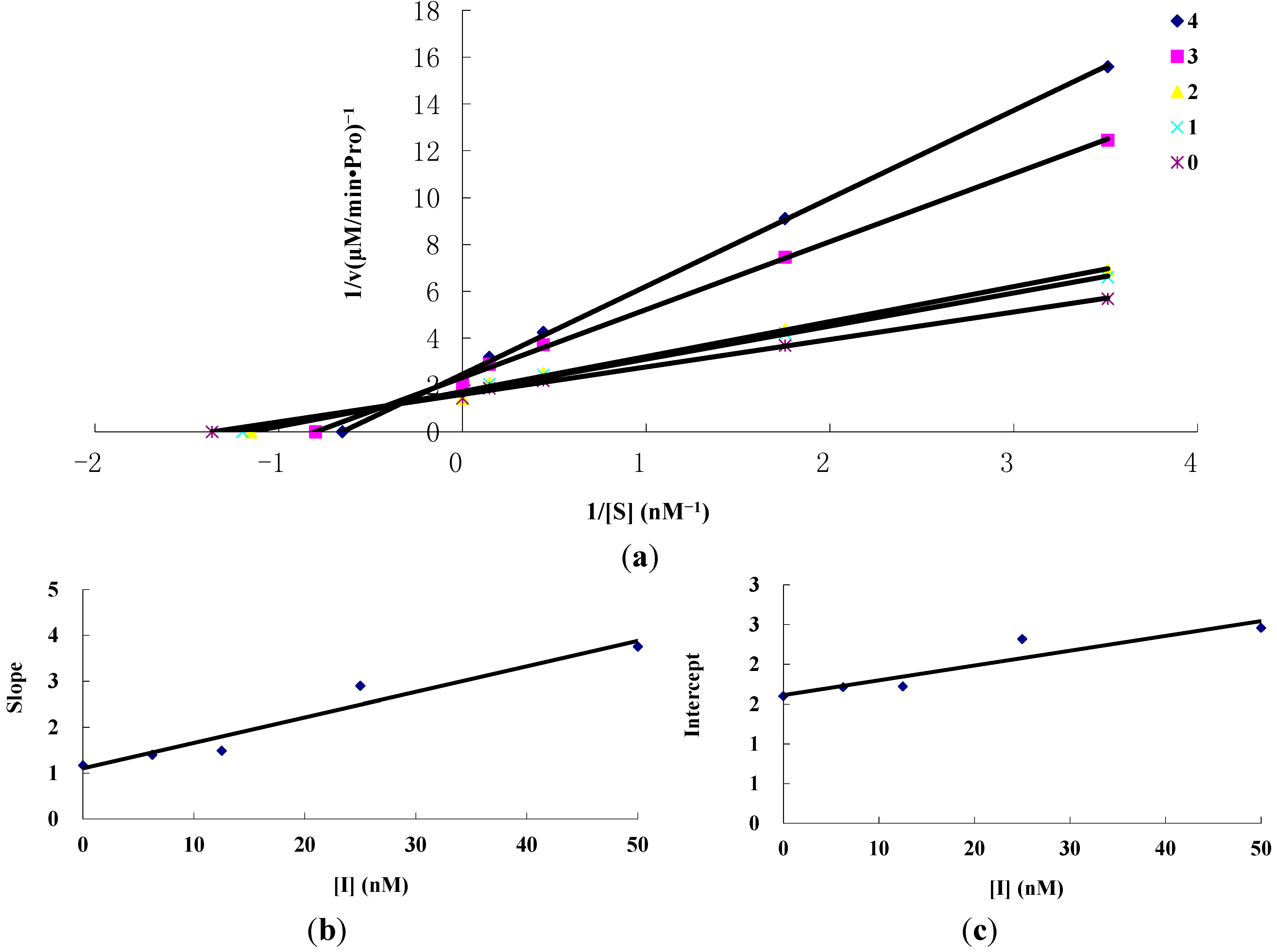
2.1.3. Inhibition Kinetics of Amorphigenin and Rotenone on the Mitochondrial Complex I Activity


2.2. Discussion

3. Experimental Section
3.1. Chemicals
3.2. Mosquito Culture
3.3. Isolation of Mitochondria
3.4. Assay of the Mitochondrial Complex I Activity and in Vitro Inhibition
3.5. Determination of the Type and Constant of Amorphigenin and Rotenone Inhibition of the Mitochondrial Complex I Activity
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Li, X.L.; Ma, L.; Sun, L.X.; Zhu, C.L. Biotic characteristics in the deltamethrin-susceptible and resistant strains of Culex pipiens pallens (Diptera: Culicidae) in China. Appl. Entomol. Zool. 2002, 37, 305–308. [Google Scholar] [CrossRef]
- Jiang, S.F.; Zhao, T.Y.; Zhang, Y.M.; Dong, Y.D.; Guo, X.X. RT-PCR detection of West Nile virus in mosquitoes and leghorn chicken infected experimentally. Acta Parasitol. Med. Entomol. Sin. 2006, 13, 21–24. [Google Scholar]
- Scott, J.G.; Yoshimizu, M.H.; Kasai, S. Pyrethroid resistance in Culex pipiens mosquitoes. Pestic. Biochem. Physiol. 2015, 120, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Lin, L.F.; Qiao, C.L.; Xu, Y.; Marquine, M.; Weill, M.; Raymond, M. Insecticide resistance in Chinese populations of the Culex pipiens complex through esterase overproduction. Entomol. Exp. Appl. 2006, 120, 211–220. [Google Scholar] [CrossRef]
- Cui, F.; Raymond, M.; Berthomieu, A.; Alout, H.; Weill, M.; Qiao, C.-L. Recent emergence of insensitive acetylcholinesterase in Chinese populations of the mosquito Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2006, 43, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Tan, Y.; Qiao, C.-L. Filariasis vector in China: Insecticide resistance and population structure of mosquito Culex pipiens complex. Pest Manag. Sci. 2007, 63, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.J.; Chang, K.S.; Lee, W.J.; Ahn, Y.J. Monitoring of insecticide resistance in field-collected populations of Culex pipiens pallens (Diptera: Culicidae). J. Asia Pac. Entomol. 2007, 10, 257–261. [Google Scholar] [CrossRef]
- Shin, E.H.; Kim, N.J.; Kim, H.K.; Park, C.; Lee, D.K.; Ahn, Y.J.; Chang, K.S. Resistance of field-collected populations of Culex pipiens pallens (Diptera: Culicidae) to insecticides in the Republic of Korea. J. Asia Pac. Entomol. 2012, 15, 1–4. [Google Scholar] [CrossRef]
- Ghosh, A.; Chowdhury, N.; Chandra, G. Plant extracts as potential mosquito larvicides. Indian J. Med. Res. 2012, 135, 581–598. [Google Scholar] [PubMed]
- Shaalan, E.A.-S.; Canyon, D.; Younes, M.W.F.; Abdel-Wahab, H.; Mansour, A.-H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 31, 1149–1166. [Google Scholar] [CrossRef] [PubMed]
- Kishore, N.; Mishra, B.; Tiwari, V.; Tripathi, V.; Lall, N. Natural products as leads to potential mosquitocides. Phytochem. Rev. 2014, 13, 587–627. [Google Scholar] [CrossRef]
- Kembro, J.; Marin, R.; Zygadlo, J.; Gleiser, R. Effects of the essential oils of Lippia turbinata and lippia polystachya (Verbenaceae) on the temporal pattern of locomotion of the mosquito Culex quinquefasciatus (Diptera: Culicidae) larvae. Parasitol. Res. 2009, 104, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Agra-Neto, A.; Napoleão, T.; Pontual, E.; de Lima Santos, N.; de Andrade Luz, L.; de Oliveira, C.; de Melo-Santos, M.; Coelho, L.; do Amaral Ferraz Navarro, D.; Paiva, P. Effect of Moringa oleifera lectins on survival and enzyme activities of Aedes aegypti larvae susceptible and resistant to organophosphate. Parasitol. Res. 2014, 113, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Pontual, E.; de Lima Santos, N.; de Moura, M.; Coelho, L.; do Amaral Ferraz Navarro, D.; Napoleão, T.; Paiva, P. Trypsin inhibitor from Moringa oleifera flowers interferes with survival and development of Aedes aegypti larvae and kills bacteria inhabitant of larvae midgut. Parasitol. Res. 2014, 113, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Kotewong, R.; Duangkaew, P.; Srisook, E.; Sarapusit, S.; Rongnoparut, P. Structure–function relationships of inhibition of mosquito cytochrome p450 enzymes by flavonoids of Andrographis paniculata. Parasitol. Res. 2014, 113, 3381–3392. [Google Scholar] [CrossRef] [PubMed]
- Nunes, F.; Leite, J.; Oliveira, L.G.; Sousa, P.P.S.; Menezes, M.; Moraes, J.S.; Mascarenhas, S.; Braga, V. The larvicidal activity of Agave sisalana against l4 larvae of Aedes aegypti is mediated by internal necrosis and inhibition of nitric oxide production. Parasitol. Res. 2015, 114, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Koodalingam, A.; Mullainadhan, P.; Arumugam, M. Effects of extract of soapnut Sapindus emarginatus on esterases and phosphatases of the vector mosquito, Aedes aegypti (Diptera: Culicidae). Acta Trop. 2011, 118, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef]
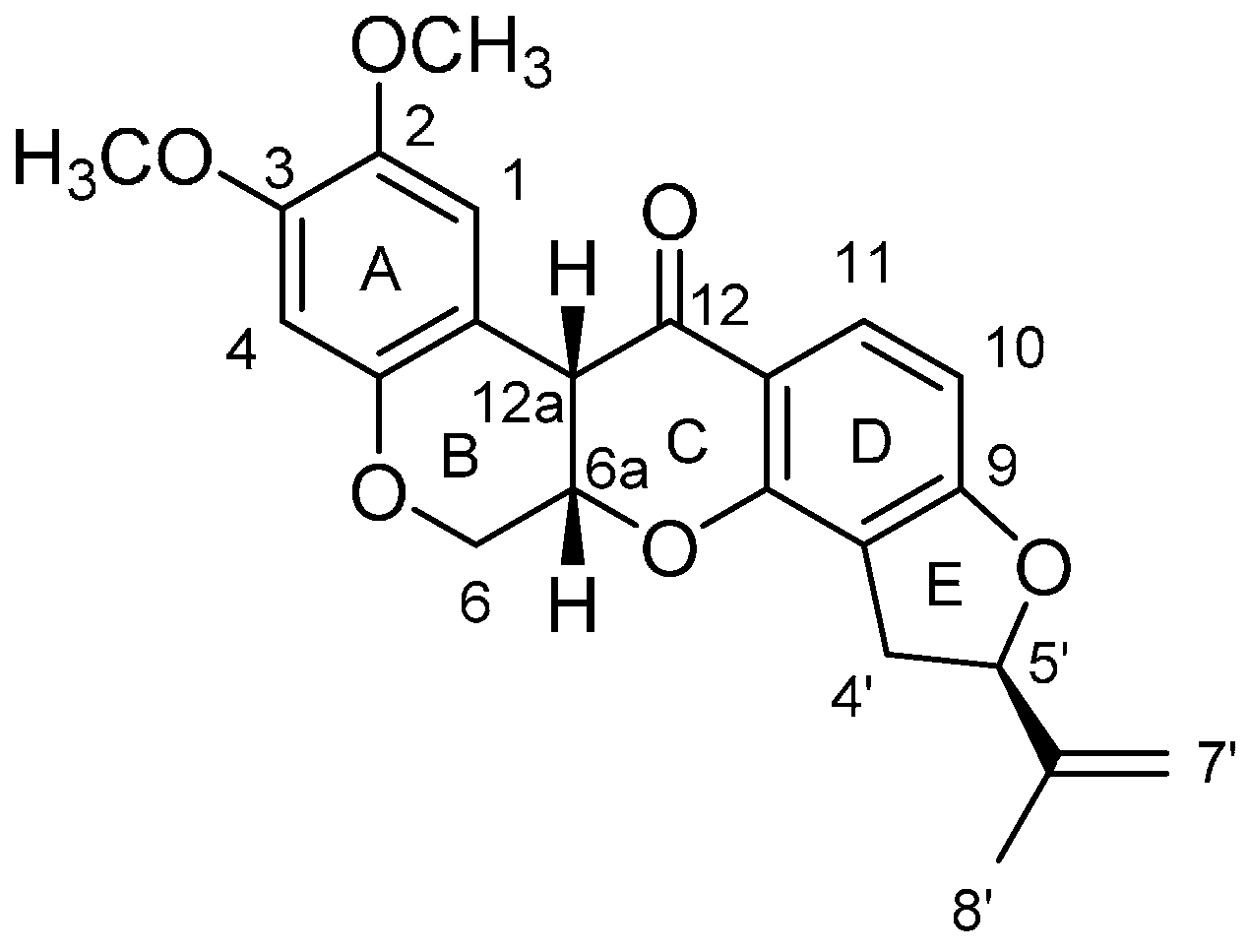
- Kondratenko, E.S.; Kasymov, A.U.; Abubakirov, N.K. Structure of amorphigenin. Chem. Nat. Compd. 1967, 3, 260–262. [Google Scholar] [CrossRef]
- Kasymov, A.U.; Kondratenko, E.S.; Abubakirov, N.K. Stepwise hydrolysis of amorphin. Chem. Nat. Compd. 1970, 6, 482–482. [Google Scholar] [CrossRef]
- Crombie, L.; Dewick, P.M.; Whiting, D.A. Biosynthesis of rotenoids. Chalcone, isoflavone, and rotenoid stages in the formation of amorphigenin by Amorpha fruticosa seedlings. J. Chem. Soc. Perkin Trans. 1973, 1, 1285–1294. [Google Scholar] [CrossRef]
- Kadyrova, F.R.; Shamsutdinov, M.R.I.; Shakirov, T.T. The isolation of fruticin from the seeds of Amorpha fruticosa. Chem. Nat. Compd. 1973, 9, 107–107. [Google Scholar] [CrossRef]
- Li, L.; Wang, H.K.; Chang, J.J.; McPhail, A.T.; McPhail, D.R.; Terada, H.; Konoshima, T.; Kokumai, M.; Kozuka, M.; Estes, J.R.; et al. Antitumor agents, 138. Rotenoids and isoflavones as cytotoxic constitutents from Amorpha fruticosa. J. Nat. Prod. 1993, 56, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Konoshima, T.; Terada, H.; Kokumai, M.; Kozuka, M.; Tokuda, H.; Estes, J.R.; Li, L.; Wang, H.K.; Lee, K.H. Studies on inhibitors of skin tumor promotion, XII. Rotenoids from Amorpha fruticosa. J. Nat. Prod. 1993, 56, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Kwak, H.B.; Choi, E.Y.; Kim, H.S.; Kim, M.H.; Kim, S.H.; Choi, M.K.; Chun, C.H.; Oh, J.; Kim, J.J. Amorphigenin inhibits Osteoclast differentiation by suppressing c-Fos and nuclear factor of activated T cells. Anat. Cell Biol. 2010, 43, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Kloutek, E.; Popov, A.; Drenska, D.; Uzunov, P. Experimental research on the hepatoprotective activity of flavonoids isolated from Amorpha fructiosa. Eksp. Med. Morfol. 1985, 24, 50–54. [Google Scholar] [PubMed]
- Kim, Y.S.; Ryu, Y.B.; Curtis-Long, M.J.; Yuk, H.J.; Cho, J.K.; Kim, J.Y.; Kim, K.D.; Lee, W.S.; Park, K.H. Flavanones and rotenoids from the roots of Amorpha fruticosa L. That inhibit bacterial neuraminidase. Food Chem. Toxicol. 2011, 49, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Brett, C.H. Repellent properties of extract of Amorpha fruticosa. J. Econ. Entomol. 1946, 39, 810. [Google Scholar] [CrossRef] [PubMed]
- Brett, C.H. Insecticidal properties of the indigobush (Amorpha fruticosa). J. Agric. Res. 1946, 73, 81–96. [Google Scholar] [PubMed]
- Abe, F.; Donnelly, D.M.X.; Moretti, C.; Polonsky, J. Isoflavanoid constituents from Dalbergia monetaria. Phytochemistry 1985, 24, 1071–1076. [Google Scholar] [CrossRef]
- Ji, M.; Liu, C.; Li, X.; Liu, D.; Wu, D.; Wang, Y. Insecticidal and antifeeding activity of seeds of Amorpha fruticosa against Schizaphis graminum. Jiangsu Agric. Sci. 2011, 39, 208–210. [Google Scholar]
- Liang, Y.; Li, X.; Gu, Z.; Qin, P.; Ji, M. Toxicity of amorphigenin from the seeds of Amorpha fruticosa against the larvae of Culex pipiens pallens (Diptera: Culicidae). Molecules 2015, 20, 3238–3254. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Miyoshi, H.; Ebisui, K.; Iwamura, H. Comparison of the inhibitory action of natural rotenone and its stereoisomers with various NADH-ubiquinone reductases. Eur. J. Biochem. 1994, 225, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zeng, X.-N.; Ma, L.-M.; Ru, H.-K. Mechanism of epirotenone enantiomorphs as insecticides. Sci. Agric. Sin. 2007, 40, 2747–2275. [Google Scholar]
- Hatefi, Y. The mitochondrial electron transport and oxidative phosphorylation system. Annu. Rev. Biochem. 1985, 54, 1015–1069. [Google Scholar] [CrossRef] [PubMed]
- Hollingworth, R.M.; Ahammadsahib, K.I.; Gadelhak, G.; McLaughlin, J.L. New inhibitors of complex I of the mitochondrial electron transport chain with activity as pesticides. Biochem. Soc. Trans. 1994, 22, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Tormo, J.R.; González, M.C.; Cortes, D.; Estornell, E. Kinetic characterization of mitochondrial complex I inhibitors using annonaceous acetogenins. Arch. Biochem. Biophys. 1999, 369, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Earley, F.G.; Ragan, C.I. Photoaffinity labelling of mitochondrial NADH dehydrogenase with arylazidoamorphigenin, an analogue of rotenone. Biochem. J. 1984, 224, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Burgos, J.; Redfearn, E.R. The inhibition of mitochondrial reduced nicotinamide-adenine dinucleotide oxidation by rotenoids. Biochim. Biophys. Acta 1965, 110, 475–483. [Google Scholar] [CrossRef]
- Chen, Q. Enzymology and Research Technology; Xiamen University Press: Xiamen, China, 2010. [Google Scholar]
- Walker, J.E. The NADH:ubiquinone oxidoreductase (complex I) of respiratory chains. Q. Rev. Biophys. 1992, 25, 253–324. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.K.; Bates, R.B.; Grady, R.A.; Delfel, N.E. Crystal and molecular structure of the one to one complex of rotenone and carbon tetrachloride. J. Am. Chem. Soc. 1975, 97, 5752–5755. [Google Scholar] [CrossRef]
- Ueno, H.; Miyoshi, H.; Inoue, M.; Niidome, Y.; Iwamura, H. Structural factors of rotenone required for inhibition of various NADH-ubiquinone oxidoreductases. Biochim. Biophys. Acta 1996, 1276, 195–202. [Google Scholar] [CrossRef]
- Darrouzet, E.; Issartel, J.-P.; Lunardi, J.; Dupuis, A. The 49-kDa subunit of NADH-ubiquinone oxidoreductase (complex I) is involved in the binding of piericidin and rotenone, two quinone-related inhibitors. FEBS Lett. 1998, 431, 34–38. [Google Scholar] [CrossRef]
- Miyoshi, H. Structure–activity relationships of some complex I inhibitors. Biochim. Biophys. Acta 1998, 1364, 236–244. [Google Scholar] [CrossRef]
- Valles, S.M.; Capinera, J.L. Response of larvae of the southern armyworm, Spodoptera eridania cramer lepidoptera: Noctuidae, to selected botanical insecticides and soap. J. Agric. Entomol. 1993, 10, 145–153. [Google Scholar]
- Committee on Strategies for the Management of Pesticide Resistant Pest Populations, B.o.A., National Research Council. Pesticide Resistance: Strategies and Tactics for Management; National Academy Press: Washington, DC, USA, 1986. [Google Scholar]
- Fukami, H.; Nakajima, M. Rotenone and the Rotenoids; Marcel Dekker Inc.: New York, NY, USA, 1971. [Google Scholar]
- Rattanapan, A. Biochemical and Molecular Detection of Cypermethrin and Rotenone Resistance in the Tropical Armywrom, Spodoptera litura (Fabricius); Kasetsart University: Bangkok, Thailand, 2007. [Google Scholar]
- Crombie, L.; Josephs, J.L.; Cayley, J.; Larkin, J.; Weston, J.B. The rotenoid core structure: Modifications to define the requirements of the toxophore. Bioorg. Med. Chem. Lett. 1992, 2, 13–16. [Google Scholar] [CrossRef]
- Lümmen, P. Complex I inhibitors as insecticides and acaricides. Biochim. Biophys. Acta 1998, 1364, 287–296. [Google Scholar] [CrossRef]
- Akbar, S.M.; Sharma, H.C.; Jayalakshmi, S.K.; Sreeramulu, K. Methylparathion- and carbofuran-induced mitochondrial dysfunction and oxidative stress in Helicoverpa armigera (Noctuidae: Lepidoptera). Pestic. Biochem. Physiol. 2012, 103, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Birch-Machin, M.A.; Briggs, H.L.; Saborido, A.A.; Bindoff, L.A.; Turnbull, D.M. An evaluation of the measurement of the activities of complexes I–IV in the respiratory chain of human skeletal muscle mitochondria. Biochem. Med. Metab. Biol. 1994, 51, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Krishnamoorthy, G. The non-equivalence of binding sites of coenzyme quinone and rotenone in mitochondrial NADH-CoQ reductase. FEBS Lett. 1992, 300, 275–278. [Google Scholar] [CrossRef]
- Heinrich, H.; Werner, S. Identification of the ubiquinone-binding site of NADH: Ubiquinone oxidoreductase (complex I) from Neurospora crassa. Biochemistry 1992, 31, 11413–11419. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.S., Jr.; Greenamyre, J.T. [3H]Dihydrorotenone binding to NADH: Ubiquinone reductase (complex I) of the electron transport chain: An autoradiographic study. J. Neurosci. 1996, 16, 3807–3816. [Google Scholar] [PubMed]
- Zhang, J.-P.; Chen, Q.-X.; Song, K.-K.; Xie, J.-J. Inhibitory effects of salicylic acid family compounds on the diphenolase activity of mushroom tyrosinase. Food Chem. 2006, 95, 579–584. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control. Assoc. 1925, 18, 265–267. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, M.; Liang, Y.; Gu, Z.; Li, X. Inhibitory Effects of Amorphigenin on the Mitochondrial Complex I of Culex pipiens pallens Coquillett (Diptera: Culicidae). Int. J. Mol. Sci. 2015, 16, 19713-19727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160819713
Ji M, Liang Y, Gu Z, Li X. Inhibitory Effects of Amorphigenin on the Mitochondrial Complex I of Culex pipiens pallens Coquillett (Diptera: Culicidae). International Journal of Molecular Sciences. 2015; 16(8):19713-19727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160819713
Chicago/Turabian StyleJi, Mingshan, Yaping Liang, Zumin Gu, and Xiuwei Li. 2015. "Inhibitory Effects of Amorphigenin on the Mitochondrial Complex I of Culex pipiens pallens Coquillett (Diptera: Culicidae)" International Journal of Molecular Sciences 16, no. 8: 19713-19727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160819713