
Effects of Low Molecular Weight Yeast β-Glucan on Antioxidant and Immunological Activities in Mice

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
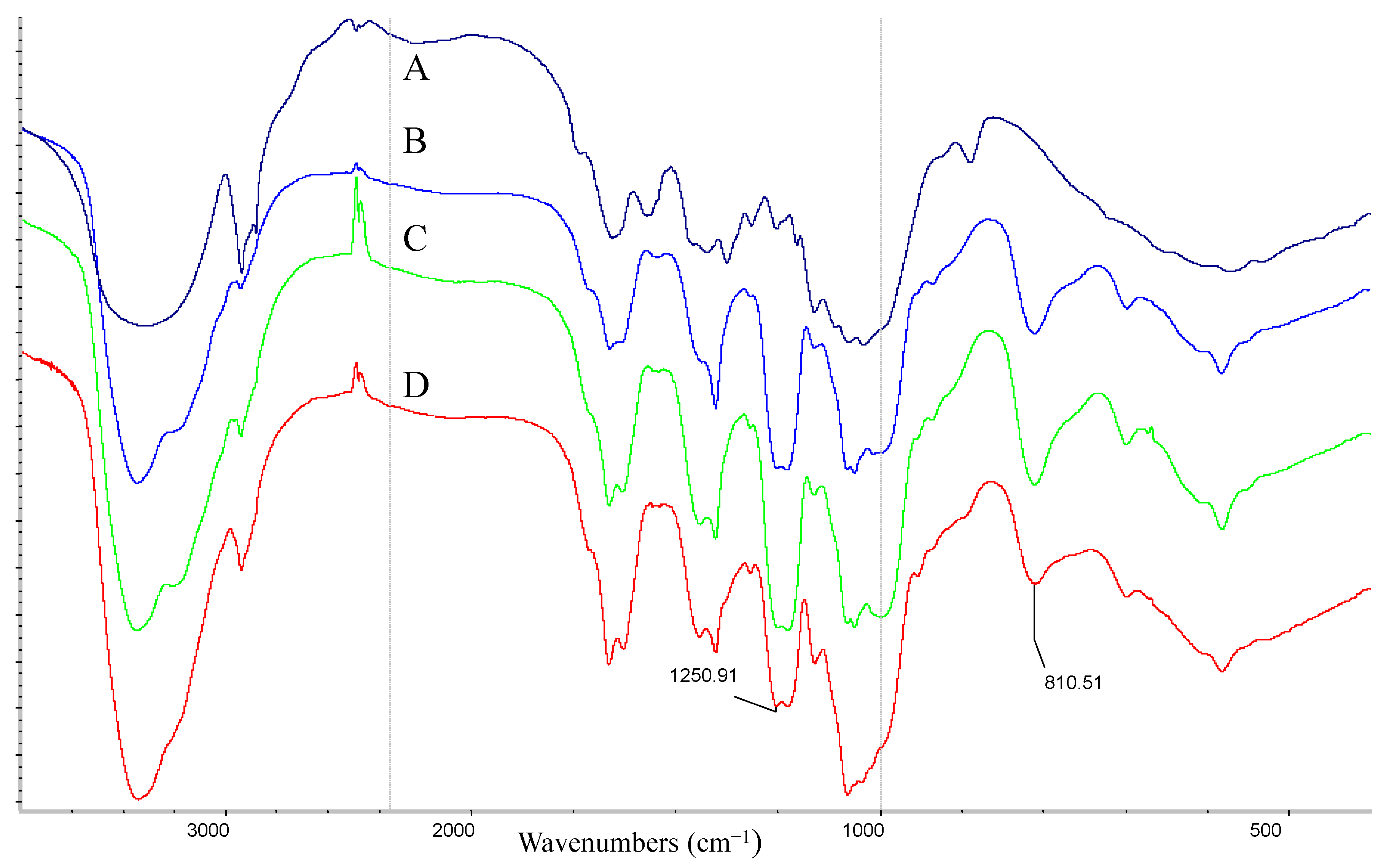
2.1.1. Chemical Analysis

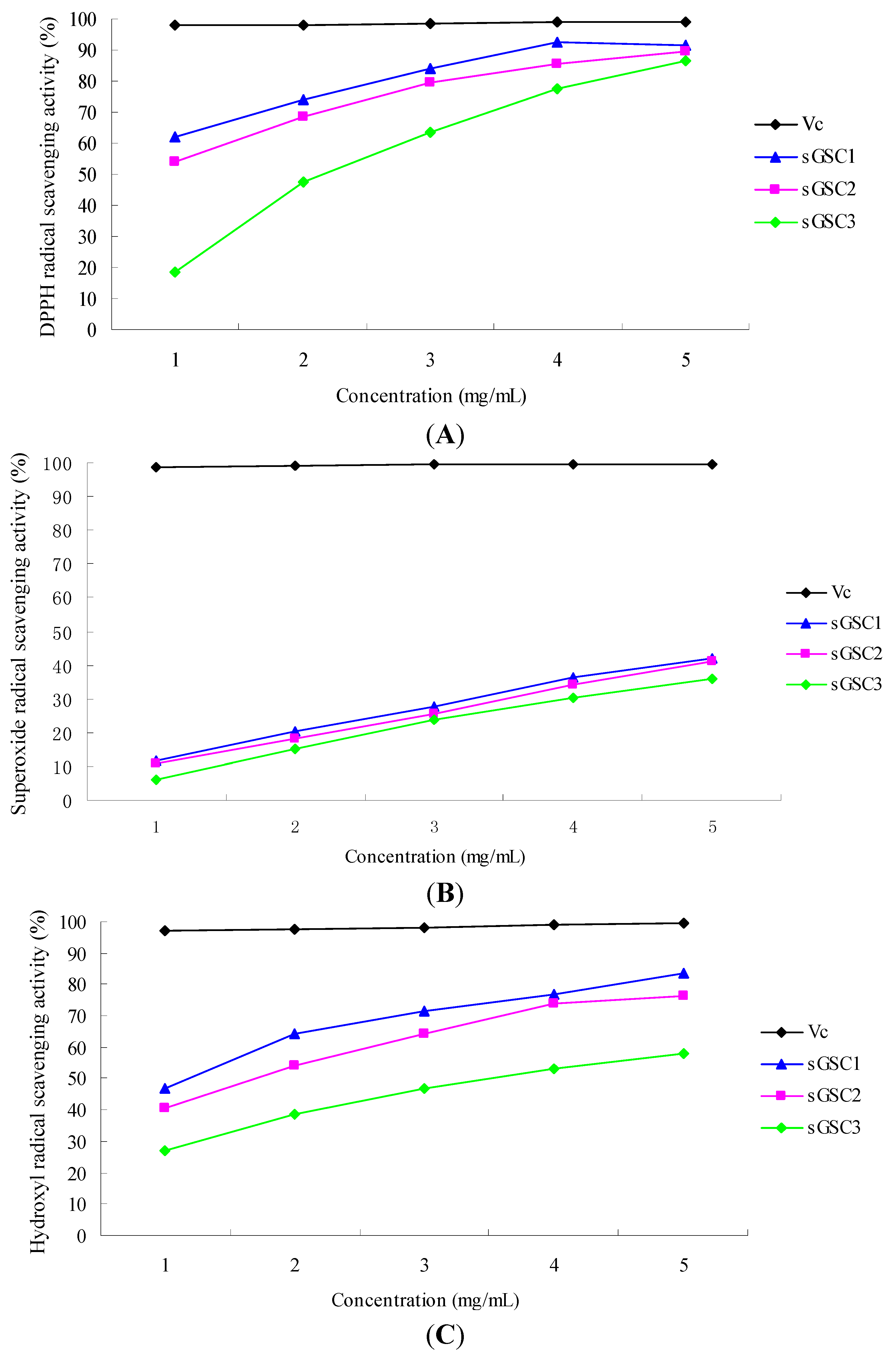
2.1.2. In Vitro Test
1,1-Diphenyl-2-picryl-hydrazyl Radical Scavenging Activity (DPPH RSA)

Superoxide RSA
Hydroxyl RSA
2.1.3. In Vivo Test
Effect of Glucan Administration on the Activities of Antioxidant Enzymes in Mouse Serum

{kind=link}
{kind=link}
| Group | SOD | CAT | GSH-PX | MDA |
|---|---|---|---|---|
| (U/mL) | (U/mL) | (μmol/L) | (nmol/mL) | |
| sGSC1 | 372.54 ± 9.18 a | 67.67 ± 1.53 c | 449.90 ± 3.61 b | 2.03 ± 0.09 b |
| sGSC2 | 371.34 ± 7.67 a | 56.33 ± 2.51 b | 413.20 ± 4.95 ab | 2.21 ± 0.04 b |
| sGSC3 | 368.66 ± 4.81 a | 41.85 ± 1.04 a | 378.35 ± 5.44 a | 2.38 ± 0.16 b |
| GSC | 371.57 ± 6.07 a | 42.00 ± 1.78 a | 384.48 ± 7.06 a | 5.84 ± 0.22 a |
| Control group | 368.88 ± 3.96 a | 37.86 ± 2.83 a | 357.30 ± 4.39 a | 6.27 ± 0.33 a |
Effect of Glucans on Spleen and Thymus Indexes in Mice
| Group | Spleen Index (%) | Thymus Index (%) |
|---|---|---|
| sGSC1 | 0.32 ± 0.03 c | 0.34 ± 0.03 c |
| sGSC2 | 0.30 ± 0.02 bc | 0.30 ± 0.02 bc |
| sGSC3 | 0.29 ± 0.03 bc | 0.30 ± 0.01 b |
| GSC | 0.28 ± 0.01 b | 0.28 ± 0.02 b |
| Control group | 0.22 ± 0.02 a | 0.24 ± 0.03 a |
Effect of Glucan Administration on Spleen Lymphocyte Proliferation in Mice
| Group | A570 Value |
|---|---|
| sGSC1 | 0.72 ± 0.02 c |
| sGSC2 | 0.69 ± 0.05 c |
| sGSC3 | 0.61 ± 0.04 b |
| GSC | 0.61 ± 0.05 b |
| Control group | 0.50 ± 0.04 a |
Effect of Glucan Administration on CD4+and CD8+ T Lymphocytes
| Group | CD4+ (%) | CD8+ (%) | CD4+/CD8+ |
|---|---|---|---|
| sGSC1 | 75.48 ± 0.71 d | 20.69 ± 0.37 c | 3.77 ± 0.75 b |
| sGSC2 | 73.35 ± 0.95 c | 21.52 ± 0.18 d | 3.41 ± 0.05 cd |
| sGSC3 | 71.92 ± 0.82 bc | 22.00 ± 0.13 bdc | 3.28 ± 0.20 d |
| GSC | 70.74 ± 0.78 ab | 23.62 ± 0.50 b | 3.00 ± 0.09 acd |
| Control group | 68.99 ± 0.52 a | 26.82 ± 0.34 a | 2.57 ± 0.10 a |
Effect of Glucan Administration on IL-2 and IFN-γ Secretion in Serum in Mice
| Group | IL-2 (pg/mL) | IFN-γ (pg/mL) |
|---|---|---|
| sGSC1 | 85.80 ± 3.56 d | 293.16 ± 6.72 bc |
| sGSC2 | 75.55 ± 2.21 c | 280.69 ± 4.45 cd |
| sGSC3 | 66.24 ± 3.31 b | 276.55 ± 7.27 bcd |
| GSC | 55.00 ± 4.12 a | 268.02 ± 5.96 ad |
| Control group | 54.40 ± 2.30 a | 249.80 ± 9.10 a |
2.2. Discussion
3. Materials and Methods
3.1. Reagents
3.2. Preparation of Glucan
3.3. Preparation of Sulfated Glucan
3.4. Determination of DS
3.5. Molecular Weight Analysis
3.6. FT-IR Spectroscopy
3.7. Antioxidant Activities in Vitro
3.7.1. DPPH RSA
3.7.2. Superoxide RSA
3.7.3. Hydroxyl RSA
3.8. Animal and Experimental Design
3.8.1. Relative Spleen and Thymus Weight
3.8.2. Biochemical Assay
3.8.3. Lymphocyte Proliferation Assay
3.8.4. Detection of CD4+ and CD8+ T Lymphocytes
3.8.5. Serum IL-2 and IFN-γ Concentration Assay
3.9. Statistical Analysis
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Zhu, F.M.; Du, B.; Bian, Z.X.; Xu, B.J. β-Glucans from edible and medicinal mushrooms: Characteristics, physicochemical and biological activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Rieder, A.; Samuelsen, A.B. Do cereal mixed-linked β-glucans possess immune-modulating activities? Mol. Nutr. Food Res. 2012, 56, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Jaehrig, S.C.; Rohn, S.; Kroh, L.W.; Fleischer, L.G.; Kurz, T. In vitro protential antioxidant activity of (1→3), (1→6)-β-d-glucan and protein fractions from Saccharomyces cerevisiae cell walls. J. Agric. Food Chem. 2007, 55, 4710–4716. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, S.D.V.; Cordeiro, S.L.; Cavalcanti, J.E.C.; Melchuna, K.M.; Lima, A.M.S.; Filho, I.A. Effects of purified Saccharomyces cerevisiae (1→3)-β-glucan on venous ulcer healing. Int. J. Mol. Sci. 2012, 13, 8142–8158. [Google Scholar] [CrossRef] [PubMed]
- Hofer, M.; Pospíšil, M. Modulation of animal and human hematopoiesis by β-glucans: A review. Molecules 2011, 16, 7969–7979. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Lin, C.Y.; Bian, Z.X.; Xu, B.J. An insight into anti-inflammatory effects of fungal β-glucan. Trends Food Sci. Technol. 2015, 41, 49–59. [Google Scholar] [CrossRef]
- Kim, H.J.; White, P.J. In vitro bile-acid binding and fermentation of high, medium, and low molecular weight β-glucan. J. Agric. Food Chem. 2010, 13, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; White, P.J. Impact of the molecular weight, viscosity, and solubility of β-glucan on in vitro oat starch digestibility. J. Agric. Food Chem. 2013, 61, 3270–3277. [Google Scholar] [CrossRef] [PubMed]
- Wolever, T.M.S.; Tosh, S.M.; Gibbs, A.L.; Brand-Miller, J.; Duncan, A.M.; Hart, V.; Lamarche, B.; Thomson, B.A.; Duss, R.; Wood, P.J. Physicochemical properties of oat β-glucan influence its ability to reduce serum LDL cholesterol in humans. A randomized clinical trial. Am. J. Clin. Nutr. 2010, 92, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Rop, O.; Mlcek, J.; Jurikova, T. β-Glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.N.; Queenan, K.M.; Thomas, W.; Fulcher, R.G.; Slavin, J.L. Physiological effects of concentrated barley β-glucan in mildly hypercho-lesterolemic adults. J. Am. Coll. Nutr. 2008, 27, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Thondre, P.S.; Shafat, A.; Clegg, M.E. Molecular weight of barley β-glucan influences energy expenditure, gastric emptying and glycemic response in human subjects. Br. J. Nutr. 2013, 110, 2173–2179. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, K.; Wilczak, J.; Harasym, J.; Gudej, S.; Suchecka, D.; Krolikowski, T.; Lange, E.; Gromadzka-Ostrowska, J. Impact of low and high molecular weight oat β-glucan on oxidative stress and antioxidant defense in spleen of rats with LPS induced enteritis. Food Hydrocoll. 2015, 51, 272–280. [Google Scholar] [CrossRef]
- Choromanska, A.; Kulbacka, J.; Rembialkowska, N.; Pilat, J.; Oledzki, R.; Harasym, J.; Saczko, J. Anticancer properties of low molecular weight oat β-glucan—An in vitro study. Int. J. Biol. Macromol. 2015, 80, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Methacanon, P.; Weerawatsophon, U.; Tanjak, P.; Rachtawee, P.; Prathumpai, W. Interleukin-8 stimulating activity of low molecular weight β-glucan depolymerized by γ-irradiation. Carbohydr. Polym. 2011, 86, 574–580. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.Y.; Pang, X.B.; Yao, W.B.; Gao, X.D. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 46, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cheung, P.C.K.; Zhang, L. Evaluation of mushroom dietary fiber (nonstarch polysaccharides) from sclerotia of Pleurotus tuber-regium (Fries) singer as a potential antitumor agent. J. Agric. Food Chem. 2001, 49, 5059–5062. [Google Scholar] [CrossRef] [PubMed]
- Zeković, D.B.; Wiatkowski, S.K.; Vrvić, M.M.; Jakovljević, D.; Moran, C.A. Natural and modified (1→3)-β-d-glucans in health promotion and disease alleviation. Crit. Rev. Biotechnol. 2005, 25, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L.; Pretus, H.A.; McNamee, R.B.; Jones, E.L.; Ensley, H.E.; Browder, I.W. Development of a water-soluble, sulfated (1→3)-β-d-glucan biological response modifier derived from Saccharomyces cerevisiae. Carbohydr. Res. 1992, 235, 247–257. [Google Scholar] [CrossRef]
- Wang, M.; Yang, R.L.; Zhang, L.F.; Meng, X.Y.; Fei, C.Z.; Zhang, K.Y.; Wang, X.Y.; Zheng, W.L.; Xiao, S.; Zhang, S.Q.; et al. Sulfated glucan can improve the immune efficacy of Newcastle disease vaccine in chicken. Int. J. Biol. Macromol. 2014, 59, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Niu, S.F.; Zhao, B.T.; Wang, X.F.; Yao, J.; Zhang, J.; Zhao, W.W.; Zhao, Y.T. Regioselective synthesis of sulfated guar gum: Comparative studies of structure and antioxidant activities. Int. J. Biol. Macromol. 2013, 62, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Du, Y.M.; Li, Y.; Yang, J.H.; Feng, T.; Zhang, L.; Kennedy, J.F. Preparation of lacquer polysaccharide sulfates and their antioxidant activity in vitro. Carbohydr. Polym. 2008, 73, 322–331. [Google Scholar] [CrossRef]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Xing, R.; Liu, S.; Yu, H.H.; Guo, Z.Y.; Li, Z.E.; Li, P.C. Preparation of high-molecular weight and high-sulfate content chitosans and their potential antioxidant activity in vitro. Carbohydr. Polym. 2005, 61, 148–154. [Google Scholar] [CrossRef]
- Wang, J.L.; Niu, S.F.; Zhao, B.T.; Luo, T.; Liu, D.; Zhang, J. Catalytic synthesis of sulfated polysaccharides II: Comparative studies of solution conformation and antioxidant activities. Carbohydr. Polym. 2014, 107, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Stout, A.K.; Raphael, H.M.; Kanterewicz, B.I.; Klann, E.; Reynolds, I.J. Glutamate-induced neuron death requires mitochondrial calcium uptake. Nat. Neurosci. 1998, 1, 366–373. [Google Scholar] [PubMed]
- White, E.; Shannon, J.S.; Patterson, R.E. Relationship between vitamin and calcium supplement use and colon cancer. Cancer Epidemiol. Biomark. Prev. 1997, 6, 769–774. [Google Scholar]
- Smith, A.R.; Shenvi, S.V.; Widlansky, M.; Suh, J.H.; Hagen, T.M. Lipoic acid as a potential therapy for chronic diseases associated with oxidative stress. Curr. Med. Chem. 2004, 11, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Sharoni, Y.; Danilenko, M.; Dubi, N.; Ben-Dor, A.; Levy, J. Carotenoids and transcription. Arch. Biochem. Biophys. 2004, 430, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.L.; Qiao, D.L.; Gan, D.; Sun, Y.; Ye, H.; Zeng, X.X. Antioxidant acitivity in vitro and in vivo of the capsule polysaccharides from Streptococcus equi subsp. zooepidemicus. Carbohydr. Polym. 2009, 75, 677–682. [Google Scholar] [CrossRef]
- Ping, C.X.; Yan, C.; Li, S.B.; Chen, Y.G.; Lan, J.Y.; Liu, L.P. Free radical scavenging of Ganoderma lucidum polysaccharides and its effect on antioxidant enzymes and immunity activities in cervical carcinoma rats. Carbohydr. Polym. 2009, 77, 389–393. [Google Scholar]
- Chen, Y.G.; Shen, Z.J.; Chen, X.P. Modulatory effect of Ganoderma lucidum polysaccharides on serum antioxidant enzymes activities in ovarian cancer rats. Carbohydr. Polym. 2009, 78, 258–262. [Google Scholar]
- Chi, A.P.; Kang, C.Z.; Zhang, Y.; Tang, L.; Guo, H.H.; Li, H.; Zhang, K.R. Immunomodulating and antioxidant effects of polysaccharide conjugates from the fruits of Ziziphus Jujube on Chronic Fatigue Syndrome rats. Carbohydr. Polym. 2015, 122, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.H.; Yang, X.L.; Wang, Y.; Xu, H.B. Effect of degree of substitution on the antioxidative activities of the sulfated Hunai polysaccharide. J. Huazhong Univ. Sci. Technol. 2000, 28, 104–107. [Google Scholar]
- Wang, M.; Meng, X.Y.; Yang, R.L.; Qin, T.; Wang, X.Y.; Zhang, K.Y.; Fei, C.Z.; Li, Y.; Hu, Y.L.; Xue, F.Q. Cordyceps militaris polysaccharides can improve the immune efficacy of Newcastle disease vaccine in chicken. Int. J. Biol. Macromol. 2013, 59, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, Y.; Wang, D.; Liu, J.; Zhang, J.; Abula, S.; Zhao, B.; Ruan, S. Sulfated modification can enhance the immune-enhancing activity of Lycium barbarum polysaccharides. Cell. Immunol. 2010, 263, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Mullen, A.C.; High, F.A.; Hutchins, A.S.; Lee, H.W.; Villarino, A.V. Role of T-bet in commitment of TH1 cells before IL-12 dependent selection. Science 2001, 292, 1907–1910. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Ohta, A. A critical role for antigen-specific Th1 cells in acute liver injury in mice. J. Immunol. 1999, 162, 6503–6509. [Google Scholar] [PubMed]
- Zhang, N.; Bevan, M.J. CD8+ T cells: Foot soldiers of the immune systems. Immunity 2011, 35, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.F.; Wang, C.D.; Bu, Y.Q.; Xiang, T.X.; Huang, X.N.; Wang, Z.W.; Yi, F.P.; Ren, G.S.; Liu, G.L.; Song, F.Z. Antioxidative and immunoprotective effects of Pyracantha fortuneana (Maxim.) Li polysaccharides in mice. Immunol. Lett. 2010, 133, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Li, J.; Hu, Y. Polysaccharides from Inonotus obliquus sclerotia and cultured mycelia stimulate cytokine production of human peripheral blood mononuclear cells in vitro and their chemical characterization. Int. Immunopharmacol. 2014, 21, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.J.; Wang, S.; Xia, Y.J.; Tu, M.W.; Zhang, L.J.; Wang, Y.M. Antitumor effects and immune regulation activities of a purified polysaccharide extracted from Juglan regia. Int. J. Biol. Macromol. 2015, 72, 771–775. [Google Scholar]
- Ma, X.; Guo, Z.H.; Wang, D.Y.; Hu, Y.L.; Shen, Z.Q. Effects of sulfated polysaccharides and their prescriptions on immune response of ND vaccine in chicken. Carbohydr. Polym. 2010, 82, 9–13. [Google Scholar] [CrossRef]
- Saha, S.K.; Brewer, C.F. Determination of the concentrations of oligosaccharides, complex type carbonyhdrates, and glycoproteins using the phenol-sulfuric acid method. Carbohydr. Res. 1994, 254, 157–167. [Google Scholar] [CrossRef]
- Tsiapali, E.; Whaley, S.; Kalbfleisch, J.; Ensley, H.E.; Browder, I.W.; Williams, D.L. Glucans exhibit weak antioxidant activity, but stimulate macrophage free radical activity. Free Radic. Biol. Med. 2001, 30, 393–402. [Google Scholar] [CrossRef]
- Li, S.; Shah, N.P. Antioxidant and antibacterial activities of sulphated polysaccharides from Pleurotus eryngii and Streptococcus thermophilus ASCC 1275. Food Chem. 2014, 165, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, Y.; Wang, W.D. Optimization of ultrasonic-assisted extraction and in vitro antioxidant activities of polysaccharides from Trametes orientalis. Carbohydr. Polym. 2014, 111, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Robak, J.; Gryglewski, R.J. Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 1988, 37, 837–841. [Google Scholar] [CrossRef]
- Wang, M.; Meng, X.Y.; Yang, R.L.; Qin, T.; Wang, X.Y.; Zhang, K.Y.; Fei, C.Z.; Li, Y.; Hu, Y.L.; Xue, F.Q. Cordyceps militaris polysaccharides can enhance the immunity and antioxidation activity in immunosuppressed mice. Carbohydr. Polym. 2012, 89, 461–466. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, N.; Wang, M.; Zhang, L.; Xiao, S.; Fei, C.; Wang, X.; Zhang, K.; Zheng, W.; Wang, C.; Yang, R.; et al. Effects of Low Molecular Weight Yeast β-Glucan on Antioxidant and Immunological Activities in Mice. Int. J. Mol. Sci. 2015, 16, 21575-21590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160921575
Lei N, Wang M, Zhang L, Xiao S, Fei C, Wang X, Zhang K, Zheng W, Wang C, Yang R, et al. Effects of Low Molecular Weight Yeast β-Glucan on Antioxidant and Immunological Activities in Mice. International Journal of Molecular Sciences. 2015; 16(9):21575-21590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160921575
Chicago/Turabian StyleLei, Na, Mi Wang, Lifang Zhang, Sui Xiao, Chengzhong Fei, Xiaoyang Wang, Keyu Zhang, Wenli Zheng, Chunmei Wang, Ruile Yang, and et al. 2015. "Effects of Low Molecular Weight Yeast β-Glucan on Antioxidant and Immunological Activities in Mice" International Journal of Molecular Sciences 16, no. 9: 21575-21590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160921575