
Chemically Bonding of Amantadine with Gardenamide A Enhances the Neuroprotective Effects against Corticosterone-Induced Insults in PC12 Cells

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
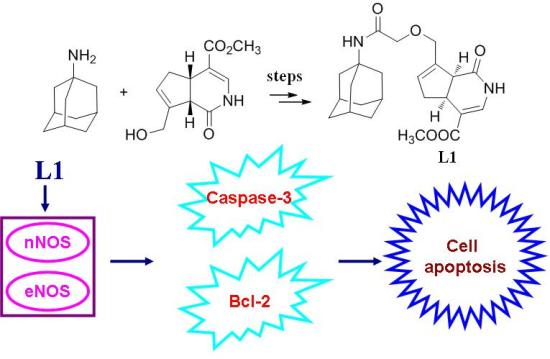
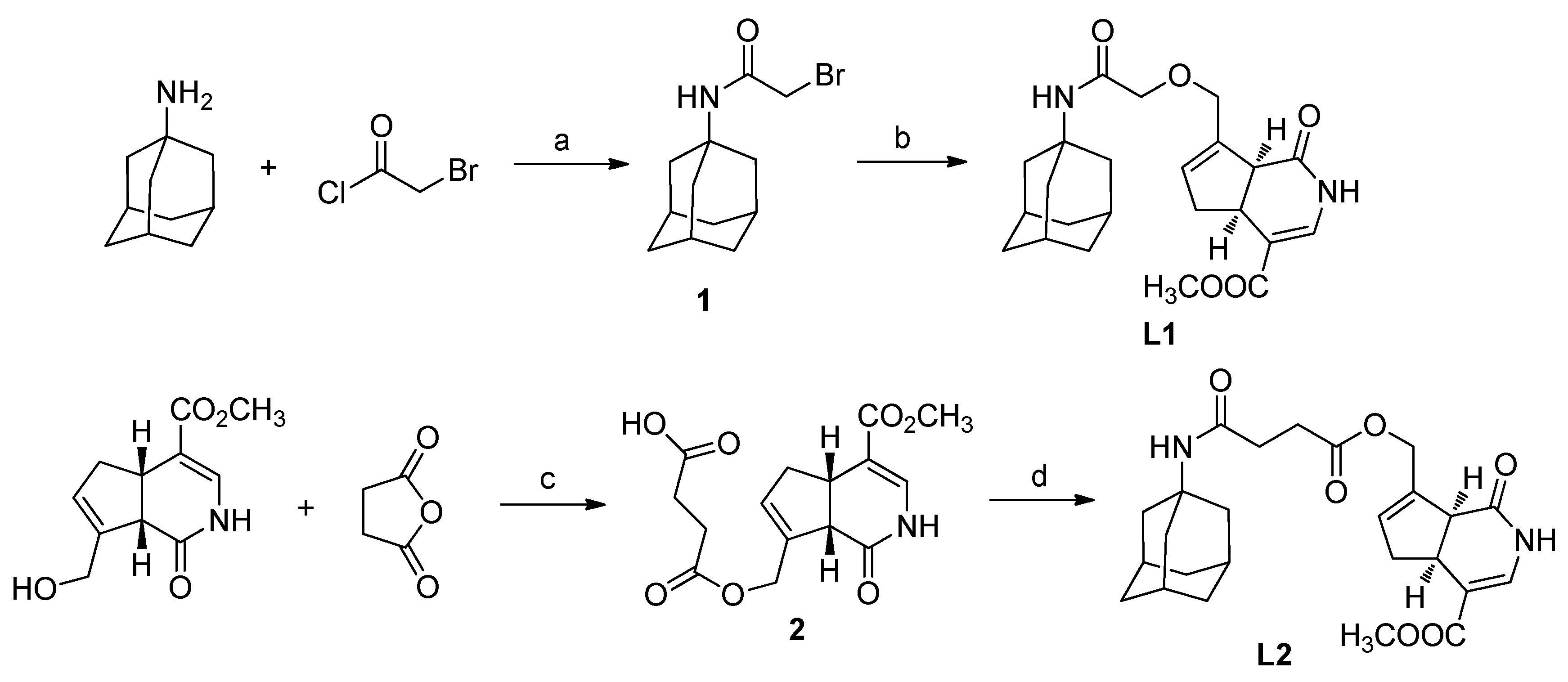
2.1. Chemistry

2.2. L1 Showed the Most Active Neuroprotective Effect

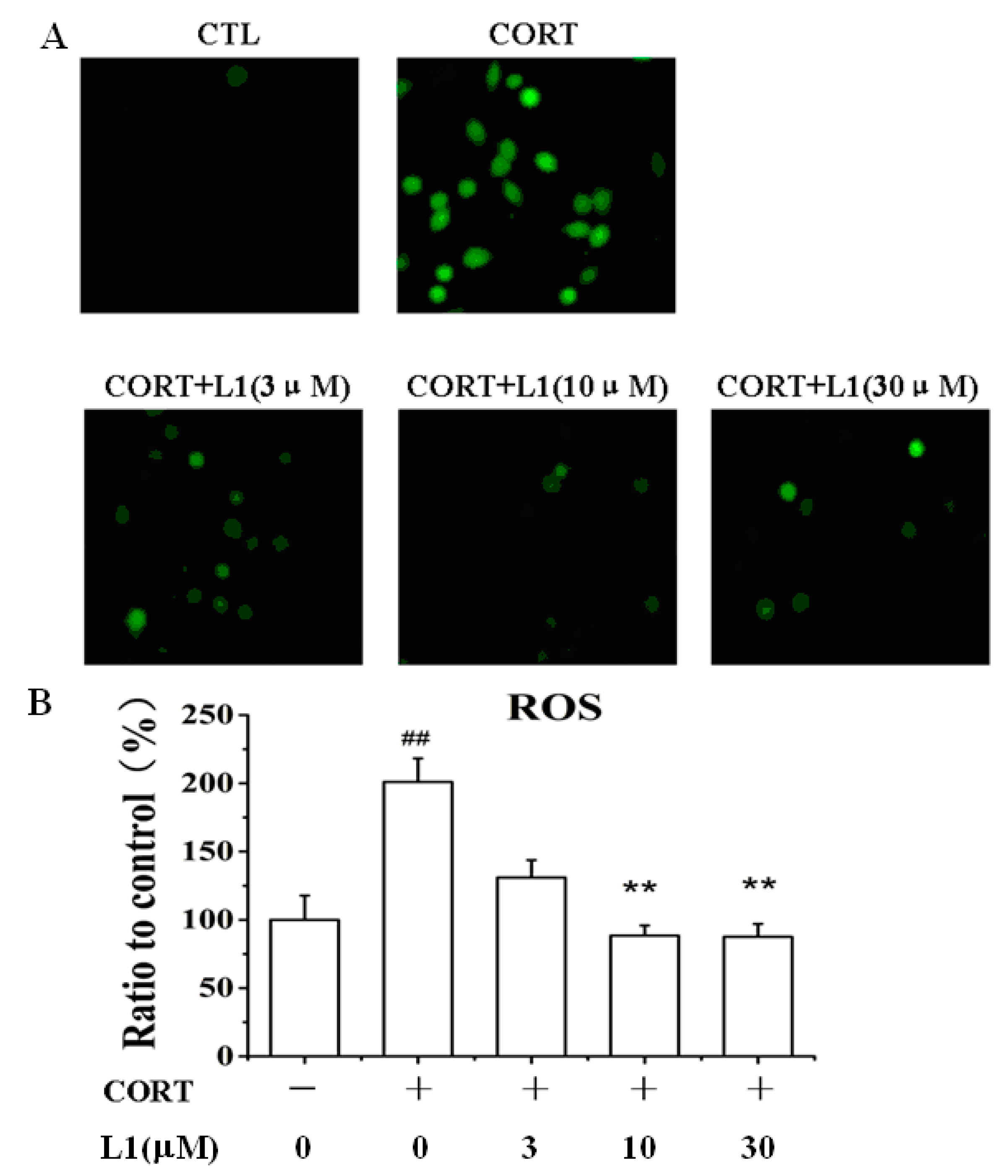
2.3. L1 Dose-Dependently Attenuated the Increase of ROS Caused by CORT

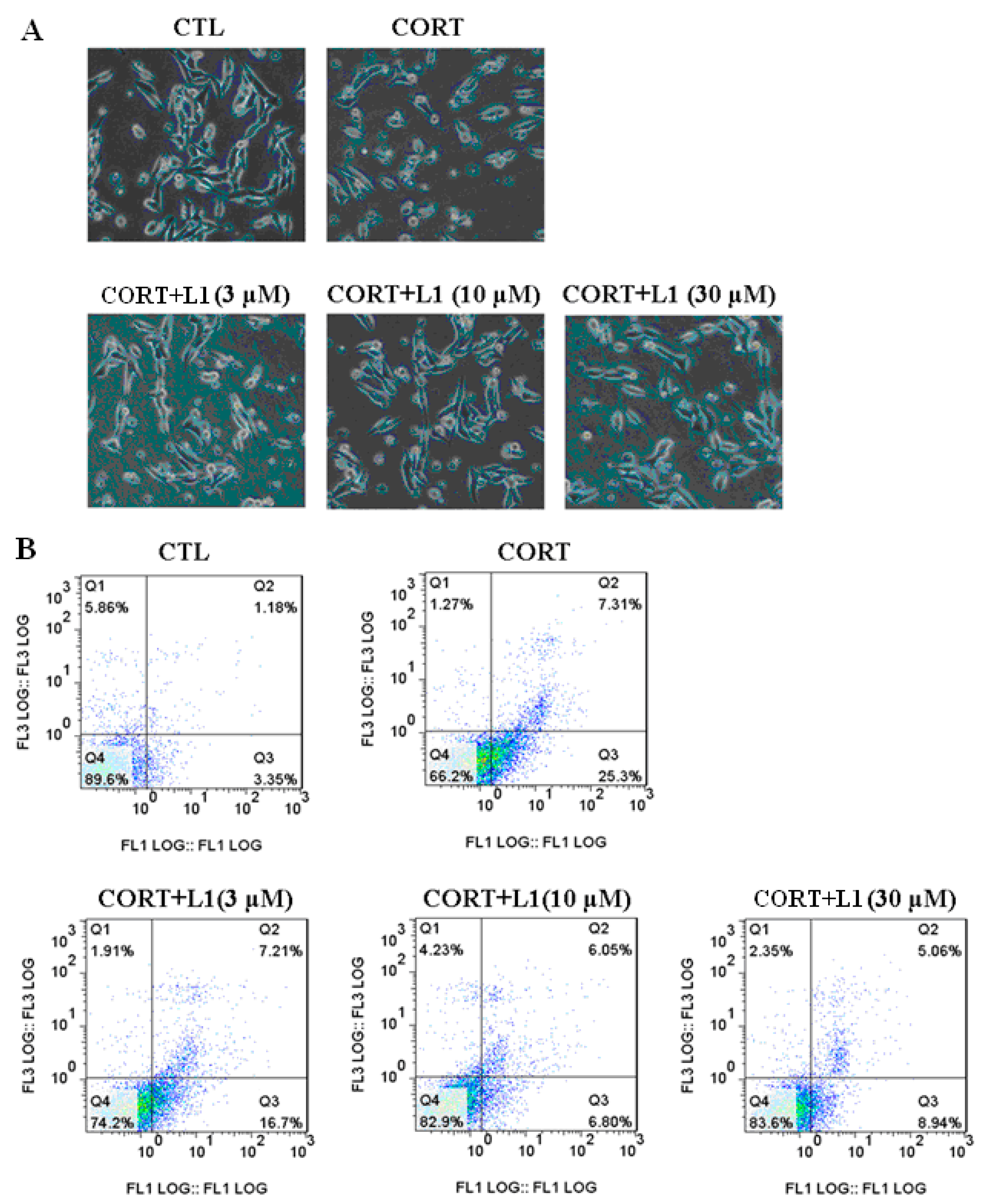
2.4. L1 Reduced CORT-Induced Cell Morphological Change and Apoptosis

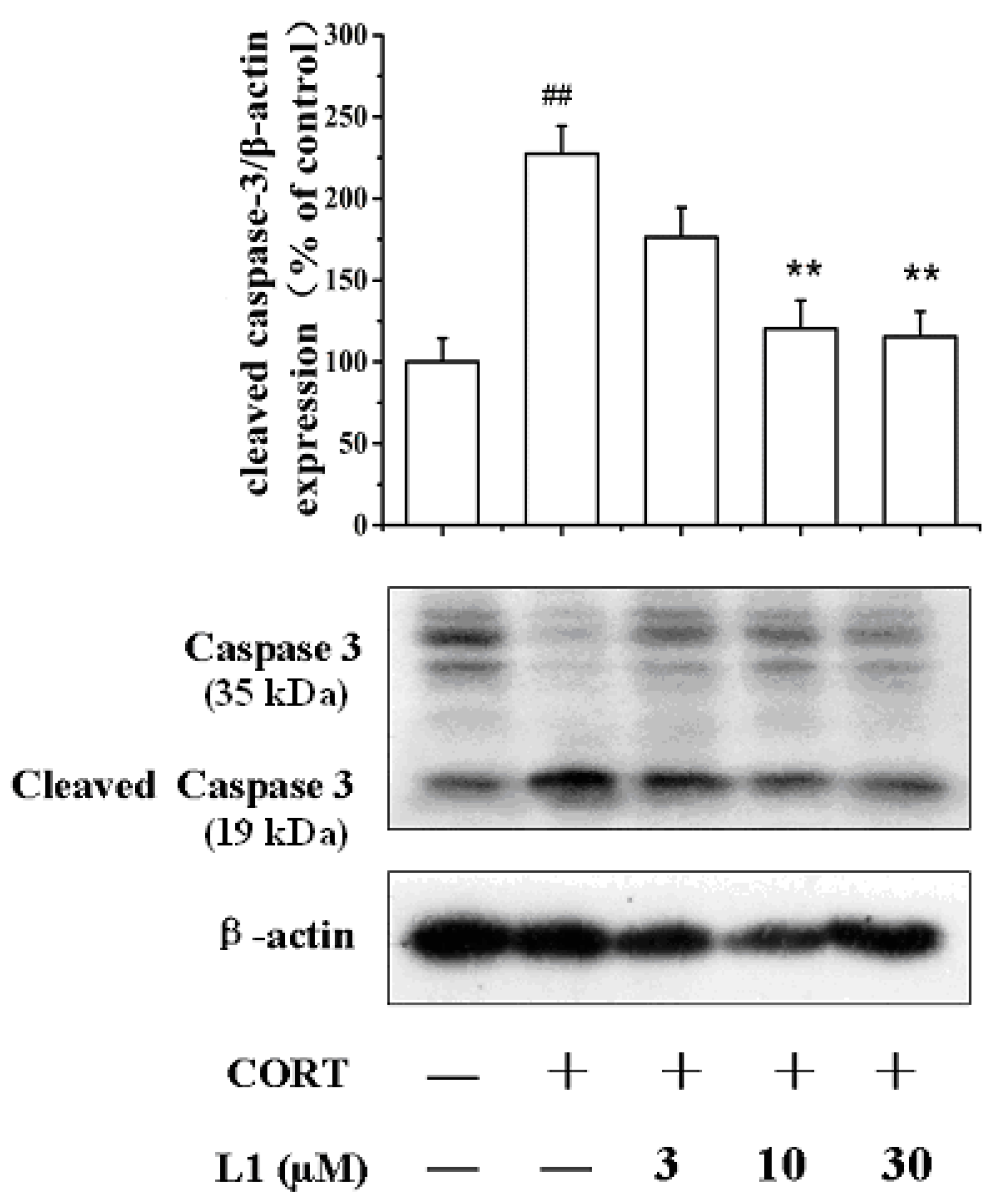
2.5. L1 Increased Caspase-3 (35 kDa) Level and Decreased Cleaved Caspase-3 (19 kDa) Level

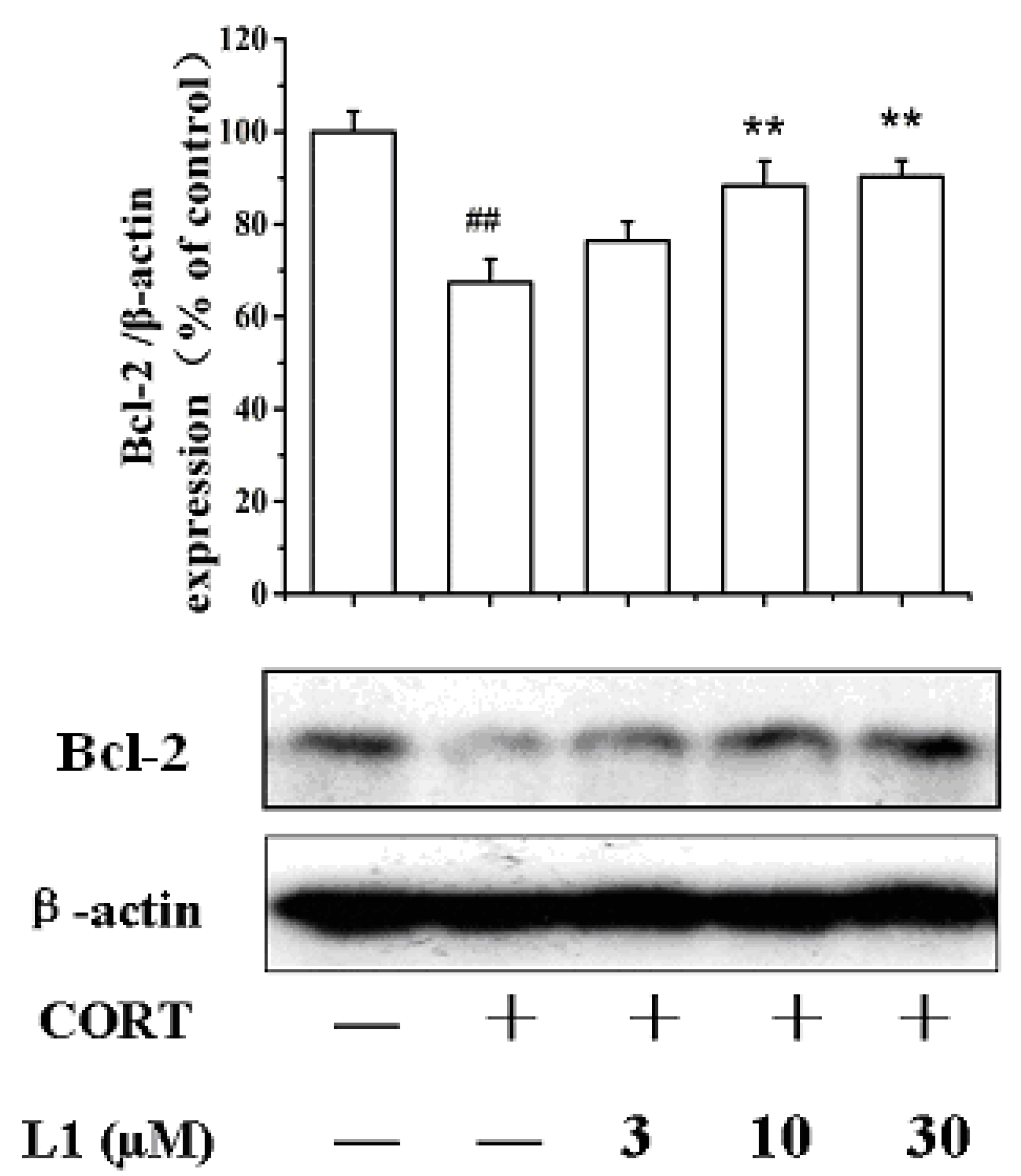
2.6. L1 Restored CORT-Inhibited Bcl-2 Expression

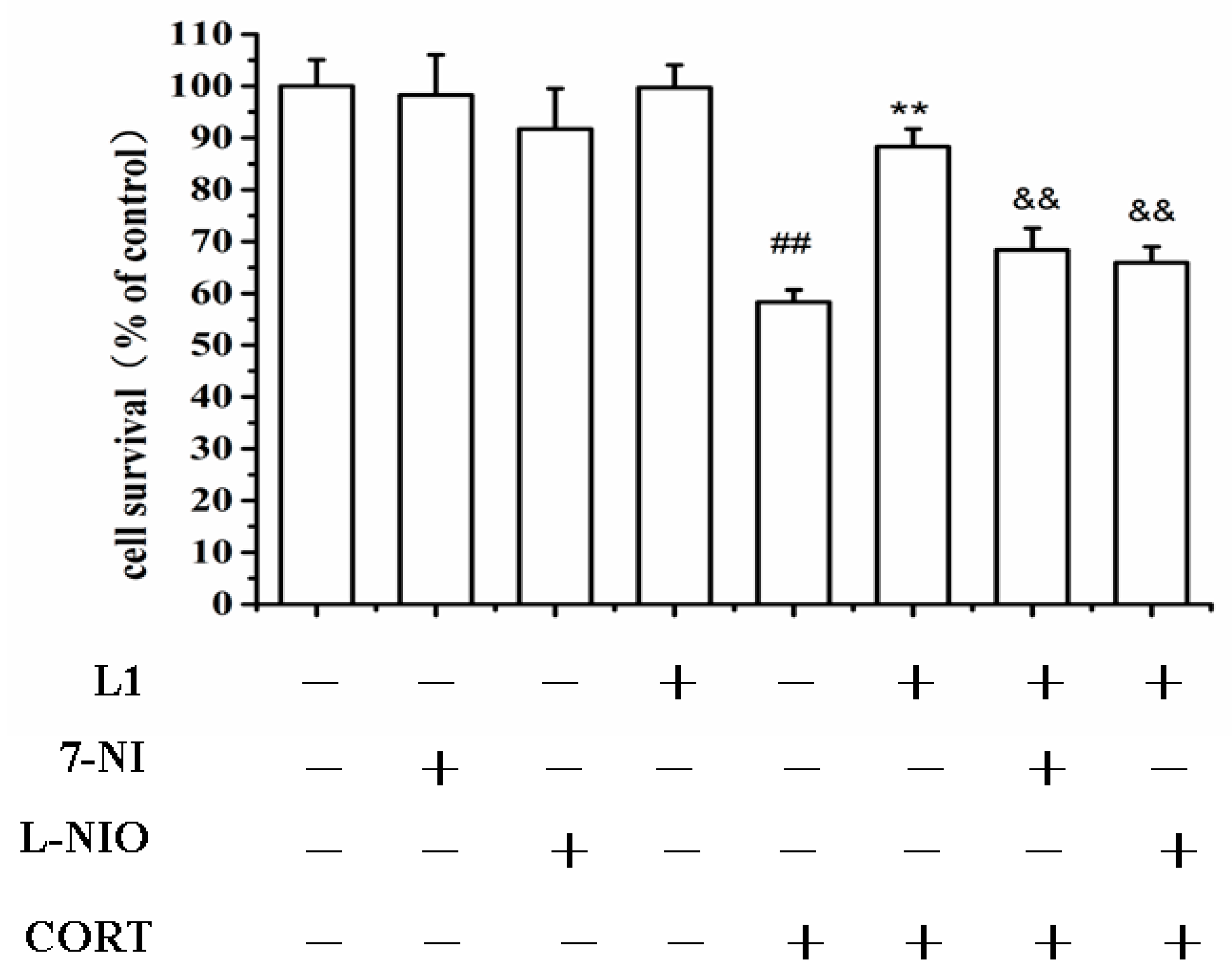
2.7. nNOS and eNOS might Involve in the Neuroprotection of L1

3. Experimental Section
3.1. Materials and Methods
3.2. Synthesis of N-α-Bromoacetyl Amantadine (1)
3.3. Synthesis of O-(2-(Adamantan-1-ylamino)-2-oxoethyl)gardenamide A (L1)
3.4. Synthesis of O-β-Carboxylpropionyl Gardenamide A (2)
3.5. Synthesis of O-(4-(Adamantan-1-ylamino)succinyl Gardenamide A (L2)
3.6. Culture of PC12 Cells
3.7. Neuroprotective Activity Tests
3.8. MTT Assay
3.9. Neuroprotective Inhibition Test
3.10. Measurement of Intracellular ROS
3.11. Morphologic Changes
3.12. Flow Cytometry Analysis for Cell Apoptosis
3.13. Western Blot
3.14. Data Analysis and Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Machida, K.; Oyama, K.; Ishii, M.; Kakuda, R.; Yaoita, Y.; Kikuchi, M. Studies of the constituents of Gardenia species. II. Terpenoids from Gardeniae Fructus. Chem. Pharm. Bull. 2000, 48, 746–748. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, G.; Hamm, A.; Kraus, J.; Ochse, M.; Noureldeen, A.; Jumbam, D. Gardenamide A from Rothmannia. urcelliformis (Rubiaceae.)-Isolation, Absolute Stereostructure, and Biomimetic Synthesis from Genipine. Eur. J. Org. Chem. 2001, 2001, 1983–1987. [Google Scholar] [CrossRef]
- Luo, J.; Wang, R.; Huang, Z.; Yang, J.; Yao, X.; Chen, H.; Zheng, W. Synthesis of stable genipin derivatives and studies of their neuroprotective activity in PC12 cells. ChemMedChem 2012, 7, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Talmi, M.; Carlier, E.; Soumireu-Mourat, B. Similar effects of aging and coricosterone treatment on mouse hippocampus function. Neurobiol. Aging 1993, 14, 239–244. [Google Scholar] [CrossRef]
- Hortnagl, H.; Berger, M.L.; Havelec, L.; Hornykiewicz, O. Role of glucocorticoids in the cholinergic degeneration in rat hippocampus induced by ethylcholine aziridinium (AF64A). J. Neurosci. 1993, 13, 2939–2945. [Google Scholar] [PubMed]
- Lussier, A.L.; Caruncho, H.J.; Kalynchuk, L.E. Repeated exposure to corticosterone, but not restraint, decreases the number of reelin-positive cells in the adult rat hippocampus. Neurosci. Lett. 2009, 460, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Lussier, A.L.; Lebedeva, K.; Fenton, E.Y.; Guskjolen, A.; Caruncho, H.J.; Kalynchuk, L.E. The progressive development of depression-like behavior in corticosterone-treated rats is paralleled by slowed granule cell maturation and decreased reelin expression in the adult dentate gyrus. Neuropharmacology 2013, 72, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.G.; Zhu, L.J.; Chen, C.; Wu, H.Y.; Luo, C.X.; Chang, L.; Zhu, D.Y. Hippocampal neuronal nitric oxide Synthase Mediates the Stress-Related Depressive Behaviors of glucocorticoids by downregulating glucocorticoid receptor. J. Neurosci. 2011, 31, 7579–7590. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.G.; Hu, Y.; Hua, Y.; Hu, M.; Luo, C.X.; Han, X.; Zhu, X.J.; Wang, B.; Xu, J.S.; Zhu, D.Y. Neuronal nitric oxide synthase contributes to chronic stress-induced depression by suppressing hippocampal neurogenesis. J. Neurochem. 2007, 103, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.M.; Flubacher, A.; Tinnes, S.; Heyer, A.; Scheller, M.; Herpfer, I.; Berger, M.; Frotscher, M.; Lieb, K.; Haas, C.A. Early life stress stimulates hippocampal reelin gene expression in a sex-specific manner: Evidence for corticosterone-mediated action. Hippocampus 2012, 22, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Blanpied, T.A.; Clarke, R.J.; Johnson, J.W. Amantadine inhibits NMDA receptors by accelerating channel closure during channel block. J. Neurosci. 2005, 25, 3312–3322. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, P.J.; Metman, L.V.; Chase, T.N. Renaissance of amantadine in the treatment of Parkinson’s disease. Adv. Neurol. 2003, 91, 251–257. [Google Scholar] [PubMed]
- Dingledine, R.; Borges, K.; Bowie, D.; Traynelis, S.F. The glutamate receptor ion channels. Pharmacol. Rev. 1999, 51, 7–61. [Google Scholar] [PubMed]
- Zhou, J.-Z.; Zheng, J.-Q.; Zhang, Y.-X.; Zhou, J.-H. Corticosterone impairs cultured hippocampal neurons and facilitates Ca2+ influx through voltage-dependent Ca2+ channel. Acta Pharmacol. Sin. 2000, 21, 156–160. [Google Scholar] [PubMed]
- Leonard, B.; Maes, M. Mechanistic explanations how cell-mediated immune activation, inflammation and oxidative and nitrosative stress pathways and their sequels and concomitants play a role in the pathophysiology of unipolar depression. Neurosci. Biobehav. Rev. 2012, 36, 764–785. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Yirmyia, R.; Noraberg, J.; Brene, S.; Hibbeln, J.; Perini, G.; Kubera, M.; Bob, P.; Lerer, B.; Maj, M. The inflammatory & neurodegenerative (I & ND) hypothesis of depression: Leads for future research and new drug developments in depression. Metab. Brain Dis. 2009, 24, 27–53. [Google Scholar] [PubMed]
- Wang, R.; Yang, J.; Peng, L.; Zhao, J.; Mu, N.; Huang, J.; Lazarovici, P.; Chen, H.; Zheng, W. Gardenamide A attenuated cell apoptosis induced by serum deprivation via ERK1/2 and PI3K/Akt signaling pathways. Neuroscience 2014, 286, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.W.; Machado-Vieira, R.; Pavlatou, M.G. Clinical and biochemical manifestations of depression: relation to the neurobiology of stress. Neural Plast. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Ge, H.; Tang, J.; Fu, C.; Duanmu, W.; Chen, Y.; Hu, R.; Sui, J.; Liu, X.; Feng, H. Amantadine preserves dopamine level and attenuates depression-like behavior induced by traumatic brain injury in rats. Behav. Brain Res. 2015, 279, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y.; Finger, L.R.; Yunis, J.; Nowell, P.C.; Croce, C.M. The Bcl-2 protein family: Arbiters of cell survival. Science 1984, 226, 1097–1099. [Google Scholar] [CrossRef] [PubMed]
- Cleary, M.L.; Smith, S.D.; Sklar, J. Cloning and structural analysis of cDNAs for bcl-2 and a hybrid bcl-2/immunoglobulin transcript resulting from the t(14;18) translocation. Cell 1986, 47, 19–28. [Google Scholar] [CrossRef]
- Oliveira, R.M.; Guimarães, F.S.; Deakin, J.F. Expression of neuronal nitric oxide synthase in the hippocampal formation in affective disorders. Braz. J. Med. Biol. Res. 2008, 41, 333–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildiz, F.; Erden, B.F.; Ulak, G.; Utkan, T.; Gacar, N. Antidepressant-like effect of 7-nitroindazole in the forced swimming test in rats. Psychopharmacology 2000, 149, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Spiacci, A., Jr.; Kanamaru, F.; Guimarães, F.S.; Oliveira, R.M. Nitric oxide-mediated anxiolytic-like and antidepressant-like effects in animal models of anxiety and depression. Pharmacol. Biochem. Behav. 2008, 88, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.R.; Oliveira, A.M.; Dinarte, A.R.; Pinheiro, D.G.; Greene, L.J.; Silva, W.A., Jr.; Joca, S.R.; Guimarães, F.S. Changes in hippocampal gene expression by 7-nitroindazole in rats submitted to forced swimming stress. Genes Brain Behav. 2012, 11, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Ochi, S.; Yamazaki, K.; Nakata, S.; Abe, M.; Mori, Y.; Ueno, S. Antidepressant action via the nitric oxide system: A pilot study in an acute depressive model induced by arginin. Neurosci. Lett. 2015, 599, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.H.; Quirion, R. Glutamate acting on N-methyl-d-aspartate receptors attenuates insulin-like growth factor-1 receptor tyrosine phosphorylation and its survival signaling properties in rat hippocampal neurons. J. Biol. Chem. 2009, 284, 855–861. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Peng, L.; Zheng, W.; Wang, R.; Zhang, L.; Yang, J.; Chen, H. Chemically Bonding of Amantadine with Gardenamide A Enhances the Neuroprotective Effects against Corticosterone-Induced Insults in PC12 Cells. Int. J. Mol. Sci. 2015, 16, 22795-22810. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160922795
Zhao J, Peng L, Zheng W, Wang R, Zhang L, Yang J, Chen H. Chemically Bonding of Amantadine with Gardenamide A Enhances the Neuroprotective Effects against Corticosterone-Induced Insults in PC12 Cells. International Journal of Molecular Sciences. 2015; 16(9):22795-22810. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160922795
Chicago/Turabian StyleZhao, Jiaqiang, Lizhi Peng, Wenhua Zheng, Rikang Wang, Lei Zhang, Jian Yang, and Heru Chen. 2015. "Chemically Bonding of Amantadine with Gardenamide A Enhances the Neuroprotective Effects against Corticosterone-Induced Insults in PC12 Cells" International Journal of Molecular Sciences 16, no. 9: 22795-22810. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160922795