
Platelet-Rich Plasma Increases the Levels of Catabolic Molecules and Cellular Dedifferentiation in the Meniscus of a Rabbit Model

 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. In Vitro
2.1.1. Assessment of the Prepared PRP
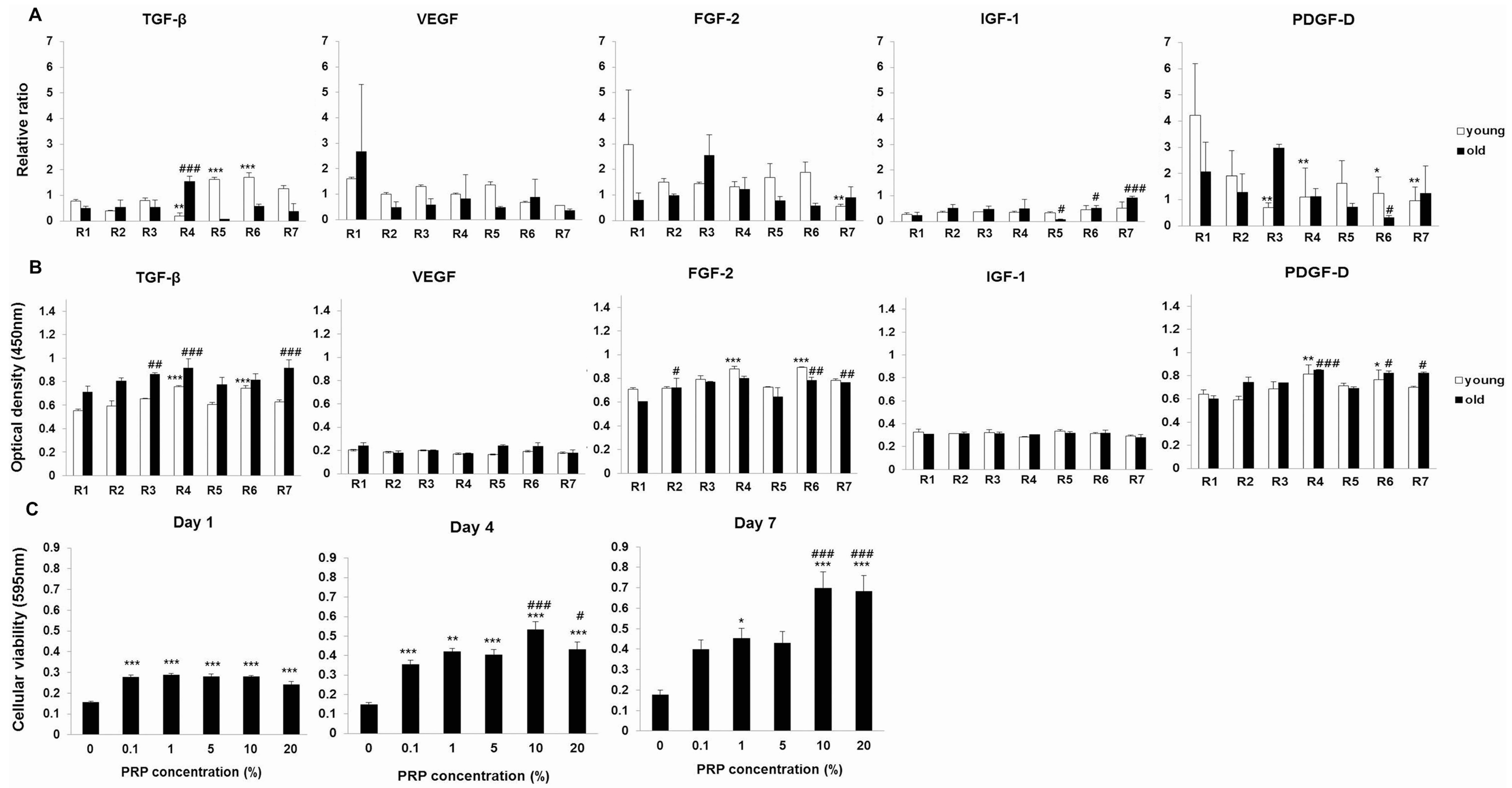
2.1.2. Effects of PRP on Meniscal Cell Viability
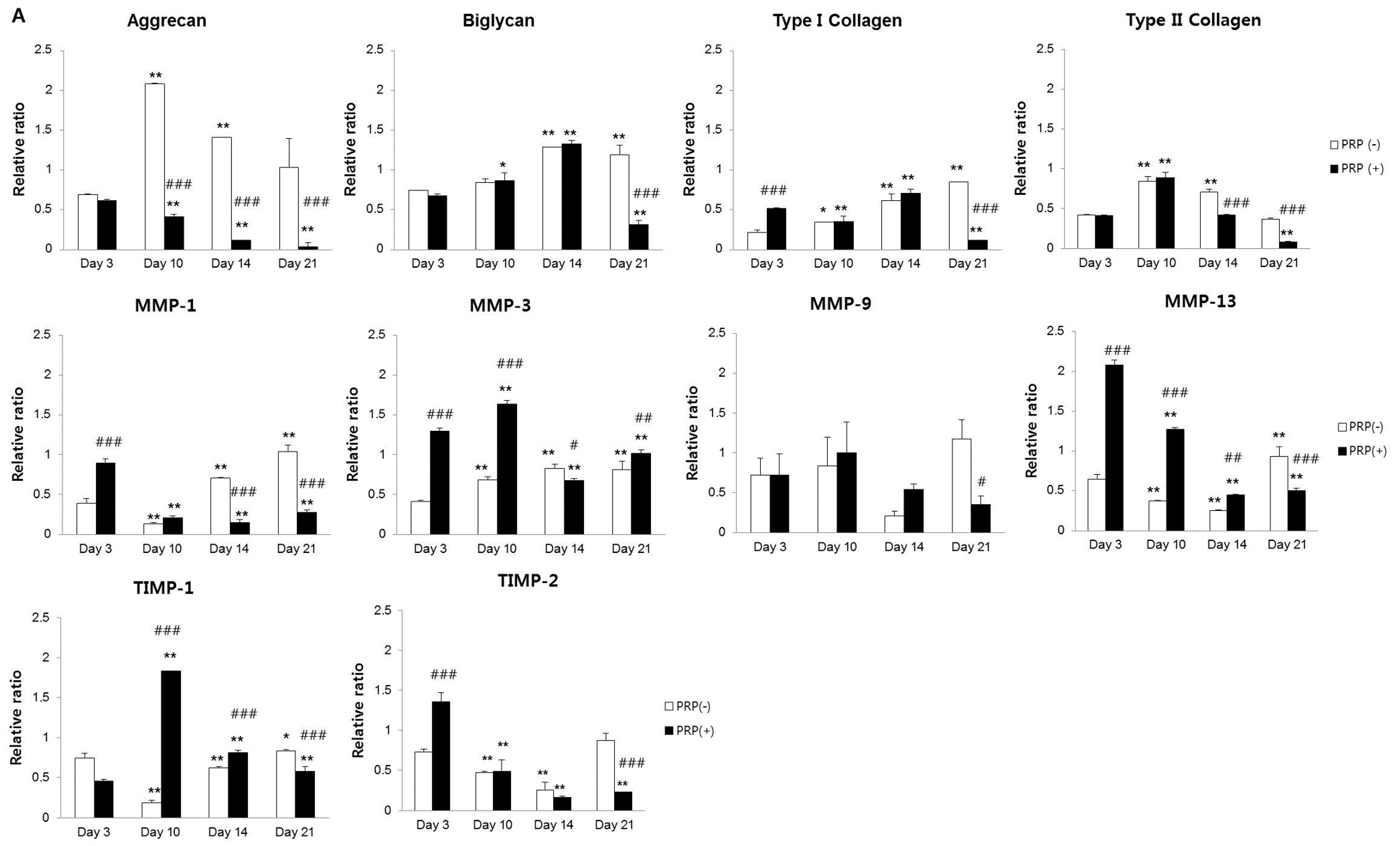
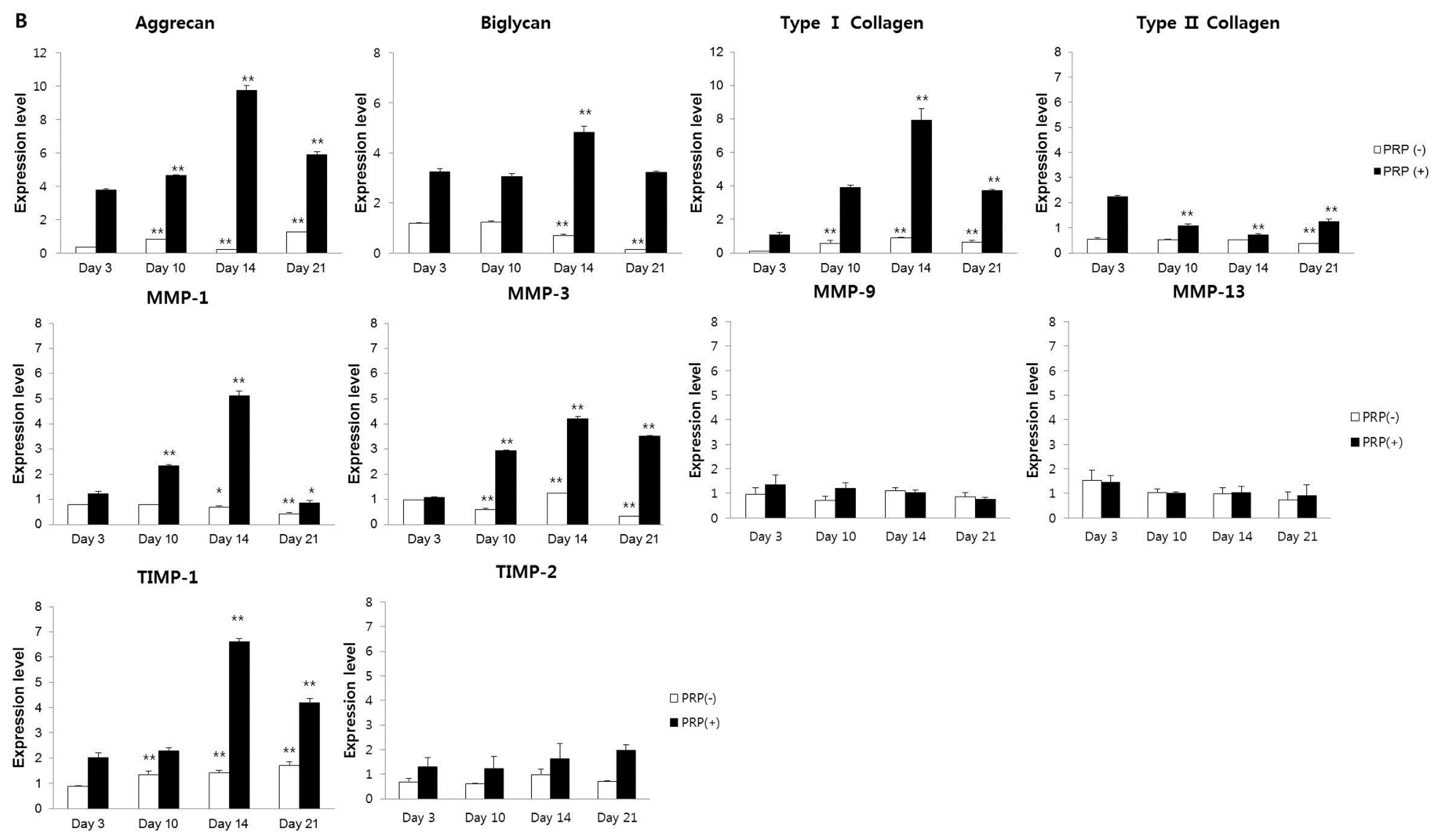
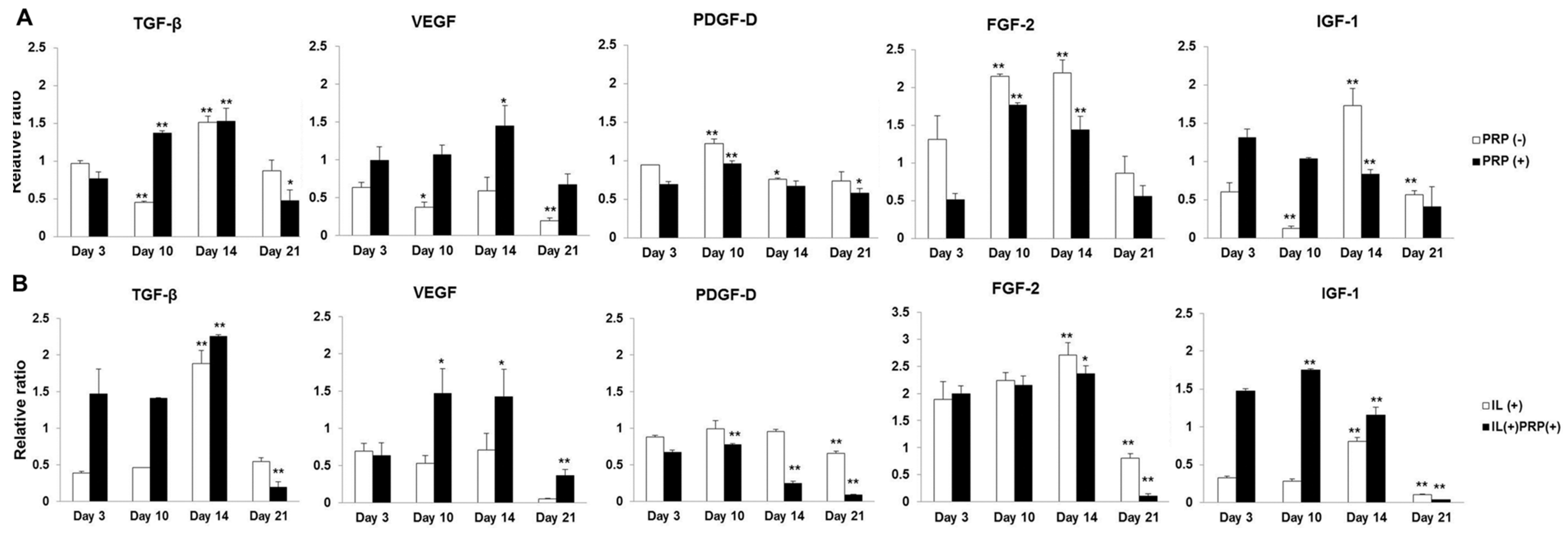
2.1.3. Effects of PRP on Gene and Protein Expression of Meniscal Cells
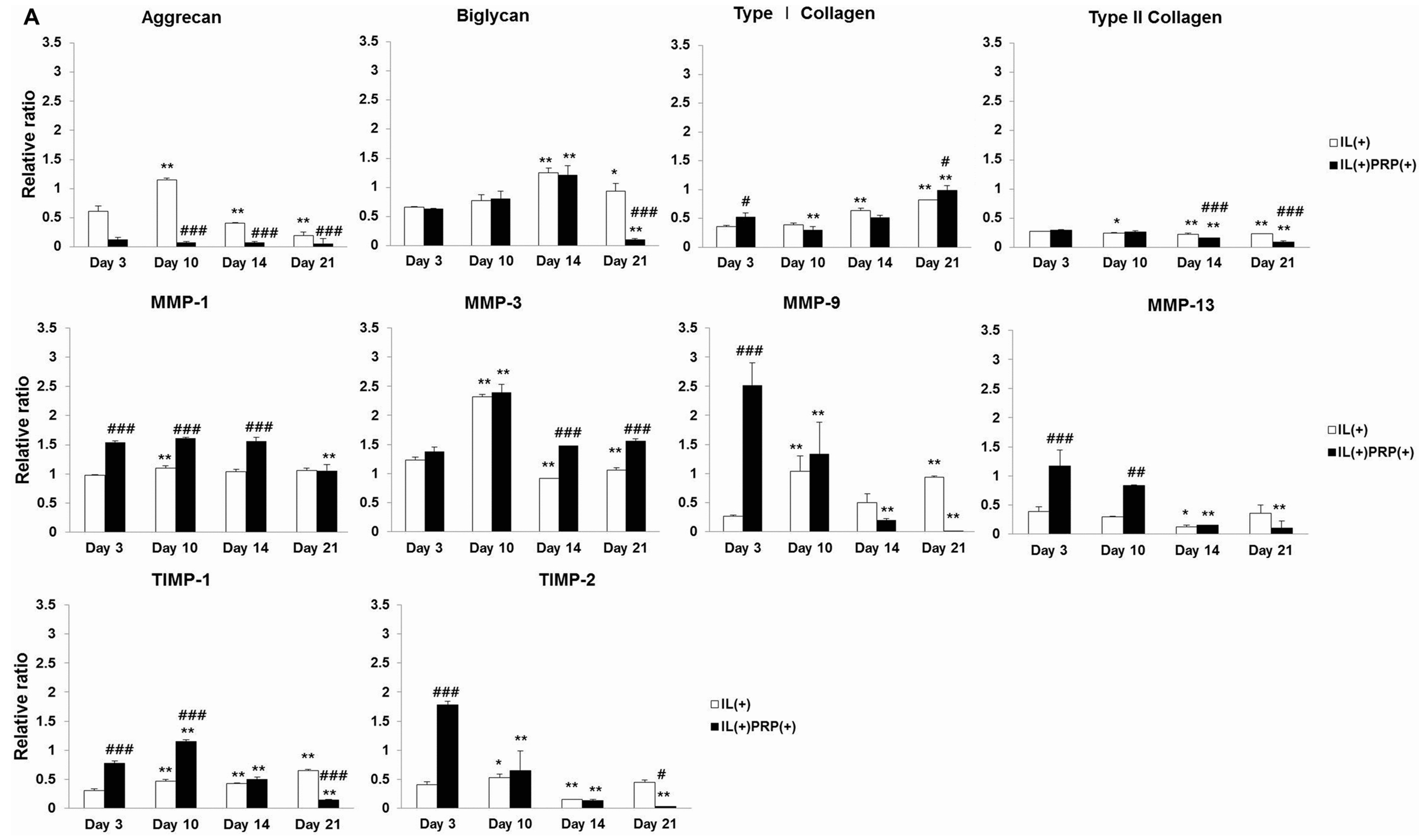
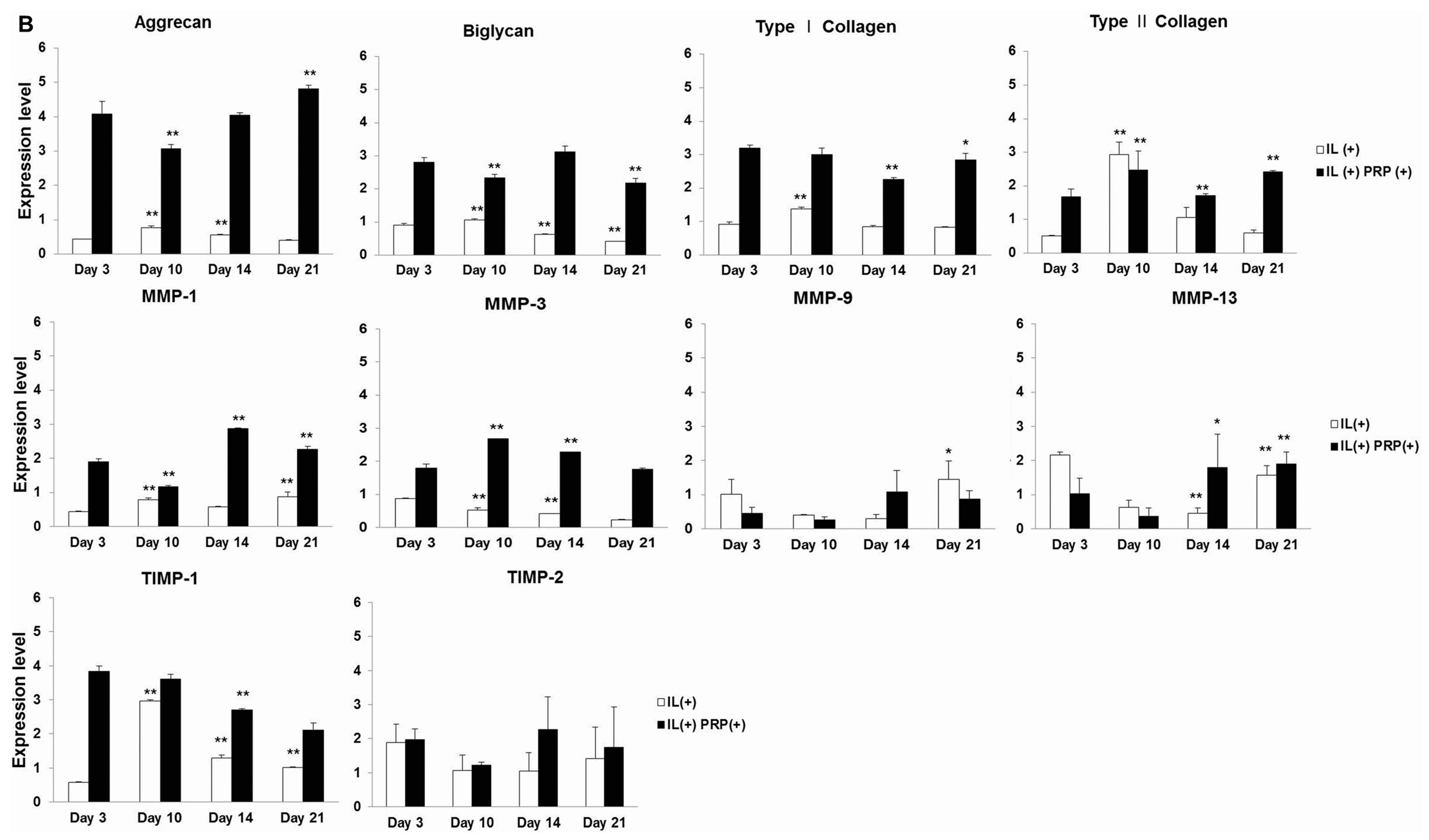
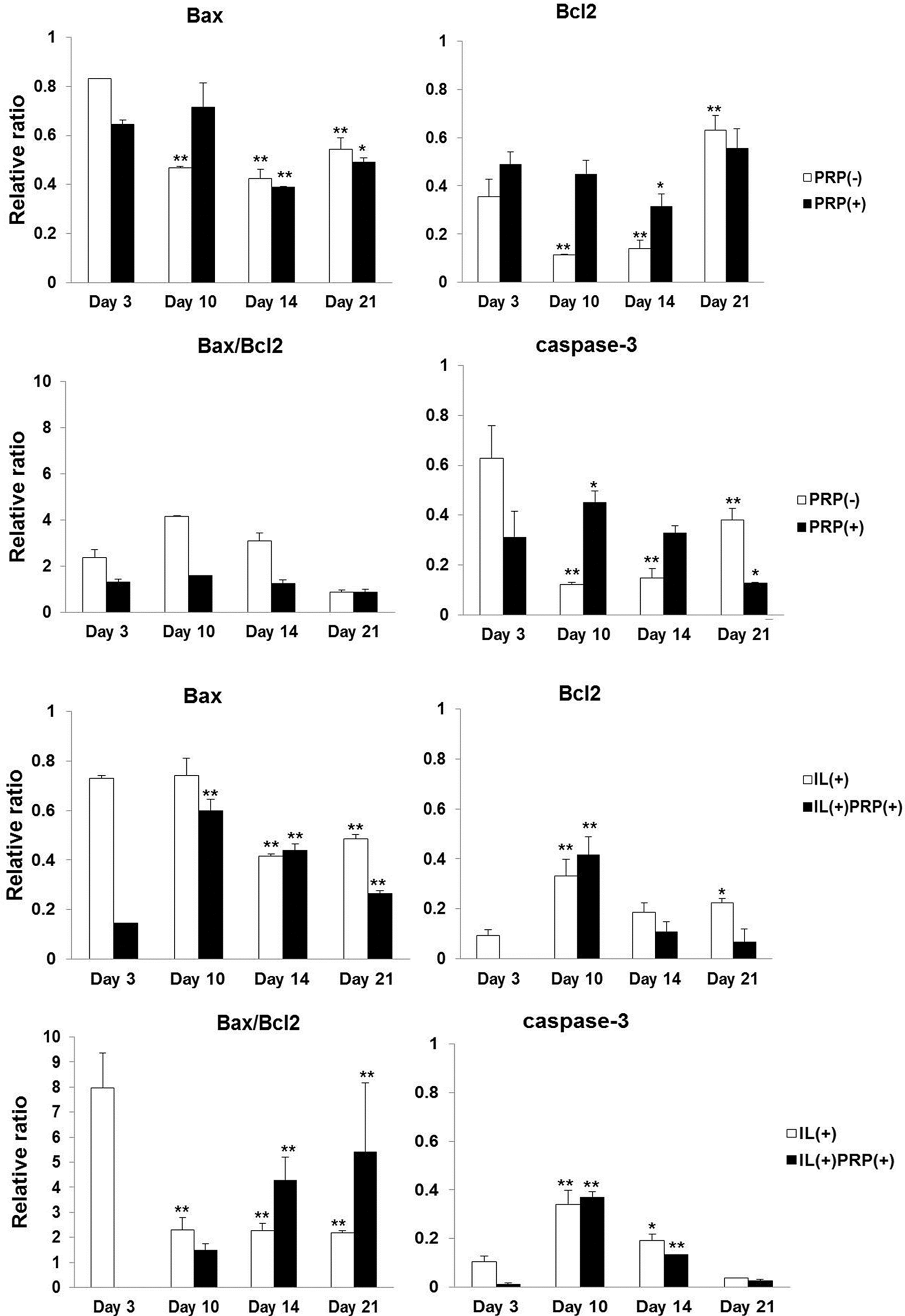
2.1.4. Effects of IL-1α and PRP on Gene and Protein Expression in Meniscal Cells
2.1.5. Temporal Sequential Modulation of PRP-Derived Growth Factors on Meniscal Cells

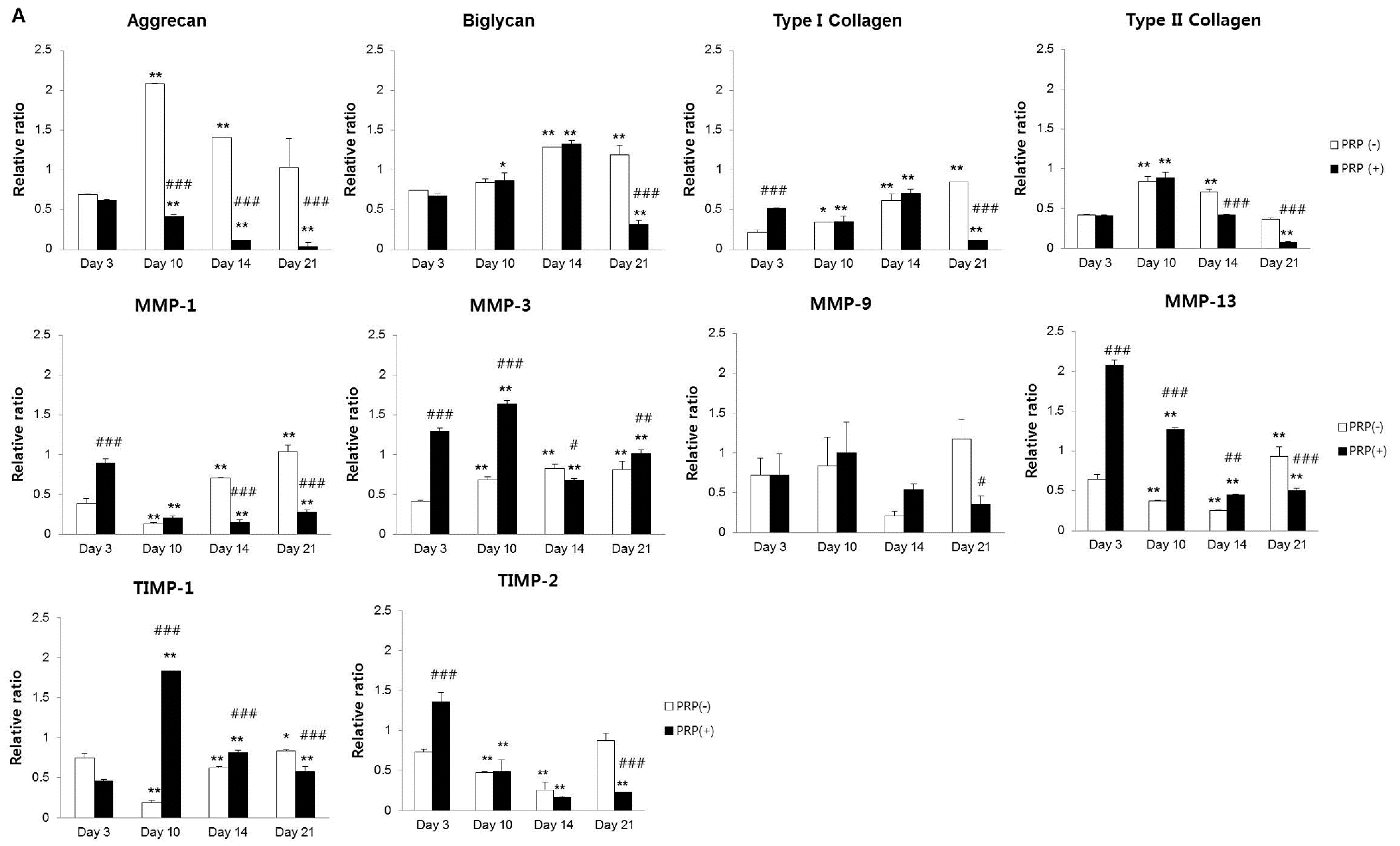

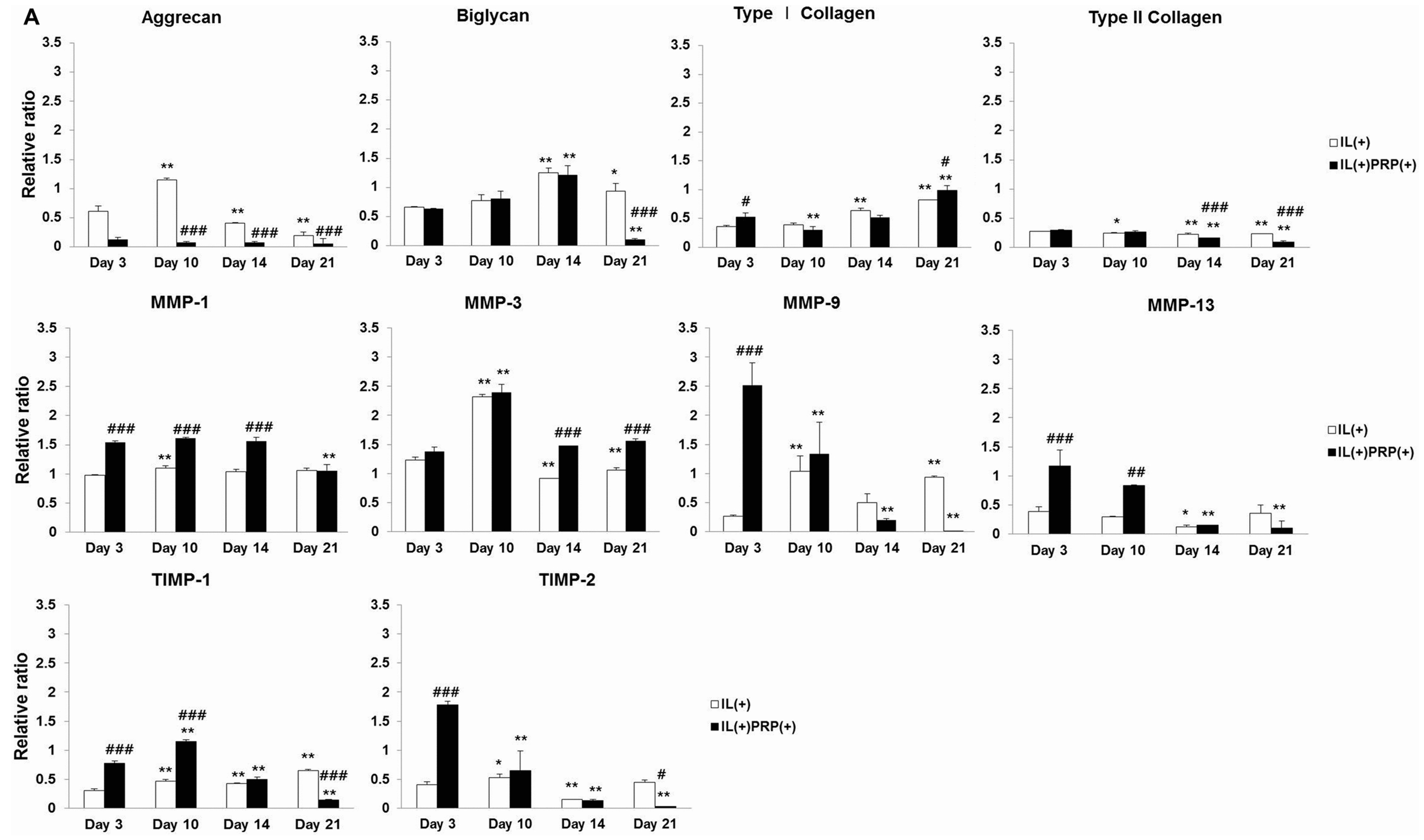
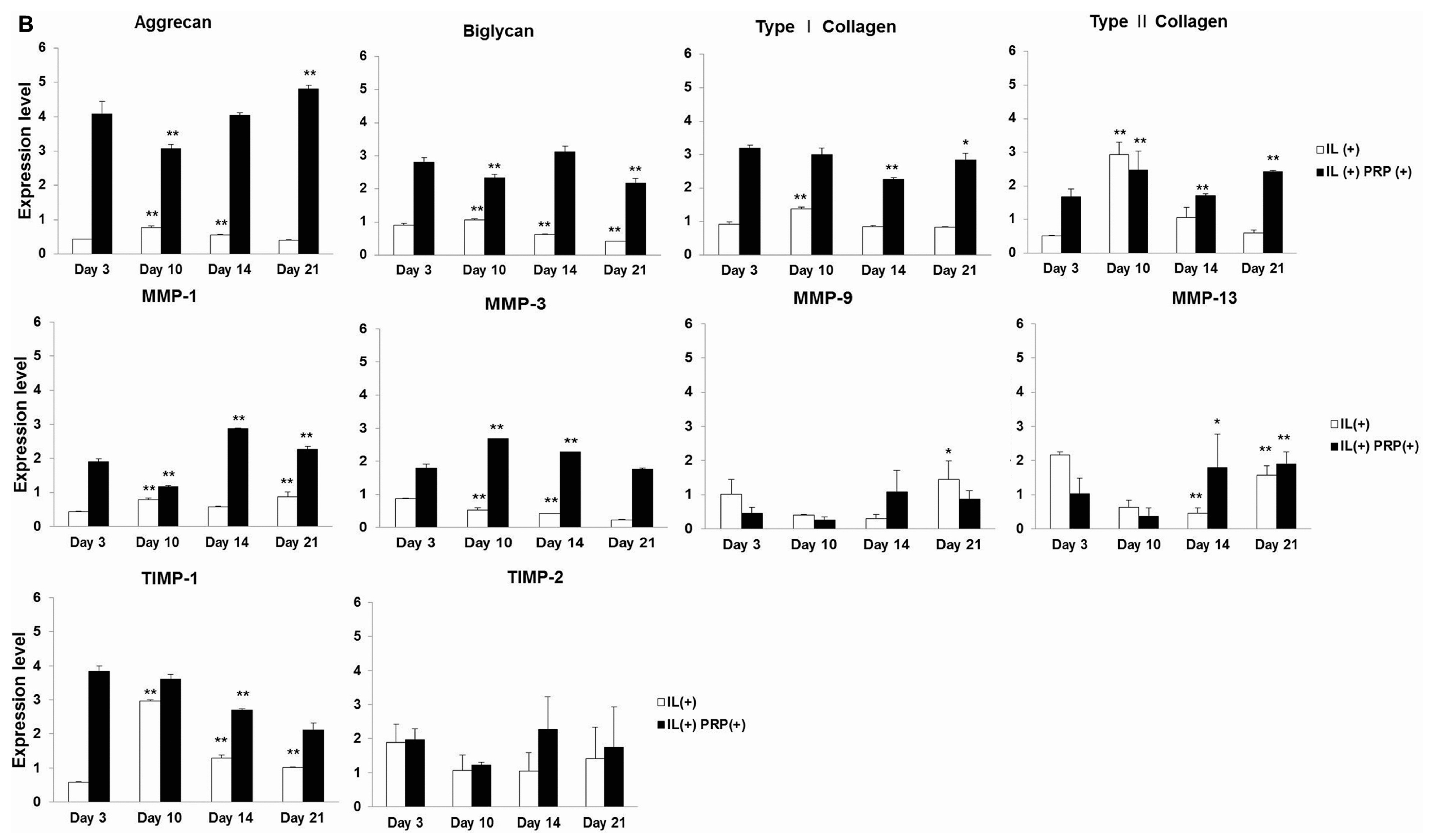
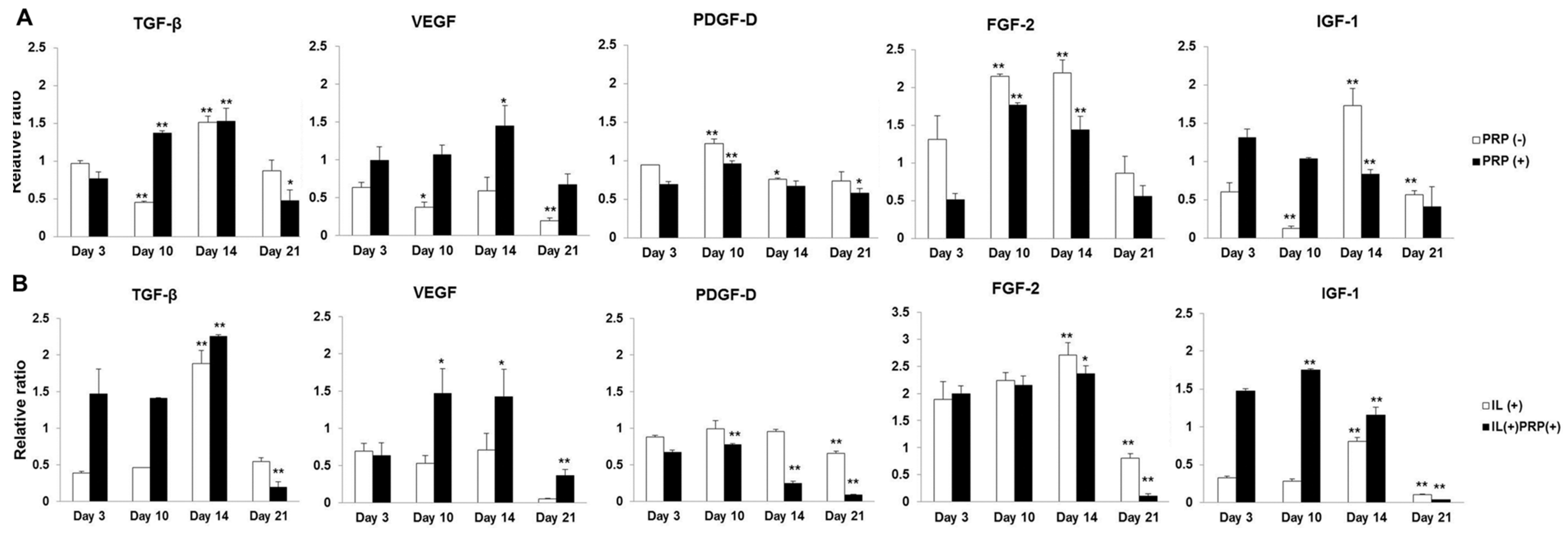

2.1.6. Apoptosis Evaluation of the Effects of PRP on Meniscal Cells
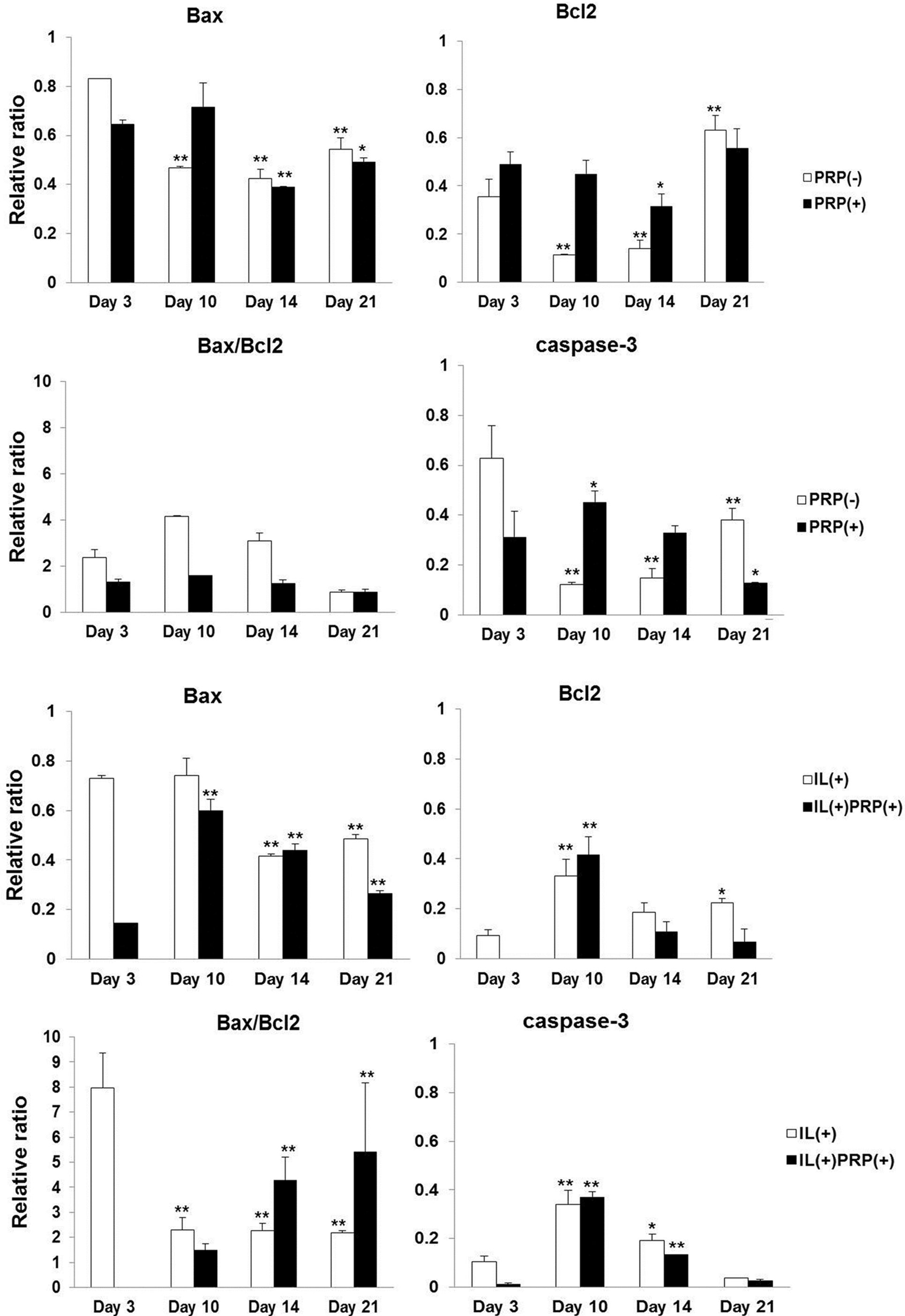
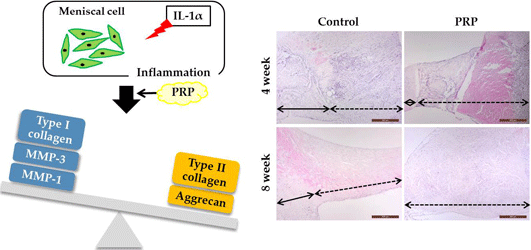
2.2. In Vivo
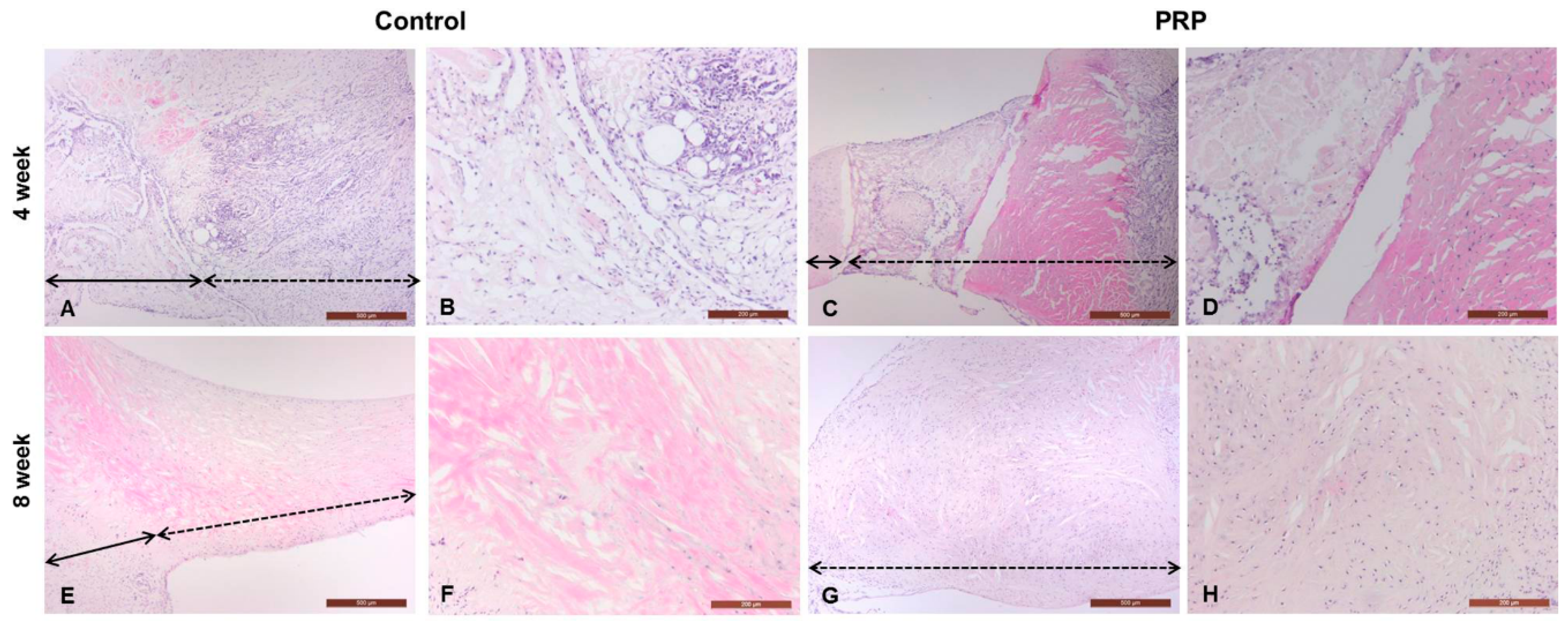
Histopathological Changes in Meniscal Wounds after PRP Implantation

3. Discussion
4. Experimental Section
4.1. Animals
4.2. In Vitro
4.2.1. PRP Preparation
4.2.2. Enzyme-Linked Immunosorbent Assay (ELISA)
4.2.3. Isolation and Culturing of Meniscal Cells
4.2.4. Cell Viability
4.2.5. RNA Extraction and Semi-Quantitative RT-PCR
4.2.6. Immunoblot Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Primer | Gene Bank | Size (bp) | Sequence (5′-3′) |
|---|---|---|---|---|
| Chondrogenesis | GAPDH | DQ403051.1 | 304 | TCA CCA TCT TCC AGG AGC GA |
| CAC AAT GCC GAA GTG GTC GT | ||||
| Aggrecan | XM002723376.1 | 744 | CTC ACC CCG AGA ATC AAA TG | |
| AGG AGG TTT CCG CCG CAG TT | ||||
| Type I Collagen | XM 002713800.1 | 310 | CGC GAT GGT CAG CCT GGA CA | |
| CCG GGA GGG CCA GCA GGA CC | ||||
| Type II Collagen | D 83228.1 | 370 | GAC CCC ATG CAG TAC ATG | |
| AGC CGC CAT TGA TGG TCT CC | ||||
| Biglycan | XM 002722633.1 | 386 | CAC TGC CAC CTG CGG GTT GT | |
| TCT AGG GGG TTC CCG CCC ATC | ||||
| Catabolic molecule | MMP-1 | NM 001171139.1 | 553 | GCA ACC CAG GTG TGG AGT GCC |
| TGG GCC TAC TGG CTG ACT GGG | ||||
| MMP-3 | NM 001082280.1 | 339 | CTG GAG GTT TGA TGA GAA GA | |
| CAG TTC ATG CTC GAG ATT CC | ||||
| MMP-9 | NM 001082203.1 | 519 | GAC GGC AAG CCC TGC GAG TT | |
| TGT GGT GGT GGC TGG AGG CT | ||||
| MMP-13 | NM 001082037.1 | 328 | GCA GCA GTC TCC AGG CAC GG | |
| TCA GGG ACC CCG CAT CTC GG | ||||
| Anabolic molecule | TIMP-1 | NM 001082232.2 | 326 | GCA ACT CCG ACC TTG TCA TC |
| AGC GTA GGT CTT GGT GAA GC | ||||
| TIMP-2 | XM 002723776.1 | 460 | AAC GGA GTC TGG TGG TGC ATT CC | |
| CTT GGC CTG GTG CCC GTT GAT | ||||
| Growth factor | TGF-β | AB 020217.1 | 231 | GCA AGG ACC TGG GCT GGA A |
| AGT AAC ACG ATG GGC AGT GGC | ||||
| VEGF | XM 002714697.1 | 100 | CGC AGC TAC TGC CAG CCG AT | |
| GCA CCA GAG GCA CGC AGG AA | ||||
| PDGF-D | XM 002708534.1 | 125 | TGC ACC GGC TCA TCC TCG TCT A | |
| GTC ATC TCG CCG GAG ATT GGC GTT G | ||||
| FGF-2 | XM 002717238.1 | 278 | GGA GAA GAG CGA CCC ACA CAT CA | |
| TAG CCT TCT GCC CAG GTC CTG TT | ||||
| IGF-1 | NM 001082026.1 | 194 | TCT GCG GTG CTG AGC TGG TG | |
| TGC CTT TGC CGG CTT GAG GG | ||||
| Apoptosis | Bcl2 | DQ 529234.1 | 233 | GTG GGA TAC TGG AGA TGA AGA |
| GAC GGT AGC GAC GAG AGA | ||||
| Bax | XM 002723697.1 | 400 | CCA AGA AGC TGA GCG AGT G | |
| TTC CAG ATG GTG AGT GAG G | ||||
| caspase-3 | NM 001082280.1 | 489 | CAA TGG ACT CTG GGA AAT | |
| GCA AGC CTG AAT AAT GA |
4.3. In Vivo
4.3.1. Meniscal Defect for PRP Application in Rabbits
4.3.2. Histopathological Analysis
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kuettner, K.E.; Cole, A.A. Cartilage degeneration in different human joints. Osteoarthr. Cartil. 2005, 13, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Almarza, A.J.; Athanasiou, K.A. Seeding techniques and scaffolding choice for tissue engineering of the temporomandibular joint disk. Tissue Eng. 2004, 10, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Aigner, T.; Stöve, J. Collagens—Major component of the physiological cartilage matrix, major target of cartilage degeneration, major tool in cartilage repair. Adv. Drug Deliv. Rev. 2003, 55, 1569–1593. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, P.C.; Forsyth, R.G.; Wang, J.; Almqvist, K.F.; Verdonk, R.; Veys, E.M.; Verbruggen, G. Characterisation of human knee meniscus cell phenotype. Osteoarthr. Cartil. 2005, 13, 548–560. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, C.A.; Webber, R.J. The ultrastructure and biochemistry of meniscal cartilage. Clin. Orthop. Relat. Res. 1990, 252, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Collier, S.; Ghosh, P. Effects of transforming growth factor beta on proteoglycan synthesis by cell and explant cultures derived from the knee joint meniscus. Osteoarthr. Cartil. 1995, 3, 127–138. [Google Scholar] [CrossRef]
- Makris, E.A.; Hadidi, P.; Athanasiou, K.A. The knee meniscus: Structure-function, pathophysiology, current repair techniques, and prospects for regeneration. Biomaterials 2011, 32, 7411–7431. [Google Scholar] [CrossRef] [PubMed]
- Mauck, R.L.; Martinez-Diaz, G.J.; Yuan, X.; Tuan, R.S. Regional multilineage differentiation potential of meniscal fibrochondrocytes: Implications for meniscus repair. Anat. Rec. 2007, 290, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Higashioka, M.M.; Chen, J.A.; Hu, J.C.; Athanasiou, K.A. Building an anisotropic meniscus with zonal variations. Tissue Eng. A 2014, 20, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Driscoll, M.D.; Sampson, H.W.; Sekiya, I.; Caroom, C.T.; Prockop, D.J.; Thomas, D.B. Implantation of allogenic synovial stem cells promotes meniscal regeneration in a rabbit meniscal defect model. J. Bone Jt. Surg. Am. 2010, 94, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Scotti, C.; Hirschmann, M.T.; Antinolfi, P.; Martin, I.; Peretti, G.M. Meniscus repair and regeneration: Review on current methods and research potential. Eur. Cell Mater. 2013, 26, 150–170. [Google Scholar] [PubMed]
- Furman, B.D.; Kimmerling, K.A.; Zura, R.D.; Reilly, R.M.; Zlowodzki, M.P.; Huebner, J.L.; Kraus, V.B.; Guilak, F.; Olson, S.A. Articular ankle fracture results in increased synovitis, synovial macrophage infiltration, and synovial fluid concentrations of inflammatory cytokines and chemokines. Arthritis Rheumatol. 2015, 67, 1234–1239. [Google Scholar] [CrossRef] [PubMed]
- Moyer, H.R.; Wang, Y.; Farooque, T.; Wick, T.; Singh, K.A.; Xie, L.; Guldberg, R.E.; Williams, J.K.; Boyan, B.D.; Schwartz, Z. A new animal model for assessing cartilage repair and regeneration at a nonarticular site. Tissue Eng. A 2010, 16, 2321–2330. [Google Scholar] [CrossRef] [PubMed]
- Riera, K.M.; Rothfusz, N.E.; Wilusz, R.E.; Weinberg, J.B.; Guilak, F.; McNulty, A.L. Interleukin-1, tumor necrosis factor-α, and transforming growth factor-β 1 and integrative meniscal repair: Influences on meniscal cell proliferation and migration. Arthritis Res. Ther. 2011, 13, R187. [Google Scholar] [CrossRef] [PubMed]
- Lotz, M. Cytokines in cartilage injury and repair. Clin. Orthop. Relat. Res. 2001, 391, S108–S115. [Google Scholar] [CrossRef] [PubMed]
- Schlaak, J.F.; Pfers, I.; Meyer ZumBüschenfelde, K.H.; Märker-Hermann, E. Different cytokine profiles in the synovial fluid of patients with osteoarthritis, rheumatoid arthritis and seronegative spondylarthropathies. Clin. Exp. Rheumatol. 1996, 14, 155–162. [Google Scholar] [PubMed]
- Ferretti, M.; Madhavan, S.; Deschner, J.; Rath-Deschner, B.; Wypasek, E.; Agarwal, S. Dynamic biophysical strain modulates proinflammatory gene induction in meniscal fibrochondrocytes. Am. J. Physiol. Cell Physiol. 2006, 290, C1610–C1615. [Google Scholar] [CrossRef] [PubMed]
- Upton, M.L.; Chen, J.; Setton, L.A. Region-specific constitutive gene expression in the adult porcine meniscus. J. Orthop. Res. 2006, 24, 1562–1570. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Ibán, M.Á.; Comellas Melero, N.; Martinez-Botas, J.; Ortiz, A.; Diaz Heredia, J. Growth factor expression after lesion creation in the avascular zone of the meniscus: A quantitative PCR study in rabbits. Arthroscopy 2014, 30, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Argentieri, E.C.; Sturnick, D.R.; DeSarno, M.J.; Gardner-Morse, M.G.; Slauterbeck, J.R.; Johnson, R.J.; Beynnon, B.D. Changes to the articular cartilage thickness profile of the tibia following anterior cruciate ligament injury. Osteoarthr. Cartil. 2014, 22, 1453–1460. [Google Scholar] [CrossRef]
- Aghaloo, T.L.; Moy, P.K.; Freymiller, E.G. Investigation of platelet-rich plasma in rabbit cranial defects: A pilot study. J. Oral Maxillofac. Surg. 2002, 60, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Kanthan, S.R.; Kavitha, G.; Addi, S.; Choon, D.S.; Kamarul, T. Platelet-rich plasma (PRP) enhances bone healing in non-united critical-sized defects: A preliminary study involving rabbit models. Injury 2011, 42, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Eppley, B.L.; Woodell, J.E.; Higgins, J. Platelet quantification and growth factor analysis from platelet-rich plasma: Implications for wound healing. Plast. Reconstr. Surg. 2004, 114, 1502–1508. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, L.C.; Lee, G.C.; Huang, K.L.; Mauck, R.L. Growth factor supplementation improves native and engineered meniscus repair in vitro. Acta Biomater. 2012, 8, 3687–3694. [Google Scholar] [CrossRef] [PubMed]
- Izal, I.; Ripalda, P.; Acosta, C.A.; Forriol, F. In vitro healing of avascular meniscal injuries with fresh and frozen plugs treated with TGF-β1 and IGF-1 in sheep. Int. J. Clin. Exp. Pathol. 2008, 1, 426–434. [Google Scholar] [PubMed]
- Tumia, N.S.; Johnstone, A.J. Platelet derived growth factor-AB enhances knee meniscal cell activity in vitro. Knee 2009, 16, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Tummala, P.; King, A.; Lee, B.; Kraus, M.; Tse, V.; Jacobs, C.R. Buffered platelet-rich plasma enhances mesenchymal stem cell proliferation and chondrogenic differentiation. Tissue Eng. C Methods 2009, 15, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Park, K.M.; Joung, Y.K.; Park, K.D.; Do, S.H. Platelet-rich plasma loaded in situ-formed hydrogel enhances hyaline cartilage regeneration by CB1 upregulation. J. Biomed. Mater. Res. A 2012, 100, 3099–3107. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Park, K.M.; Joung, Y.K.; Park, K.D.; Do, S.H. Platelet-rich plasma loaded hydrogel scaffold enhances chondrogenic differentiation and maturation with up-regulation of CB1 and CB2. J. Control. Release 2012, 159, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Buma, P.; Ramrattan, N.N.; van Tienen, T.G.; Veth, R.P. Tissue engineering of the meniscus. Biomaterials 2004, 25, 1523–1532. [Google Scholar] [CrossRef]
- Ishida, K.; Kuroda, R.; Miwa, M.; Tabata, Y.; Hokugo, A.; Kawamoto, T.; Sasaki, K.; Doita, M.; Kurosaka, M. The regenerative effects of platelet-rich plasma on meniscal cells in vitro and its in vivo application with biodegradable gelatin hydrogel. Tissue Eng. 2007, 13, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Zellner, J.; Taeger, C.D.; Schaffer, M.; Roldan, J.C.; Loibl, M.; Mueller, M.B.; Berner, A.; Krutsch, W.; Huber, M.K.; Kujat, R.; et al. Are applied growth factors able to mimic the positive effects of mesenchymal stem cells on the regeneration of meniscus in the avascular zone? Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Fennis, J.P.; Stoelinga, P.J.; Merkx, M.A.; Jansen, J.A. Reconstruction of the mandible with a poly(d,l-lactide) scaffold, autogenous corticocancellous bone graft, and autogenous platelet-rich plasma: An animal experiment. Tissue Eng. 2009, 11, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Ueda, M.; Naiki, T.; Takahashi, M.; Hata, K.; Nagasaka, T. Autogenous injectable bone for regeneration with mesenchymal stem cells and platelet-rich plasma: Tissue-engineered bone regeneration. Tissue Eng. 2004, 10, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Van den Dolder, J.; Mooren, R.; Vloon, A.P.; Stoelinga, P.J.; Jansen, J.A. Platelet-rich plasma: Quantification of growth factor levels and the effect on growth and differentiation of rat bone marrow cells. Tissue Eng. 2006, 12, 3067–3073. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.S.; Song, I.H.; Park, S.Y.; Sung, M.C.; Ahn, M.W.; Song, K.E. Individual variation in growth factor concentrations in platelet-rich plasma and its influence on human mesenchymal stem cells. Korean J. Lab. Med. 2011, 31, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Lee, H.R.; Kim, S.; Ahn, M.W.; Do, S.H. Time-sequential modulation in expression of growth factors from platelet-rich plasma (PRP) on the chondrocyte cultures. Mol. Cell. Biochem. 2012, 361, 9–17. [Google Scholar] [CrossRef]
- Chen, W.H.; Lo, W.C.; Hsu, W.C.; Wei, H.J.; Liu, H.Y.; Lee, C.H.; Tina Chen, S.Y.; Shieh, Y.H.; Williams, D.F.; Deng, W.P. Synergistic anabolic actions of hyaluronic acid and platelet-rich plasma on cartilage regeneration in osteoarthritis therapy. Biomaterials 2014, 35, 9599–9607. [Google Scholar] [CrossRef] [PubMed]
- Narita, A.; Takahara, M.; Sato, D.; Ogino, T.; Fukushima, S.; Kimura, Y.; Tabata, Y. Biodegradable gelatin hydrogels incorporating fibroblast growth factor 2 promote healing of horizontal tears in rabbit meniscus. Arthroscopy 2012, 28, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.W.; Hadeed, M.M.; Werner, B.C.; Diduch, D.R.; Carson, E.W.; Miller, M.D. Platelet-rich plasma in meniscal repair: Does augmentation improve surgical outcomes? Clin. Orthop. Relat. Res. 2015, 473, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Dimauro, I.; Grasso, L.; Fittipaldi, S.; Fantini, C.; Mercatelli, N.; Racca, S.; Geuna, S.; di Gianfrancesco, A.; Caporossi, D.; Pigozzi, F.; et al. Platelet-rich plasma and skeletal muscle healing: A molecular analysis of the early phases of the regeneration process in an experimental animal model. PLoS ONE 2014, 9, e102993. [Google Scholar] [CrossRef] [PubMed]
- McNulty, A.L.; Weinberg, J.B.; Guilak, F. Inhibition of matrix metalloproteinases enhances in vitro repair of the meniscus. Clin. Orthop. Relat. Res. 2009, 467, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Lemke, A.K.; Sandy, J.D.; Voigt, H.; Dreier, R.; Lee, J.H.; Grodzinsky, A.J.; Mentlein, R.; Fay, J.; Schünke, M.; Kurz, B. Interleukin-1α treatment of meniscal explants stimulates the production and release of aggrecanase-generated, GAG-substituted aggrecan products and also the release of pre-formed, aggrecanase-generated G1 and m-calpain-generated G1–G2. Cell Tissue Res. 2010, 340, 179–188. [Google Scholar] [CrossRef]
- Zwierzchowski, T.J.; Stasikowska-Kanicka, O.; Danilewicz, M.; Fabiś, J. Assessment of apoptosis and MMP-1, MMP-3 and TIMP-2 expression in tibial hyaline cartilage after viable medial meniscus transplantation in the rabbit. Arch. Med. Sci. 2012, 8, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Groma, G.; Xin, W.; Grskovic, I.; Niehoff, A.; Brachvogel, B.; Paulsson, M.; Zaucke, F. Abnormal bone quality in cartilage oligomeric matrix protein and matrilin 3 double-deficient mice caused by increased tissue inhibitor of metalloproteinases 3 deposition and delayed aggrecan degradation. Arthritis Rheum. 2012, 64, 2644–2654. [Google Scholar] [CrossRef] [PubMed]
- Toegel, S.; Wu, S.Q.; Otero, M.; Goldring, M.B.; Leelapornpisid, P.; Chiari, C.; Kolb, A.; Unger, F.M.; Windhager, R.; Viernstein, H. Caesalpiniasappan extract inhibits IL1β-mediated overexpression of matrix metalloproteinases in human chondrocytes. Genes Nutr. 2012, 7, 307–318. [Google Scholar] [CrossRef]
- Rai, M.F.; Patra, D.; Sandell, L.J.; Brophy, R.H. Transcriptome analysis of injured human meniscus reveals a distinct phenotype of meniscus degeneration with aging. Arthritis Rheum. 2013, 65, 2090–2101. [Google Scholar] [CrossRef] [PubMed]
- Eppley, B.L.; Pietrzak, W.S.; Blanton, M. Platelet-rich plasma: A review of biology and applications in plastic surgery. Plast. Reconstr. Surg. 2006, 118, 147e–159e. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Akhtar, N.; Khan, A.; Khan, K.A.; Haqqi, T.M. Butrin, isobutrin, and butein from medicinal plant Butea monosperma selectively inhibit nuclear factor-κB in activated human mast cells: Suppression of tumor necrosis factor-α, interleukin (IL)-6, and IL-8. J. Pharmacol. Exp. Ther. 2010, 333, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.; Tol, J.L.; Knez, W.; Chalabi, H. Exercise and the platelet activator calcium chloride both influence the growth factor content of platelet-rich plasma (PRP): Overlooked biochemical factors that could influence PRP treatment. Br. J. Sports Med. 2013, 49, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Abrams, G.D.; Hussey, K.E.; Harris, J.D.; Cole, B.J. Clinical results of combined meniscus and femoral osteochondral allograft transplantation: Minimum 2-year follow-up. Arthroscopy 2014, 30, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Jung, J.C. Suppression of tunicamycin-induced CD44v6 ectodomain shedding and apoptosis is correlated with temporal expression patterns of active ADAM10, MMP-9 and MMP-13 proteins in Caki-2 renal carcinoma cells. Oncol. Rep. 2012, 28, 1869–1874. [Google Scholar] [PubMed]
- Guo, J.; Jie, W.; Shen, Z.; Li, M.; Lan, Y.; Kong, Y.; Guo, S.; Li, T.; Zheng, S. SCF increases cardiac stem cell migration through PI3K/AKT and MMP-2/-9 signaling. Int. J. Mol. Med. 2014, 34, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nagae, M.; Ikeda, T.; Mikami, Y.; Hase, H.; Ozawa, H.; Matsuda, K.; Sakamoto, H.; Tabata, Y.; Kawata, M.; Kubo, T. Intervertebral disc regeneration using platelet-rich plasma and giodegradable gelatin hydrogel microspheres. Tissue Eng. 2007, 13, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Marx, R.E. Platelet-rich plasma: Evidence to support its use. J. Oral Maxillofac. Surg. 2004, 62, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Pauli, C.; Grogan, S.P.; Patil, S.; Otsuki, S.; Hasegawa, A.; Koziol, J.; Lotz, M.K.; D’Lima, D.D. Macroscopic and histopathologic analysis of human knee menisci in aging and osteoarthritis. Osteoarthr. Cartil. 2011, 19, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-R.; Shon, O.-J.; Park, S.-I.; Kim, H.-J.; Kim, S.; Ahn, M.-W.; Do, S.H. Platelet-Rich Plasma Increases the Levels of Catabolic Molecules and Cellular Dedifferentiation in the Meniscus of a Rabbit Model. Int. J. Mol. Sci. 2016, 17, 120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010120
Lee H-R, Shon O-J, Park S-I, Kim H-J, Kim S, Ahn M-W, Do SH. Platelet-Rich Plasma Increases the Levels of Catabolic Molecules and Cellular Dedifferentiation in the Meniscus of a Rabbit Model. International Journal of Molecular Sciences. 2016; 17(1):120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010120
Chicago/Turabian StyleLee, Hye-Rim, Oog-Jin Shon, Se-Il Park, Han-Jun Kim, Sukyoung Kim, Myun-Whan Ahn, and Sun Hee Do. 2016. "Platelet-Rich Plasma Increases the Levels of Catabolic Molecules and Cellular Dedifferentiation in the Meniscus of a Rabbit Model" International Journal of Molecular Sciences 17, no. 1: 120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010120