
Verification of SNPs Associated with Growth Traits in Two Populations of Farmed Atlantic Salmon

 ,
,
Abstract
:
1. Introduction
2. Results
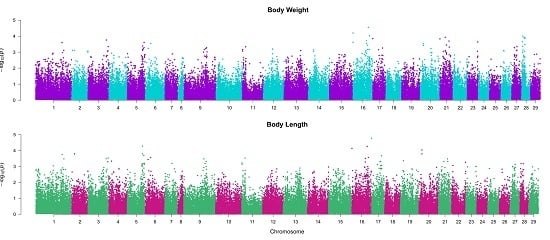
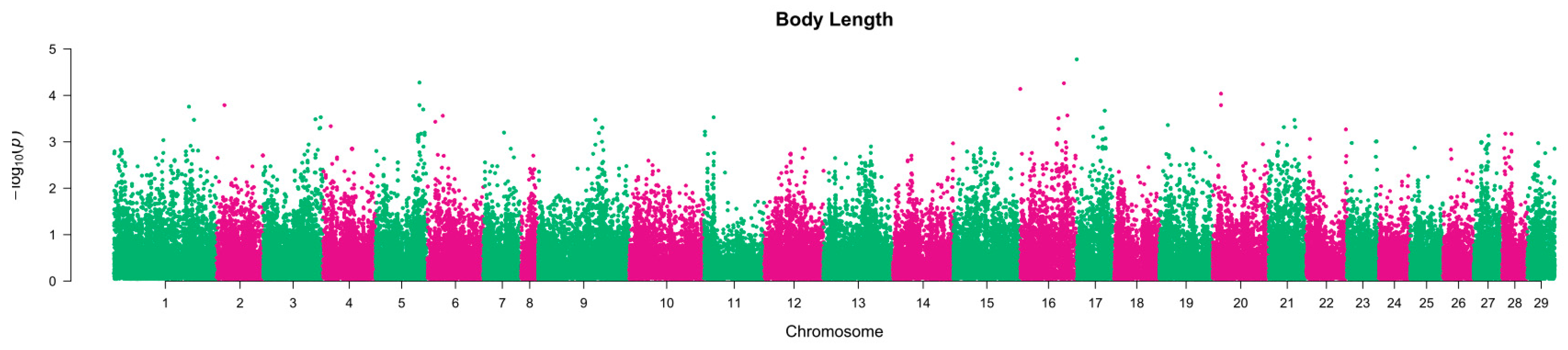
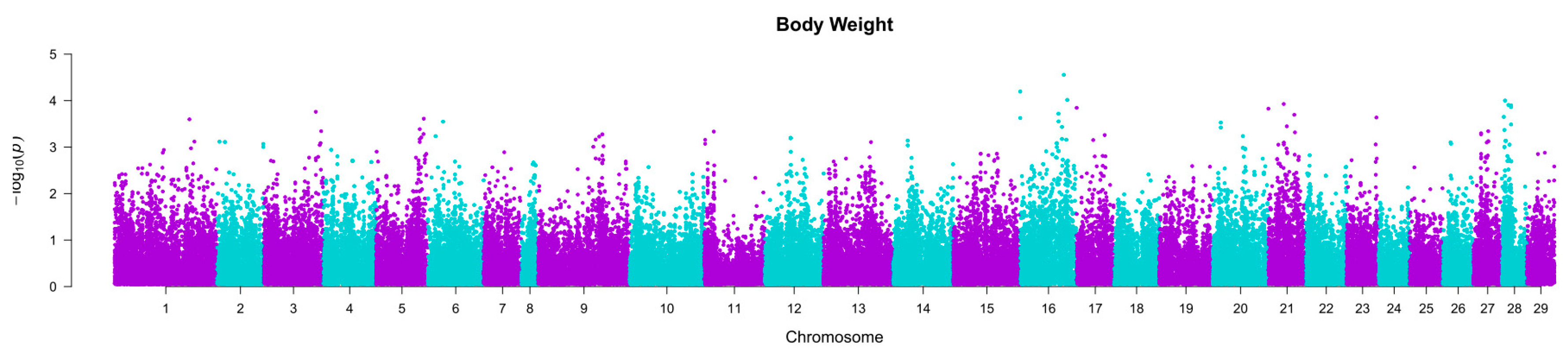
2.1. Heritability Estimation

{kind=link}
{kind=link}
{kind=link}
| Traits | Mean (SD) | Heritability (SE) |
|---|---|---|
| Harvest weight (kg) | 2.65 (0.72) | 0.52 (0.05) |
| Head weight (kg) | 0.30 (0.12) | 0.21 (0.03) |
| Body waste weight (kg) | 0.34 (0.15) | 0.15 (0.02) |
| Total waste weight (kg) | 0.67 (0.21) | 0.32 (0.04) |
| Gutted weight (kg) | 2.42 (0.65) | 0.53 (0.05) |
| Deheaded weight (kg) | 2.11 (0.57) | 0.52 (0.05) |
| Fillet weight (kg) | 1.76 (0.48) | 0.53 (0.05) |
| Fat percentage (%) | 13.2 (5.98) | 0.18 (0.03) |
| Fillet colour (20–34) | 28.9 (0.74) | 0.14 (0.03) |
| Gut weight (kg) | 0.22 (0.08) | 0.30 (0.04) |
2.2. Association between SNPs and Traits of Interest
| Traits | AX88141678 (Gene: MEP1A) | AX88270804 (Gene: PCNT) | ||||||
|---|---|---|---|---|---|---|---|---|
| A/A | A/G | G/G | PVE (%) | A/A | A/G | G/G | PVE (%) | |
| # of fish | n = 651 | n = 436 | n = 52 | n = 281 | n = 581 | n = 265 | ||
| Harvest weight | 2.59 (0.04) | 2.63 (0.05) | 2.33 (0.1) ** | 0.3 | 2.66 (0.05) | 2.60 (0.04) | 2.50 (0.06) * | 1 |
| Head weight | 0.30 (0.01) | 0.30 (0.01) | 0.26 (0.02) ** | 1.3 | 0.31 (0.01) | 0.3 (0.01) | 0.28 (0.01) ** | 1 |
| Body waste weight | 0.33 (0.01) | 0.35 (0.01) | 0.32 (0.02) | 0.1 | 0.34 (0.01) | 0.35 (0.01) | 0.31 (0.01) ** | 3 |
| Total waste weight | 0.65 (0.01) | 0.67 (0.01) | 0.61 (0.03) | 0 | 0.66 (0.02) | 0.67 (0.01) | 0.61 (0.02) ** | 2 |
| Gutted weight | 2.37 (0.04) | 2.39 (0.04) | 2.19 (0.09) * | 0.2 | 2.41 (0.05) | 2.38 (0.04) | 2.28 (0.05) ** | 1 |
| Deheaded weight | 2.07 (0.03) | 2.10 (0.03) | 1.94 (0.08) | 0.05 | 2.10 (0.04) | 2.09 (0.03) | 1.99 (0.04) * | 1 |
| Fillet weight | 1.71 (0.03) | 1.76 (0.03) | 1.59 (0.07) ** | 0 | 1.76 (0.04) | 1.72 (0.03) | 1.67 (0.04) | 1 |
| Fat percentage | 13.19 (0.27) | 13.12 (0.31) | 12.41 (0.84) | 0.4 | 13.65 (0.39) | 13.17 (0.28) | 12.45 (0.4) * | 4 |
| Fillet colour | 28.96 (0.04) | 28.90 (0.05) | 29.03 (0.12) | 0.1 | 28.98 (0.06) | 28.90 (0.04) | 28.97 (0.06) | 0.02 |
| Gut weight | 0.21 (0) | 0.22 (0) | 0.20 (0.01) | 0.01 | 0.22 (0.01) | 0.22 (0) | 0.20 (0.01) ** | 3 |
2.3. QTL Region Characterization and Putative Gene Identification
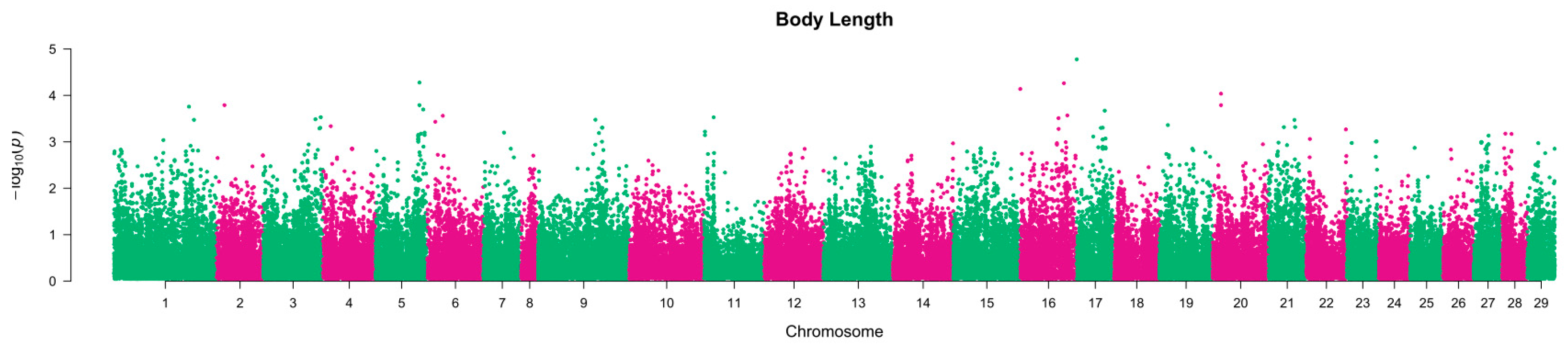
3. Discussion
4. Experimental Section
4.1. Animals
4.2. SNP Selection and Genotyping
4.3. Statistical Analysis
4.3.1. Heritability Estimation and SNP Associations
4.3.2. Allelic Substitution Assessment
4.4. Candidate Gene Identification
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- FAO. Yearbook. Fishery and Aquaculture Statistics; F.A.O.: Rome, Italy, 2010. [Google Scholar]
- Yáñez, J.M.; Newman, S.; Houston, R.D. Genomics in aquaculture to better understand species biology and accelerate genetic progress. Front. Genet. 2015, 6, 128. [Google Scholar] [PubMed]
- Bishop, S.C.; Woolliams, J.A. Genomics and disease resistance studies in livestock. Livest. Sci. 2014, 166, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Gjedrem, T.; Robinson, N.; Rye, M. The importance of selective breeding in aquaculture to meet future demands for animal protein: A review. Aquaculture 2012, 350–353, 117–129. [Google Scholar] [CrossRef]
- Lien, S.; Gidskehaug, L.; Moen, T.; Hayes, B.J.; Berg, P.R.; Davidson, W.S.; Omholt, S.W.; Kent, M.P. A dense SNP-based linkage map for Atlantic salmon (Salmo salar) reveals extended chromosome homeologies and striking differences in sex-specific recombination patterns. BMC Genom. 2011, 12, 615. [Google Scholar] [CrossRef] [PubMed]
- Houston, R.D.; Taggart, J.B.; Cézard, T.; Bekaert, M.; Lowe, N.R.; Downing, A.; Talbot, R.; Bishop, S.C.; Archibald, A.L.; Bron, J.E.; et al. Development and validation of a high density SNP genotyping array for Atlantic salmon (Salmo salar). BMC Genom. 2014, 15, 90. [Google Scholar]
- Palti, Y.; Gao, G.; Liu, S.; Kent, M.P.; Lien, S.; Miller, M.R.; Rexroad, C.E.; Moen, T. The Development and Characterization of a 57K SNP Array for Rainbow Trout. Mol. Ecol. Resour. 2015, 15, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.C.; Smith, T.P.L.; Wiedmann, R.T.; Vallejo, R.L.; Salem, M.; Yao, J.; Rexroad, C.E. Single nucleotide polymorphism discovery in rainbow trout by deep sequencing of a reduced representation library. BMC Genom. 2009, 10, 559. [Google Scholar] [CrossRef] [PubMed]
- Everett, M.V.; Seeb, J.E. Detection and mapping of QTL for temperature tolerance and body size in Chinook salmon (Oncorhynchus tshawytscha) using genotyping by sequencing. Evol. Appl. 2014, 7, 480–492. [Google Scholar]
- Gutierrez, A.P.; Yáñez, J.M.; Fukui, S.; Swift, B.; Davidson, W.S. Genome-Wide Association Study (GWAS) for Growth Rate and Age at Sexual Maturation in Atlantic Salmon (Salmo salar). PLoS ONE 2015, 10, e0119730. [Google Scholar] [CrossRef] [PubMed]
- Sodeland, M.; Gaarder, M.; Moen, T.; Thomassen, M.; Kjøglum, S.; Kent, M.; Lien, S. Genome-wide association testing reveals quantitative trait loci for fillet texture and fat content in Atlantic salmon. Aquaculture 2013, 408–409, 169–174. [Google Scholar] [CrossRef]
- Houston, R.D.; Bishop, S.C.; Guy, D.R.; Tinch, A.E.; Taggart, J.B.; Bron, J.E.; Downing, A.; Stear, M.J.; Gharbi, K.; Hamilton, A. Genome Wide Association Analysis for Resistance to Sea Lice in Atlantic Salmon: Application of a Dense SNP Array. In Proceedings of the 10th World Congress on Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 August 2014; pp. 10–12.
- Correa, K.; Lhorente, J.; Lopez, M.; Bassini, L.; Naswa, S.; Deeb, N.; di Genova, A.; Maas, A.; Davidson, W.; Yáñez, J. Genome-wide association analysis reveals loci associated with resistance against Piscirickettsia salmonis in two Atlantic salmon (Salmo salar L.) chromosomes. BMC Genom. 2015, 16, 854. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Hamilton, A.; Tinch, A.E.; Guy, D.R.; Gharbi, K.; Stear, M.J.; Oswald, M.; Bishop, S.C.; Houston, R.D. Genome wide association and genomic prediction for growth traits in juvenile farmed Atlantic salmon using a high density SNP array. BMC Genom. 2015, 16, 969. [Google Scholar] [CrossRef] [PubMed]
- Macqueen, D.J.; Johnston, I.A. A well-constrained estimate for the timing of the salmonid whole genome duplication reveals major decoupling from species diversification. Proc. R. Soc. B 2014, 281, 1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthelot, C.; Brunet, F.; Chalopin, D.; Juanchich, A.; Bernard, M.; Noël, B.; Bento, P.; Da Silva, C.; Labadie, K.; Alberti, A.; et al. The rainbow trout genome provides novel insights into evolution after whole-genome duplication in vertebrates. Nat. Commun. 2014, 5, 3657. [Google Scholar]
- Gidskehaug, L.; Kent, M.; Hayes, B.J.; Lien, S. Genotype calling and mapping of multisite variants using an Atlantic salmon iSelect SNP array. Bioinformatics 2011, 27, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H. Yesterday’s polyploids and the mystery of diploidization. Nat. Rev. Genet. 2001, 2, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Hamilton, A.; Guy, D.R.; Houston, R.D. Single nucleotide polymorphisms in the insulin-like growth factor 1 (IGF1) gene are associated with growth-related traits in farmed Atlantic salmon. Anim. Genet. 2014, 45, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Hamilton, A.; Guy, D.R.; Tinch, A.E.; Bishop, S.C.; Houston, R.D. The genetic architecture of growth and fillet traits in farmed Atlantic salmon (Salmo salar). BMC Genet. 2015, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.; White, I.; Guy, D.; Brotherstone, S. Genetic parameters of production traits in Atlantic salmon (Salmo salar). Aquaculture 2008, 274, 225–231. [Google Scholar] [CrossRef]
- Mackay, T.F.C.; Stone, E.A.; Ayroles, J.F. The genetics of quantitative traits: challenges and prospects. Nat. Rev. Genet. 2009, 10, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar]
- Moghadam, H.K.; Poissant, J.; Fotherby, H.; Haidle, L.; Ferguson, M.M.; Danzmann, R.G. Quantitative trait loci for body weight, condition factor and age at sexual maturation in Arctic charr (Salvelinus alpinus): comparative analysis with rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Mol. Genet. Genom. 2007, 277, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Peñaloza, C.; Hamilton, A.; Guy, D.R.; Bishop, S.C.; Houston, R.D. A SNP in the 5’ flanking region of the myostatin-1b gene is associated with harvest traits in Atlantic salmon (Salmo salar). BMC Genet. 2013, 14, 112. [Google Scholar] [CrossRef] [PubMed]
- Davidson, W.S.; Koop, B.F.; Jones, S.J.M.; Iturra, P.; Vidal, R.; Maass, A.; Jonassen, I.; Lien, S.; Omholt, S.W. Sequencing the genome of the Atlantic salmon (Salmo salar). Genome Biol. 2010, 11, 403. [Google Scholar] [PubMed]
- Yáñez, J.M.; Houston, R.D.; Newman, S. Genetics and genomics of disease resistance in salmonid species. Front. Genet. 2014, 5, 415. [Google Scholar] [PubMed]
- Rauch, A.; Thiel, C.T.; Schindler, D.; Wick, U.; Crow, Y.J.; Ekici, A.B.; van Essen, A.J.; Goecke, T.O.; Al-Gazali, L.; Chrzanowska, K.H.; et al. Mutations in the Pericentrin (PCNT) Gene Cause Primordial Dwarfism. Science 2008, 319, 816–819. [Google Scholar]
- Müller, T.; Rumpel, E.; Hradetzky, S.; Bollig, F.; Wegner, H.; Blumenthal, A.; Greinacher, A.; Endlich, K.; Endlich, N. Non-muscle myosin IIA is required for the development of the zebrafish glomerulus. Kidney Int. 2011, 80, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Sterchi, E.E.; Stöcker, W.; Bond, J.S. Meprins, membrane-bound and secreted astacin metalloproteinases. Mol. Asp. Med. 2010, 29, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, S.; McDaniel, N.K.; Sugiura, S.H.; Soteropoulos, P.; Tian, B.; Fletcher, J.W.; Ferraris, R.P. Salmonid microarrays identify intestinal genes that reliably monitor P deficiency in rainbow trout aquaculture. Anim. Genet. 2007, 38, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Sauvage, C.; Derôme, N.; Normandeau, E.; St-Cyr, J.; Audet, C.; Bernatchez, L. Fast transcriptional responses to domestication in the brook charr Salvelinus fontinalis. Genetics 2010, 185, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.P.; Lubieniecki, K.P.; Davidson, E.A.; Lien, S.; Kent, M.P.; Fukui, S.; Withler, R.E.; Swift, B.; Davidson, W.S. Genetic mapping of quantitative trait loci (QTL) for body-weight in Atlantic salmon (Salmo salar) using a 6.5K SNP array. Aquaculture 2012, 358–359, 61–70. [Google Scholar] [CrossRef]
- Baranski, M.; Moen, T.; Våge, D.I. Mapping of quantitative trait loci for flesh colour and growth traits in Atlantic salmon (Salmo salar). Genet. Sel. Evol. 2010, 42, 17. [Google Scholar] [CrossRef] [PubMed]
- Houston, R.D.; Bishop, S.C.; Hamilton, A.; Guy, D.R.; Tinch, A.E.; Taggart, J.B.; Derayat, A.; McAndrew, B.J.; Haley, C.S. Detection of QTL affecting harvest traits in a commercial Atlantic salmon population. Anim. Genet. 2009, 40, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Bastiaansen, J.W.M.; Bovenhuis, H.; Lopes, M.S.; Silv, F.F.; Megens, H.J.; Calu, M.P.L. SNP Effects Depend on Genetic and Environmental Context. In Proceedings of the 10th World Congress on Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 August 2014; pp. 356–362.
- Odegård, J.; Moen, T.; Santi, N.; Korsvoll, S.A.; Kjøglum, S.; Meuwissen, T.H.E. Genomic prediction in an admixed population of Atlantic salmon (Salmo salar). Front. Genet. 2014, 5, 402. [Google Scholar] [PubMed]
- LGC Genomics KASP (kompetitive allele-specific PCR) assays. Available online: http://www.lgcgroup.com/products/kasp-genotyping-chemistry/kasp-technicalresources/#.VWC3TU9VhXt (accessed on 1 November 2015).
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Thompson, R. ASReml User Guide, 4th ed.; VSN International Ltd: Hemel Hempstead, UK, 2014. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longmans Green: Harlow, Essex, UK, 1996. [Google Scholar]
- Centre for biomedical research, RepeatMasker, University of Victoria. Available online: http://lucy.ceh.uvic.ca/repeatmasker/cbr_repeatmasker.py (accessed on 30 August 2015).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, H.Y.; Hamilton, A.; Guy, D.R.; Tinch, A.E.; Bishop, S.C.; Houston, R.D. Verification of SNPs Associated with Growth Traits in Two Populations of Farmed Atlantic Salmon. Int. J. Mol. Sci. 2016, 17, 5. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010005
Tsai HY, Hamilton A, Guy DR, Tinch AE, Bishop SC, Houston RD. Verification of SNPs Associated with Growth Traits in Two Populations of Farmed Atlantic Salmon. International Journal of Molecular Sciences. 2016; 17(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010005
Chicago/Turabian StyleTsai, Hsin Y., Alastair Hamilton, Derrick R. Guy, Alan E. Tinch, Steve C. Bishop, and Ross D. Houston. 2016. "Verification of SNPs Associated with Growth Traits in Two Populations of Farmed Atlantic Salmon" International Journal of Molecular Sciences 17, no. 1: 5. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010005