
Zinc Finger and X-Linked Factor (ZFX) Binds to Human SET Transcript 2 Promoter and Transactivates SET Expression

Abstract
:1. Introduction
2. Results
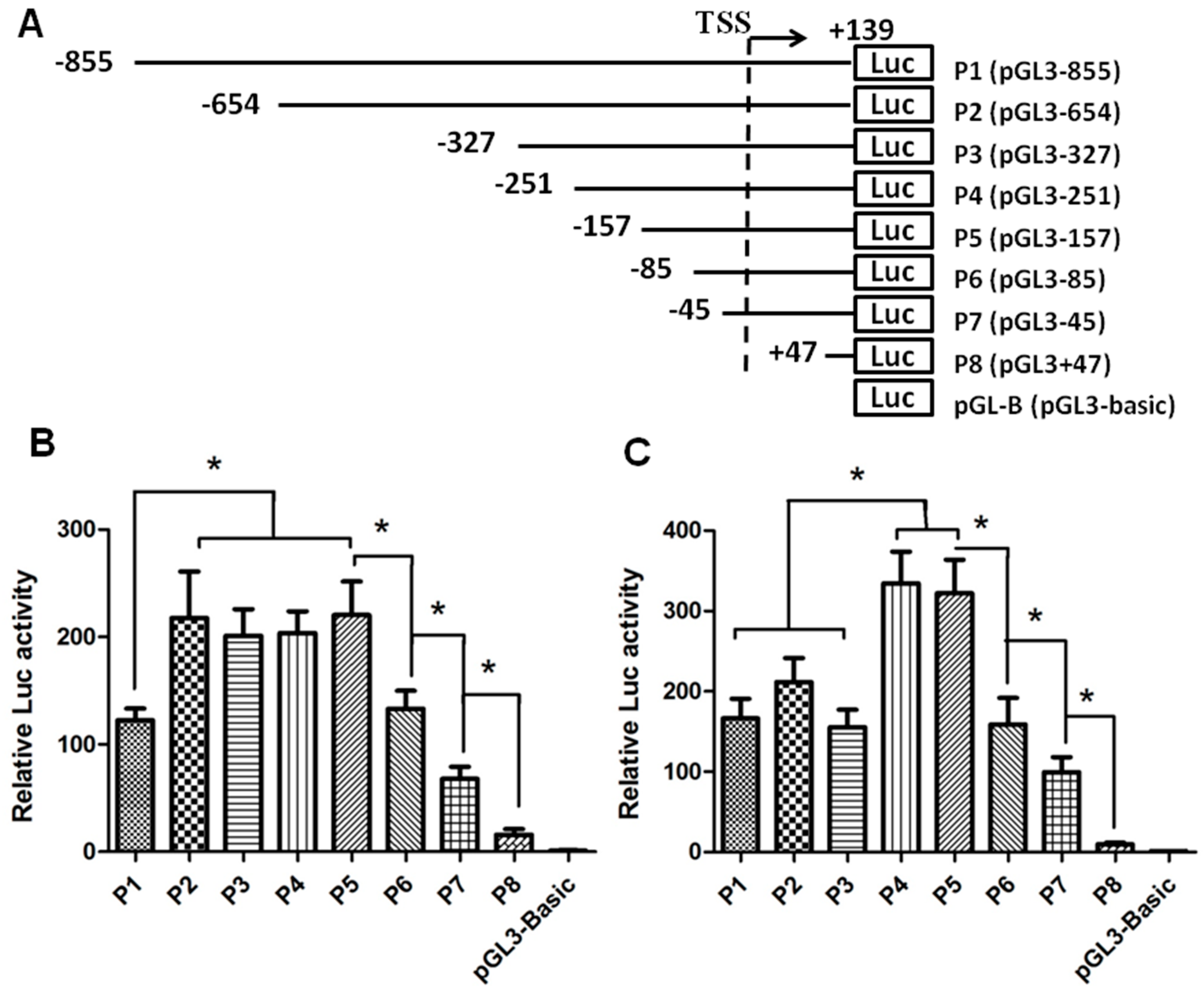
2.1. Identification of the Core Promoter Region Regulating the Expression of SET (SE Translocation) Transcript 2
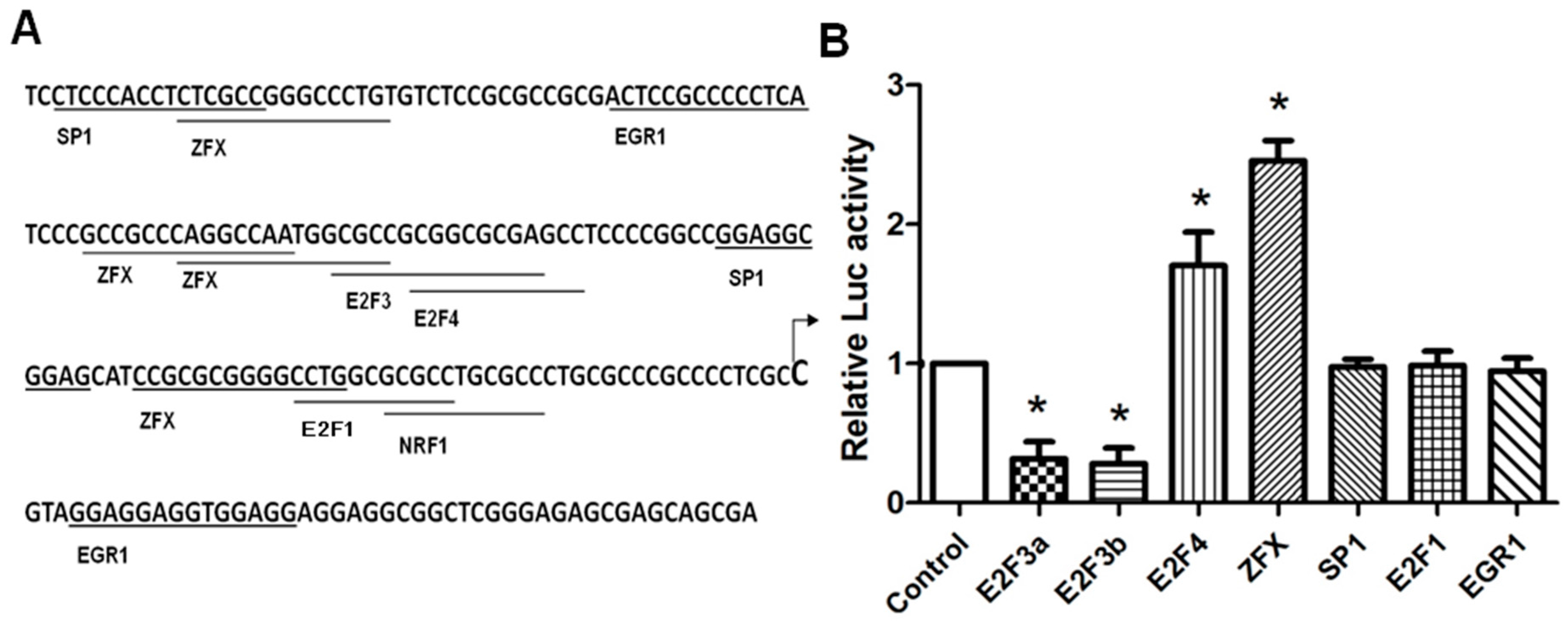
2.2. Identification of Crucial Transcription Factors Controlling SET Promoter Activity
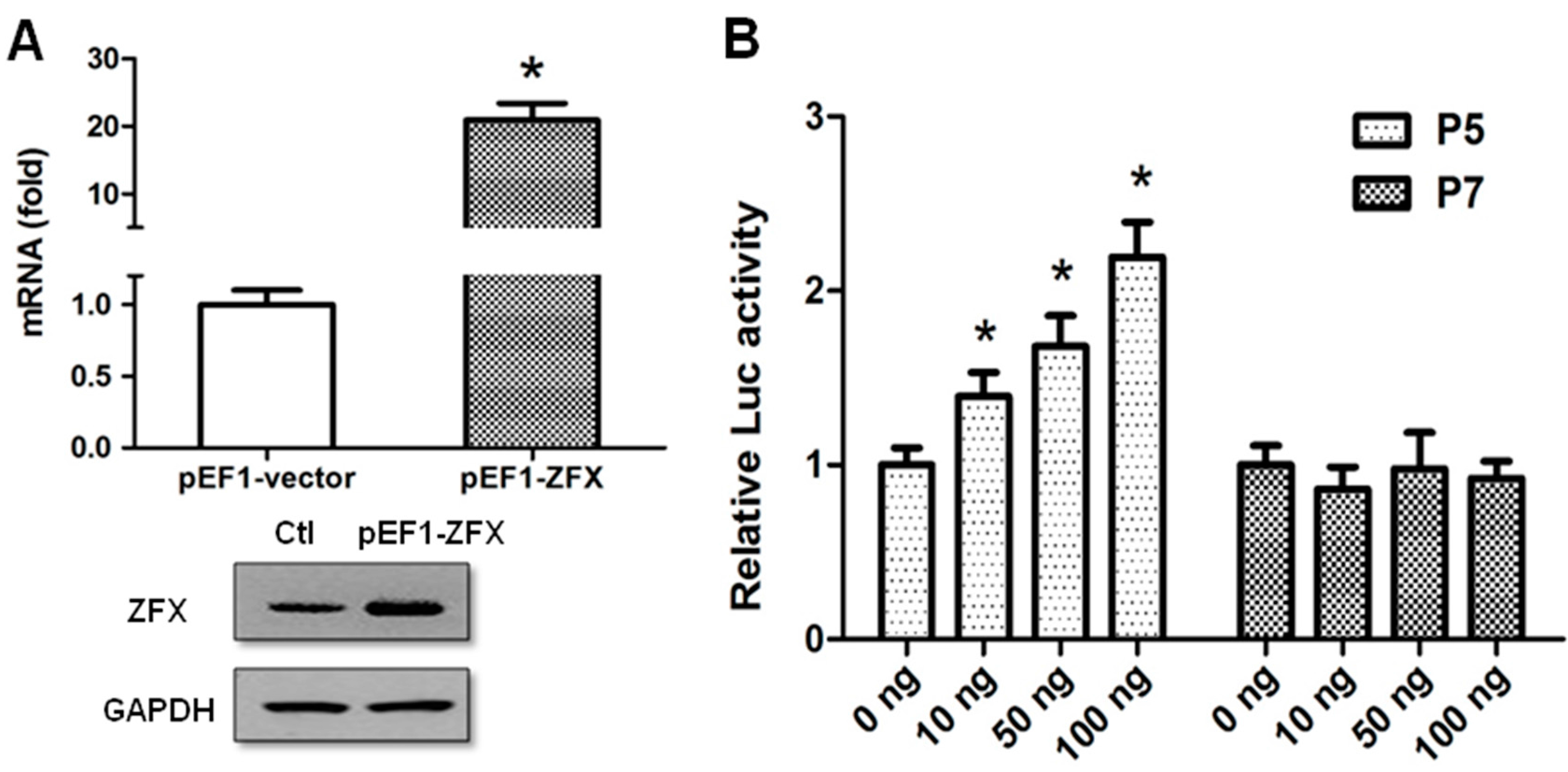
2.3. ZFX Transactivates Human SET Promoter
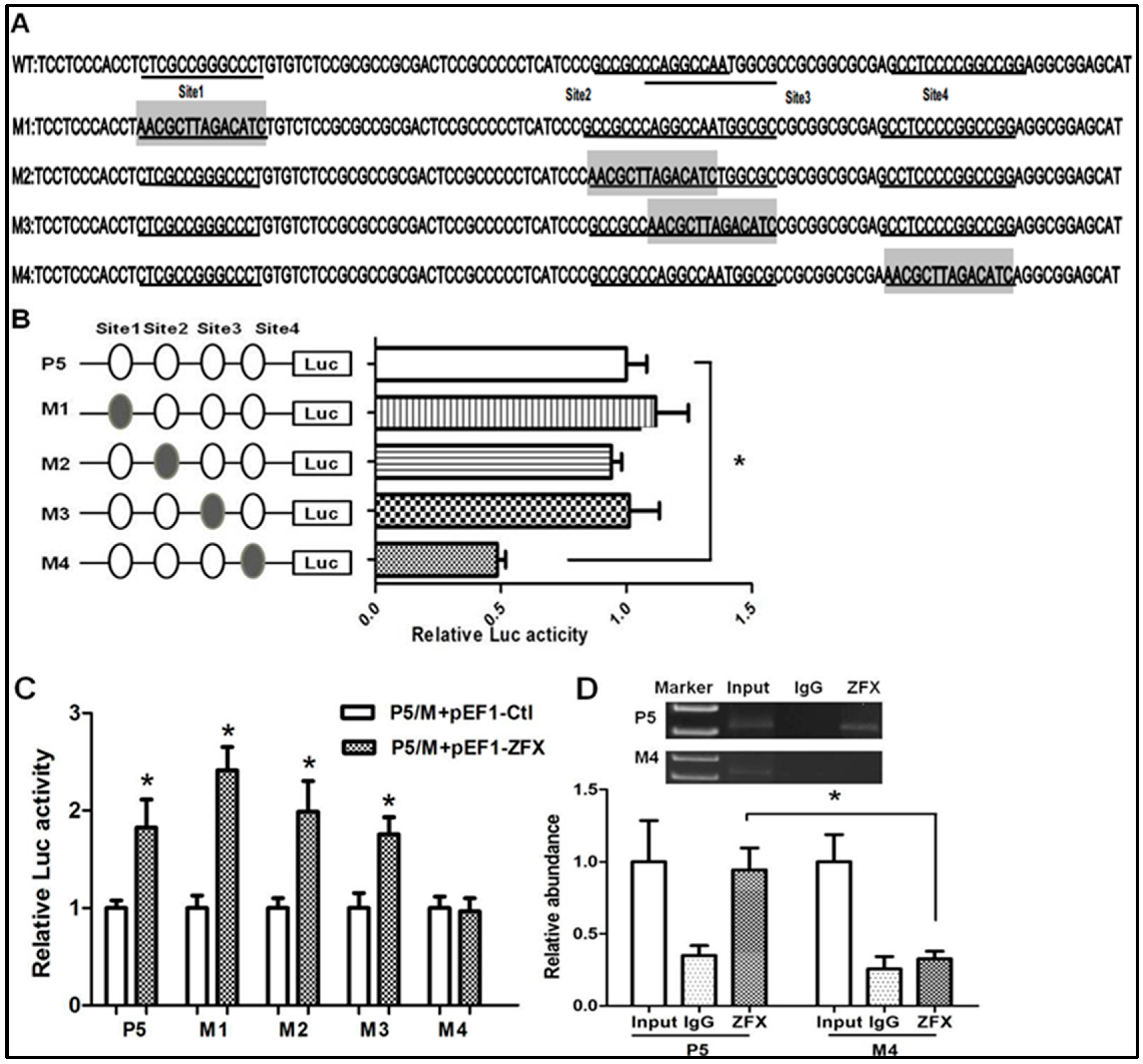
2.4. Mutagenesis Study on the Putative ZFX-Binding Sites
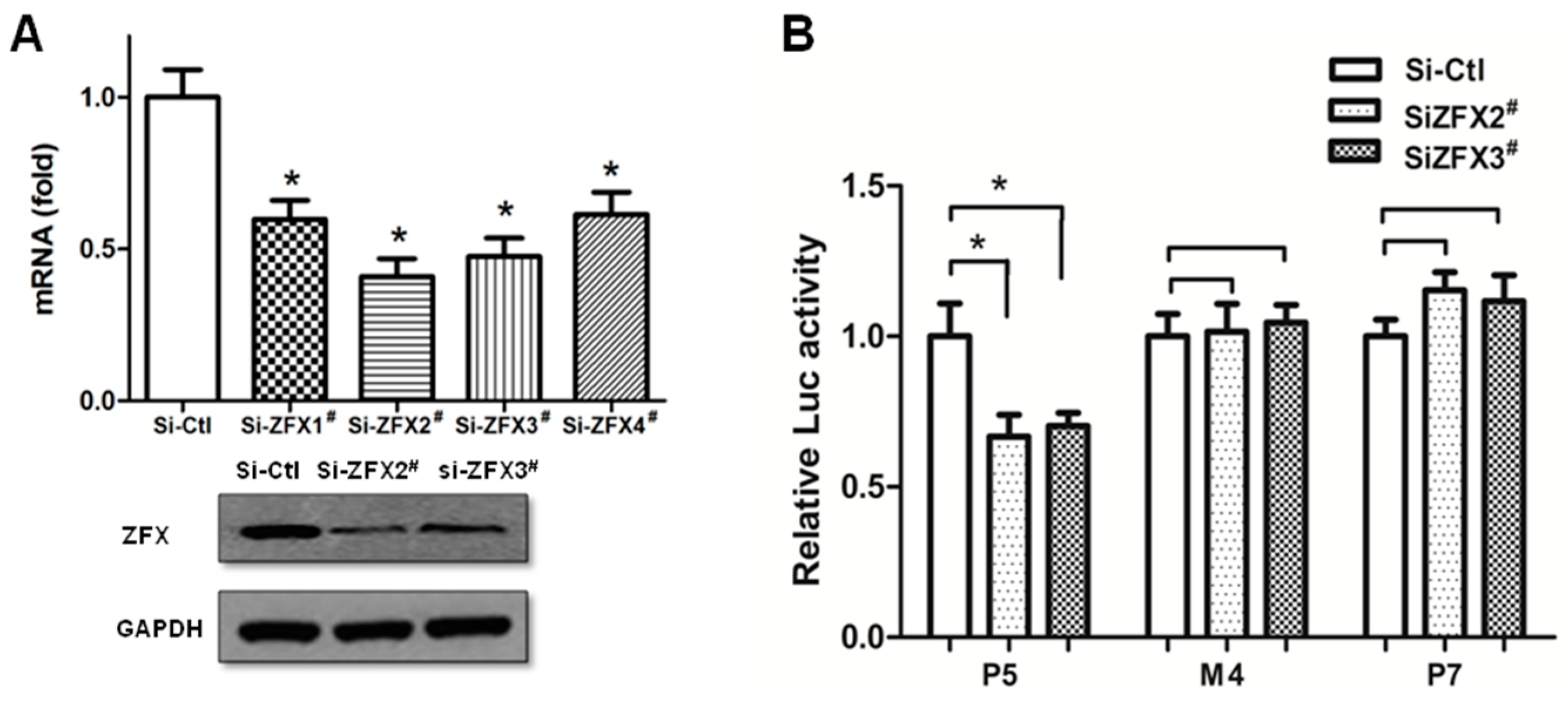
2.5. siRNA-Mediated Knockdown Verified the Significance of Site4
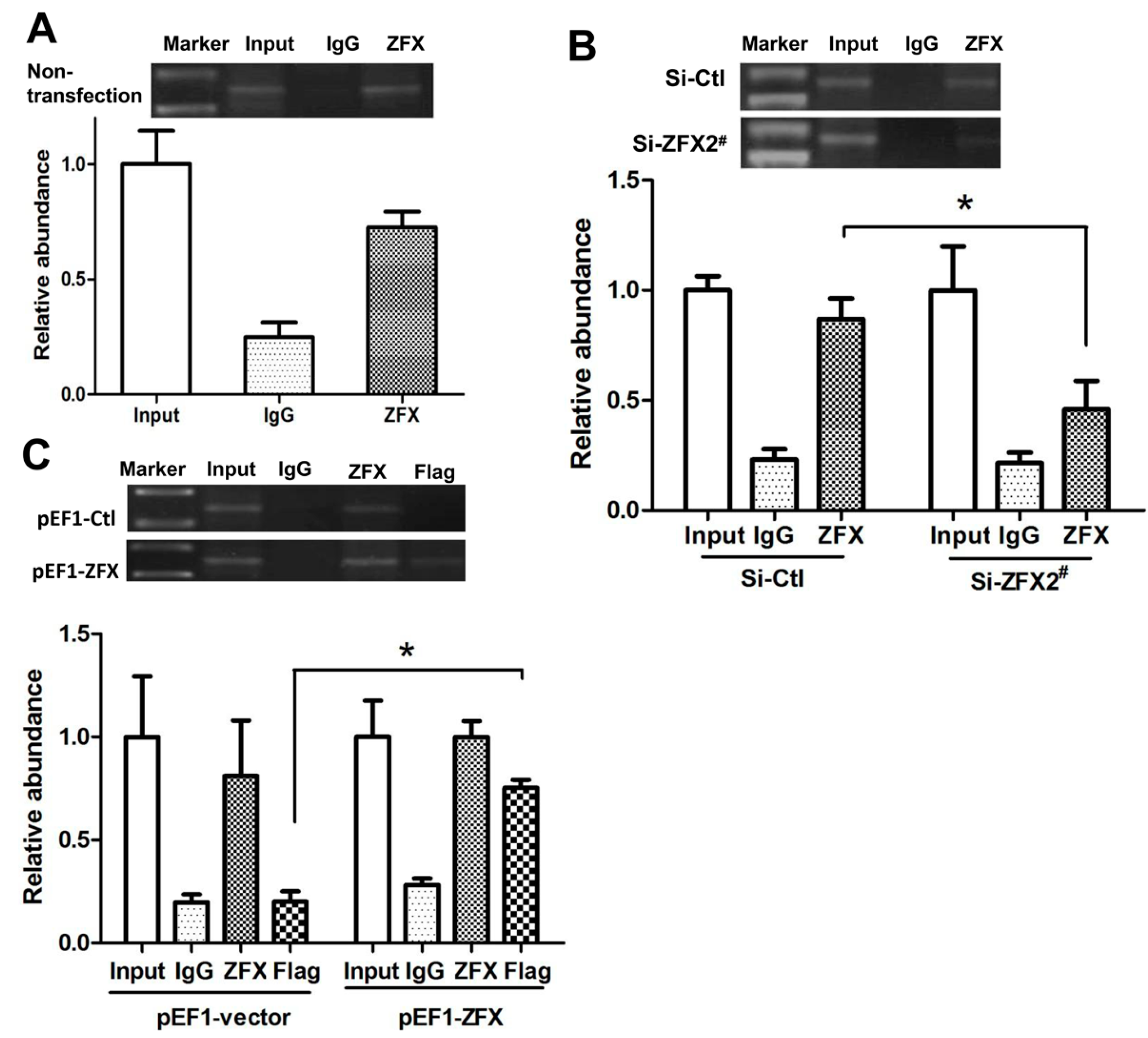
2.6. ZFX Binds to Native SET Gene Promoter
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmid Constructs and Small Interfering RNA (siRNA)
4.3. Transient Transfection and Luciferase Assay
4.4. Chromatin Immunoprecipitation (ChIP) Assay
4.5. RNA Extraction and Real-Time Quantitative-PCR
4.6. Western Blotting Analysis
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ZFX | Zinc finger and X-linked factor |
| PCOS | Polycystic Ovary Syndrome |
| TAF-1β | Template Activating Factor-1β |
| I2PP2A | protein phosphatase 2A inhibitor |
| AD | Alzheimer’s disease |
| ChIP | Chromatin immunoprecipitation |
| AML | Acute Myeloid Leukemia |
References
- Von Lindern, M.; van Baal, S.; Wiegant, J.; Raap, A.; Hagemeijer, A.; Grosveld, G. Can, a putative oncogene associated with myeloid leukemogenesis, may be activated by fusion of its 3′ half to different genes: Characterization of the set gene. Mol. Cell. Biol. 1992, 12, 3346–3355. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Saito, S.; Okuwaki, M.; Kawase, H.; Furuya, A.; Kusano, A.; Hanai, N.; Okuda, A.; Kikuchi, A. Cellular localization and expression of template-activating factor I in different cell types. Exp. Cell Res. 1998, 240, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Compagnone, N.A.; Fiore, C.; Vigne, J.L.; Culp, P.; Musci, T.J.; Mellon, S.H. Developmental gonadal expression of the transcription factor set and its target gene, p450c17 (17α-hydroxylase/c17,20 lyase). DNA Cell Biol. 2001, 20, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Chasseigneaux, S.; Clamagirand, C.; Huguet, L.; Gorisse-Hussonnois, L.; Rose, C.; Allinquant, B. Cytoplasmic set induces tau hyperphosphorylation through a decrease of methylated phosphatase 2a. BMC Neurosci. 2014, 15, 82. [Google Scholar] [CrossRef] [PubMed]
- Janghorban, M.; Farrell, A.S.; Allen-Petersen, B.L.; Pelz, C.; Daniel, C.J.; Oddo, J.; Langer, E.M.; Christensen, D.J.; Sears, R.C. Targeting C-MYC by antagonizing PP2A inhibitors in breast cancer. Proc. Natl. Acad Sci. USA 2014, 111, 9157–9162. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, X.F.; Ren, X.H.; Meng, X.J.; Huang, H.Y.; Zhao, Q.H.; Yuan, J.H.; Hong, W.X.; Xia, B.; Huang, X.F.; et al. Stable set knockdown in breast cell carcinoma inhibits cell migration and invasion. Biochem. Biophys. Res. Commun. 2014, 453, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Wei, J.; Zhang, Q.; Liu, F.; Basurto-Islas, G.; Grundke-Iqbal, I.; Iqbal, K. Cytoplasmic retention of protein phosphatase 2A inhibitor 2 (I2PP2A) induces alzheimer-like abnormal hyperphosphorylation of tau. J. Biol. Chem. 2014, 289, 27677–27691. [Google Scholar] [CrossRef] [PubMed]
- Bolognin, S.; Blanchard, J.; Wang, X.; Basurto-Islas, G.; Tung, Y.C.; Kohlbrenner, E.; Grundke-Iqbal, I.; Iqbal, K. An experimental rat model of sporadic Alzheimer’s disease and rescue of cognitive impairment with a neurotrophic peptide. Acta Neuropathol. 2012, 123, 133–151. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Liu, X.Q.; Xu, B.Q.; Jiang, S.W.; Cui, Y.G.; Liu, J.Y. SET/PP2A system regulates androgen production in ovarian follicles in vitro. Mol. Cell. Endocrinol. 2013, 374, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Gao, L.; Cui, Y.; Gao, L.; Dai, X.; Li, M.; Zhang, Y.; Ma, X.; Diao, F.; Liu, J. Set protein up-regulated testosterone production in the cultured preantral follicles. Reprod. Biol. Endocrinol. 2013, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Okuwaki, M.; Nagata, K. Template activating factor-I remodels the chromatin structure and stimulates transcription from the chromatin template. J. Biol. Chem. 1998, 273, 34511–34518. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Mellon, S.H. Multiple orphan nuclear receptors converge to regulate rat p450c17 gene transcription: Novel mechanisms for orphan nuclear receptor action. Mol. Endocrinol. 1997, 11, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.S.; Shin, F.Y.; Bae, S.C.; Kim, S.R.; Jeong, G.B.; Kwak, S.J.; Ballermann, B.J.; Kim, E.G. Expression of set is modulated as a function of cell proliferation. J. Cell Biochem. 1999, 74, 119–126. [Google Scholar] [CrossRef]
- Canela, N.; Rodriguez-Vilarrupla, A.; Estanyol, J.M.; Diaz, C.; Pujol, M.J.; Agell, N.; Bachs, O. The set protein regulates G2/M transition by modulating cyclin B-cyclin-dependent kinase 1 activity. J. Biol. Chem. 2003, 278, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Rincon, R.; Cristobal, I.; Zazo, S.; Arpi, O.; Menendez, S.; Manso, R.; Lluch, A.; Eroles, P.; Rovira, A.; Albanell, J.; et al. PP2A inhibition determines poor outcome and doxorubicin resistance in early breast cancer and its activation shows promising therapeutic effects. Oncotarget 2015, 6, 4299–4314. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, V.; Le Page, C.; Guyot, M.C.; Lussier, C.; Tonin, P.N.; Provencher, D.M.; Mes-Masson, A.M. Set complex in serous epithelial ovarian cancer. Int. J. Cancer 2006, 119, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhang, N.; Di, W. Regulation of LC3-dependent protective autophagy in ovarian cancer cells by protein phosphatase 2A. Int. J. Gynecol. Cancer 2013, 23, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Lam, B.D.; Anthony, E.C.; Hordijk, P.L. Cytoplasmic targeting of the proto-oncogene set promotes cell spreading and migration. FEBS Lett. 2013, 587, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Asaka, M.N.; Murano, K.; Nagata, K. Sp1-mediated transcription regulation of taf-ialpha gene encoding a histone chaperone. Biochem. Biophys. Res. Commun. 2008, 376, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.G.; Choi, M.E.; Ballermann, B.J. Spatially restricted expression of set mrna in developing rat kidney. Am. J. Physiol. 1994, 266, F155–F161. [Google Scholar] [PubMed]
- Pippa, R.; Dominguez, A.; Malumbres, R.; Endo, A.; Arriazu, E.; Marcotegui, N.; Guruceaga, E.; Odero, M.D. MYC-dependent recruitment of RUNX1 and GATA2 on the SET oncogene promoter enhances PP2A inactivation in acute myeloid leukemia. Oncotarget 2016. [Google Scholar] [CrossRef]
- Feng, Y.; Li, X.; Zhou, W.; Lou, D.; Huang, D.; Li, Y.; Kang, Y.; Xiang, Y.; Li, T.; Zhou, W.; et al. Regulation of SET gene expression by NFκB. Mol. Neurobiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.G.; Ren, W.; Zou, L.; Wang, Y.; Jin, R.; Zhou, G.P. Transcriptional control of human CD2AP expression: The role of sp1 and sp3. Mol. Biol. Rep. 2012, 39, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Galan-Caridad, J.M.; Harel, S.; Arenzana, T.L.; Hou, Z.E.; Doetsch, F.K.; Mirny, L.A.; Reizis, B. ZFX controls the self-renewal of embryonic and hematopoietic stem cells. Cell 2007, 129, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Gokhman, D.; Livyatan, I.; Sailaja, B.S.; Melcer, S.; Meshorer, E. Multilayered chromatin analysis reveals E2F, SMAD and ZFX as transcriptional regulators of histones. Nat. Struct. Mol. Biol. 2013, 20, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Cellot, S.; Sauvageau, G. ZFX: At the crossroads of survival and self-renewal. Cell 2007, 129, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.; Zhou, Q.; Wong, W.H. Chip-seq of transcription factors predicts absolute and differential gene expression in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2009, 106, 21521–21526. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Huang, Z.; Zhou, W.; Wu, Q.; Sloan, A.E.; Ouyang, G.; McLendon, R.E.; Yu, J.S.; Rich, J.N.; Bao, S. The zinc finger transcription factor ZFX is required for maintaining the tumorigenic potential of glioblastoma stem cells. Stem Cells 2014, 32, 2033–2047. [Google Scholar] [CrossRef] [PubMed]
- L'Haridon, M.; Paul, P.; Xerri, J.G.; Dastot, H.; Dolliger, C.; Schmid, M.; de Angelis, N.; Grollet, L.; Sigaux, F.; Degos, L.; et al. Transcriptional regulation of the MHC class I HLA-a11 promoter by the zinc finger protein ZFX. Nucleic Acids Res. 1996, 24, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Lu, Y.; Zheng, Y.B.; Yu, X.F.; Xia, X.H.; He, X.M.; Feng, W.L.; Xing, L.; Ling, Z.Q. ShRNA-mediated silencing of ZFX attenuated the proliferation of breast cancer cells. Cancer Chemoth. Pharm. 2014, 73, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Amini, S.; Fathi, F.; Mobalegi, J.; Sofimajidpour, H.; Ghadimi, T. The expressions of stem cell markers: OCT4, NANOG, SOX2, nucleostemin, BMI, ZFX, TCL1, TBX3, DPPA4, and ESRRB in bladder, colon, and prostate cancer, and certain cancer cell lines. Anat. Cell Biol. 2014, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Smith-Raska, M.R.; Esquilin, J.M.; Zhang, J.; Arenzana, T.L.; Lau, C.M.; Churchill, M.; Pan, H.; Klinakis, A.; Dixon, J.E.; et al. ZFX controls propagation and prevents differentiation of acute t-lymphoblastic and myeloid leukemia. Cell Rep. 2014, 6, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.P.; Chen, J.; He, M.; Ching, A.K.; Lau, C.; Lai, P.B.; To, K.F.; Wong, N. Overexpression of ZFX confers self-renewal and chemoresistance properties in hepatocellular carcinoma. Int. J. Cancer 2014, 135, 1790–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Zhu, Z.C.; Liu, Y.J.; Liu, J.W.; Wang, H.T.; Xiong, Z.Q.; Shen, X.; Hu, Z.L.; Zheng, J. ZFX knockdown inhibits growth and migration of non-small cell lung carcinoma cell line H1299. Int. J. Clin. Exp. Pathol. 2013, 6, 2460–2467. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Primer | Sequence | Size (bp) |
|---|---|---|---|
| P1 (pGL3-855) | F1 | TCTTACGCGTGCTAGCAGTAGAACTTGTTGGCCCCT | 994 |
| P2 (pGL3-654) | F2 | TCTTACGCGTGCTAGCCTTCAGTGTCGTAACCTTTA | 794 |
| P3 (pGL3-327) | F3 | TCTTACGCGTGCTAGCGCAGGGAAGCCGCTCGCTCA | 467 |
| P4 (pGL3-251) | F4 | TCTTACGCGTGCTAGCGCCTGCCGCGCGCCAGTGTC | 391 |
| P5 (pGL3-157) | F5 | TCTTACGCGTGCTAGCTCCTCCCACCTCTCGCCGGG | 297 |
| P6 (pGL3-85) | F6 | TCTTACGCGTGCTAGCTGGCGCCGCGGCGCGAGCCT | 225 |
| P7 (pGL3-45) | F7 | TCTTACGCGTGCTAGCTCCGCGCGGGGCCTGGCGCGCCT | 185 |
| P8 (pGL3+47) | F8 | TCTTACGCGTGCTAGCGCTGGCTGGATCGCCGAGCG | 93 |
| P1–P8 | R | GATCGCAGATCTCGAGGCCCCGGCCCGGGCTCCTGT | |
| M1 | Fr1 | TCTTACGCGTGCTAGCTCCTCCCACCTAACGCTTAGACATCTGTCTCCGCGCCGCGACGAT | 329 |
| Re1 | CGCAGATCTCGAGGCCCCGGCCCGGGCTC | ||
| M2 | Fr2 | TCTTACGCGTGCTAGCTCCTCCCACCTCTC | 101 |
| Re2 | CGCCGCGGCGCCAGATGTCTAAGCGTTGGGATGAGGGGGCGGAGTC | ||
| Fr2′ | AGACATCTGGCGCCGCGGCGCGAGCCTC | 249 | |
| Re2′ | CCGCGACGATCGCAGATCTCGAGGCCCCGGCCCGGGCTC | ||
| M3 | Fr3 | TCTTACGCGTGCTAGCTCCTCCCACCTCTC | 119 |
| Re3 | TGATGTCTAAGCGTTTCGCGCCGCGGCGCCATTGGC | ||
| Fr3′ | CGCGAAACGCTTAGACATCAGGCGGAGCATCCGCGCG | 230 | |
| Re3′ | CCGCGACGATCGCAGATCTCGAGGCCCCGGCCCGGGCTC | ||
| M4 | Fr4 | TCTTACGCGTGCTAGCTCCTCCCACCTCTC | 146 |
| Re4 | CGCGATGTCTAAGCGTTATGCTCCGCCTCCGGCC | ||
| Fr4′ | CATAACGCTTAGACATCGCGCGCCTGCGCCCTGCG | 203 | |
| Re4′ | CCGCGACGATCGCAGATCTCGAGGCCCCGGCCCGGGCTC | ||
| ChIP-promoter | F-pr | TCTCGCCGGGCCCTGTGTCT | 173 |
| R-pr | CGCCTCCTCCTCCACCTCCT | ||
| ChIP-plasmid | F′-pd | CGCCCAGGCCAATGG | 322 |
| R′-pd | TTGGCGTCTTCCATGGTG | ||
| ZFX | P1 | TTGCTGAAATCGCTGACGAAG | 133 |
| P2 | GCAATCGGCATGAAGGTTTTGAT | ||
| GAPDH | P3 | ACCATCTTCCAGGAGCGAGA | 71 |
| P4 | GACTCCACGACGTACTCAGC |
| Designation | Target Sequence |
|---|---|
| si-ZFX1# | UGAAAUCGCUGACGAAUGG |
| si-ZFX2# | AAGCAGAAAUUGUCACUGA |
| si-ZFX3# | GGACGUUGUUAUAGAAGAU |
| si-ZFX4# | ACACAGAGUCGGAAAUUGA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Duan, P.; Li, J.; Senkowski, T.; Guo, F.; Chen, H.; Romero, A.; Cui, Y.; Liu, J.; Jiang, S.-W. Zinc Finger and X-Linked Factor (ZFX) Binds to Human SET Transcript 2 Promoter and Transactivates SET Expression. Int. J. Mol. Sci. 2016, 17, 1737. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101737
Xu S, Duan P, Li J, Senkowski T, Guo F, Chen H, Romero A, Cui Y, Liu J, Jiang S-W. Zinc Finger and X-Linked Factor (ZFX) Binds to Human SET Transcript 2 Promoter and Transactivates SET Expression. International Journal of Molecular Sciences. 2016; 17(10):1737. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101737
Chicago/Turabian StyleXu, Siliang, Ping Duan, Jinping Li, Tristan Senkowski, Fengbiao Guo, Haibin Chen, Alberto Romero, Yugui Cui, Jiayin Liu, and Shi-Wen Jiang. 2016. "Zinc Finger and X-Linked Factor (ZFX) Binds to Human SET Transcript 2 Promoter and Transactivates SET Expression" International Journal of Molecular Sciences 17, no. 10: 1737. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101737