
Altered Episodic Memory in Introverted Young Adults Carrying the BDNFMet Allele

Abstract
:
1. Introduction
2. Results
2.1. Univariate Analyses of Variance
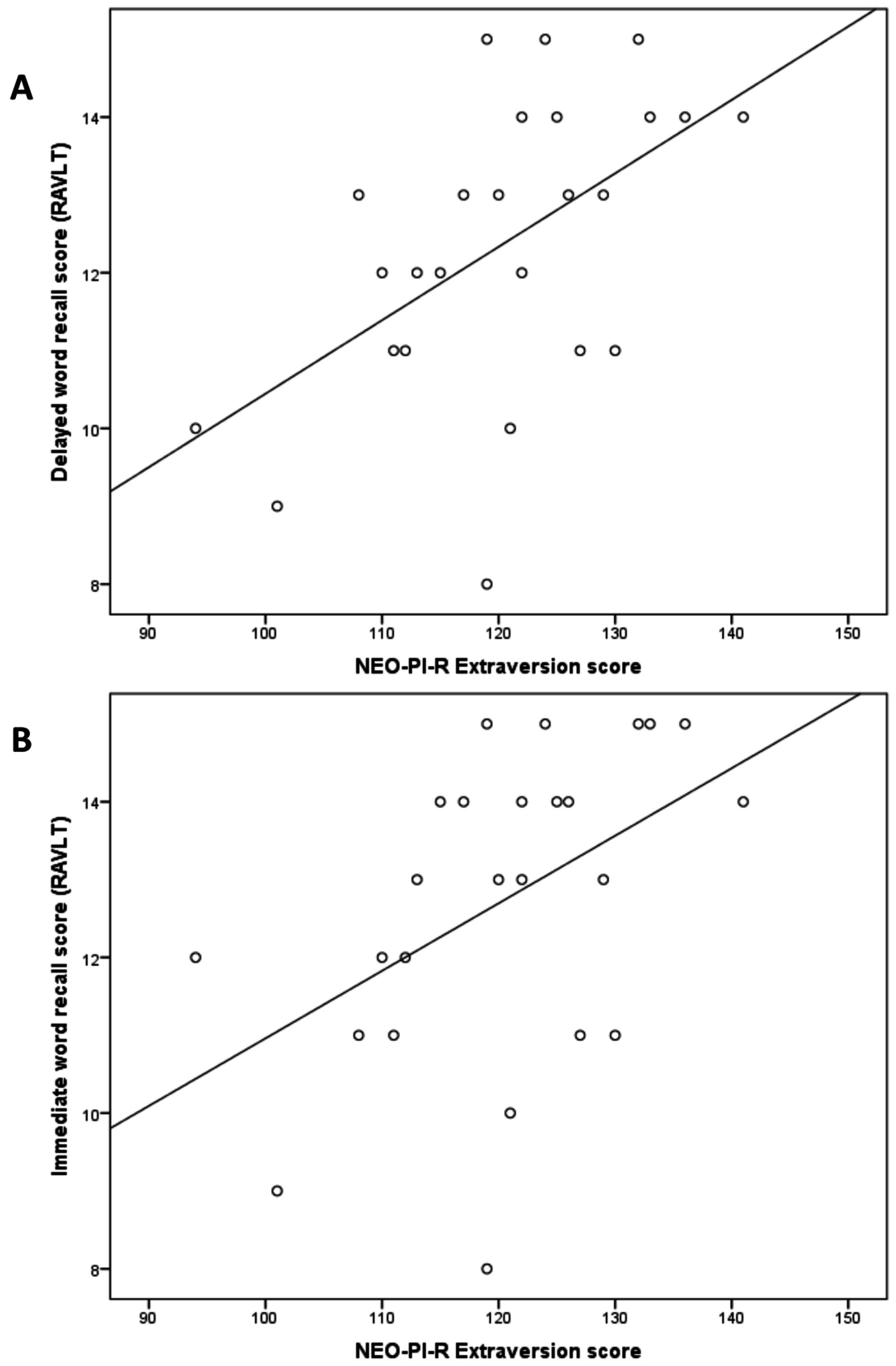
2.2. Regression Analyses
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Procedure
4.2.1. Neuropsychological Testing
4.2.2. Questionnaires
4.2.3. Genotyping
4.3. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Poo, M.M. Neurotrophins as synaptic modulators. Nat. Rev. Neurosci. 2001, 2, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tyler, W.J.; Alonso, M.; Bramham, C.R.; Pozzo-Miller, L.D. From acquisition to consolidation: On the role of brain-derived neurotrophic factor signaling in hippocampal-dependent learning. Learn. Mem. 2002, 9, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Bramham, C.R.; Messaoudi, E. BDNF function in adult synaptic plasticity: The synaptic consolidation hypothesis. Prog. Neurobiol. 2005, 76, 99–125. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.; Brene, S.; Mathe, A. BDNF in schizophrenia, depression and corresponding animal models. Mol. Psychiatry 2005, 10, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of brain-derived neurotrophic factor (BDNF) protein and mRNA in the normal adult rat CNS: Evidence for anterograde axonal transport. J. Neurosci. 1997, 17, 2295–2313. [Google Scholar] [PubMed]
- Figurov, A.; Pozzo-Miller, L.D.; Olafsson, P.; Wang, T.; Lu, B. Regulation of synaptic responses to high-frequency stimulation and LTP by neurotrophins in the hippocampus. Nature 1996, 381, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Patel, P.D.; Sant, G.; Meng, C.X.; Teng, K.K.; Hempstead, B.L.; Lee, F.S. Variant brain-derived neurotrophic factor (BDNF)(Met66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. J. Neurosci. 2004, 24, 4401–4411. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.J.; Hong, C.J.; Yu, Y.Y.; Chen, T.J. Association study of a brain-derived neurotrophic factor (BDNF) Val66Met polymorphism and personality trait and intelligence in healthy young females. Neuropsychobiology 2004, 49, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Rybakowski, J.K.; Borkowska, A.; Czerski, P.M.; Skibińska, M.; Hauser, J. Polymorphism of the brain-derived neurotrophic factor gene and performance on a cognitive prefrontal test in bipolar patients. Bipolar Disord. 2003, 5, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Rybakowski, J.K.; Borkowska, A.; Skibinska, M.; Szczepankiewicz, A.; Kapelski, P.; Leszczynska-Rodziewicz, A.; Czerski, P.M.; Hauser, J. Prefrontal cognition in schizophrenia and bipolar illness in relation to Val66Met polymorphism of the brain-derived neurotrophic factor gene. Psychiatry Clin. Neurosci. 2006, 60, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Raz, N.; Rodrigue, K.M.; Kennedy, K.M.; Land, S. Genetic and vascular modifiers of age-sensitive cognitive skills: Effects of COMT, BDNF, ApoE, and hypertension. Neuropsychology 2009, 23, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Dempster, E.; Toulopoulou, T.; McDonald, C.; Bramon, E.; Walshe, M.; Filbey, F.; Wickham, H.; Sham, P.C.; Murray, R.M.; Collier, D.A. Association between BDNF Val66Met genotype and episodic memory. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 134, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M. The BDNF Val66Met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Hariri, A.R.; Goldberg, T.E.; Mattay, V.S.; Kolachana, B.S.; Callicott, J.H.; Egan, M.F.; Weinberger, D.R. Brain-derived neurotrophic factor Val66Met polymorphism affects human memory-related hippocampal activity and predicts memory performance. J. Neurosci. 2003, 23, 6690–6694. [Google Scholar] [PubMed]
- Tan, H.Y.; Chen, Q.; Sust, S.; Buckholtz, J.W.; Meyers, J.D.; Egan, M.F.; Mattay, V.S.; Meyer-Lindenberg, A.; Weinberger, D.R.; Callicott, J.H. Epistasis between catechol-o-methyltransferase and type II metabotropic glutamate receptor 3 genes on working memory brain function. Proc. Natl. Acad. Sci. USA 2007, 104, 12536–12541. [Google Scholar] [CrossRef] [PubMed]
- Pezawas, L.; Verchinski, B.A.; Mattay, V.S.; Callicott, J.H.; Kolachana, B.S.; Straub, R.E.; Egan, M.F.; Meyer-Lindenberg, A.; Weinberger, D.R. The brain-derived neurotrophic factor Val66Met polymorphism and variation in human cortical morphology. J. Neurosci. 2004, 24, 10099–10102. [Google Scholar] [CrossRef] [PubMed]
- Montag, C.; Weber, B.; Fliessbach, K.; Elger, C.; Reuter, M. The BDNF Val66Met polymorphism impacts parahippocampal and amygdala volume in healthy humans: Incremental support for a genetic risk factor for depression. Psychol. Med. 2009, 39, 1831–1839. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Walss-Bass, C.; Nery, F.G.; Nicoletti, M.A.; Hatch, J.P.; Frey, B.N.; Monkul, E.S.; Zunta-Soares, G.B.; Bowden, C.L.; Escamilla, M.A. Neuronal correlates of brain-derived neurotrophic factor Val66Met polymorphism and morphometric abnormalities in bipolar disorder. Neuropsychopharmacology 2009, 34, 1904–1913. [Google Scholar] [CrossRef] [PubMed]
- Kambeitz, J.P.; Bhattacharyya, S.; Kambeitz-Ilankovic, L.M.; Valli, I.; Collier, D.A.; McGuire, P. Effect of BDNF Val66Met polymorphism on declarative memory and its neural substrate: A meta-analysis. Neurosci. Biobehav. Rev. 2012, 36, 2165–2177. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Fagan, A.M.; Goate, A.M.; Benzinger, T.L.; Morris, J.C.; Head, D. Lack of an association of BDNF Val66Met polymorphism and plasma BDNF with hippocampal volume and memory. Cogn. Affect. Behav. Neurosci. 2015, 15, 625–643. [Google Scholar] [CrossRef] [PubMed]
- Tulving, E. Episodic and semantic memory 1. Organ. Memory Lond. Acad. 1972, 381, 382–404. [Google Scholar]
- Shimizu, E.; Hashimoto, K.; Iyo, M. Ethnic difference of the BDNF 196G/A (Val66Met) polymorphism frequencies: The possibility to explain ethnic mental traits. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2004, 126, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Cheeran, B.; Talelli, P.; Mori, F.; Koch, G.; Suppa, A.; Edwards, M.; Houlden, H.; Bhatia, K.; Greenwood, R.; Rothwell, J.C. A common polymorphism in the brain-derived neurotrophic factor gene (BDNF) modulates human cortical plasticity and the response to rTMS. J. Physiol. 2008, 586, 5717–5725. [Google Scholar] [CrossRef] [PubMed]
- McCrae, R.R.; Allik, I.U. The Five-Factor Model of Personality Across Cultures; Marsella, A.J., Ed.; Springer Science & Business Media: New York, NY, USA, 2002; Volume 1, pp. 1–4. [Google Scholar]
- McCrae, R.R.; Costa, P.T. Validation of the five-factor model of personality across instruments and observers. J. Personal. Soc. Psychol. 1987, 52, 81–90. [Google Scholar] [CrossRef]
- McCrae, R.R.; Costa, P.T., Jr. Personality in Adulthood: A Five-Factor Theory Perspective; Guilford Press: New York, NY, USA, 2003; Volume 1, pp. 1–268. [Google Scholar]
- McCrae, R.R.; Costa, P.T., Jr. Personality trait structure as a human universal. Am. Psychol. 1997, 52. [Google Scholar] [CrossRef]
- Rawlings, D.; Carnie, D. The interaction of EPQ extraversion with WAIS subtest performance under timed and untimed conditions. Personal. Individ. Differ. 1989, 10, 453–458. [Google Scholar] [CrossRef]
- Baker, T.J.; Bichsel, J. Personality predictors of intelligence: Differences between young and cognitively healthy older adults. Personal. Individ. Differ. 2006, 41, 861–871. [Google Scholar] [CrossRef]
- Chamorro-Premuzic, T.; Furnham, A.; Petrides, K. Personality and intelligence. J. Individ. Differ. 2006, 27, 147–150. [Google Scholar] [CrossRef]
- Crowe, M.; Andel, R.; Pedersen, N.L.; Fratiglioni, L.; Gatz, M. Personality and risk of cognitive impairment 25 years later. Psychol. Aging 2006, 21, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-Premuzic, T.; Furnham, A. A possible model for understanding the personality-intelligence interface. Br. J. Psychol. 2004, 95, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-Premuzic, T.; Furhnam, A. Intellectual competence and the intelligent personality: A third way in differential psychology. Rev. Gen. Psychol. 2006, 10, 251–267. [Google Scholar] [CrossRef]
- Allen, P.A.; Kaut, K.; Baena, E.; Lien, M.C.; Ruthruff, E. Individual differences in positive affect moderate age-related declines in episodic long-term memory. J. Cogn. Psychol. 2011, 23, 768–779. [Google Scholar] [CrossRef]
- Meier, B.; Perrig-Chiello, P.; Perrig, W. Personality and memory in old age. Aging Neuropsychol. Cogn. 2002, 9, 135–144. [Google Scholar] [CrossRef]
- Mitchell, D.B. How many memory systems? Evidence from aging. J. Exp. Psychol. Learn. Mem. Cogn. 1989, 15, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Light, L.L. The organization of memory in old age. In The Handbook of Aging and Cognition; Craik, F.I.M., Salthouse, T.A., Eds.; Psychology Press: New York, NY, USA, 1992; Volume 1, pp. 111–165. [Google Scholar]
- Allen, P.A.; Sliwinski, M.; Bowie, T.; Madden, D.J. Differential age effects in semantic and episodic memory. J. Gerontol. B Psychol. Sci. Soc. Sci. 2002, 57, 173–186. [Google Scholar] [CrossRef]
- Matthews, G. Human Performance: Cognition, Stress, and Individual Differences; Psychology Press: New York, NY, USA, 2000; Volume 1, pp. 265–285. [Google Scholar]
- Terracciano, A.; Sanna, S.; Uda, M.; Deiana, B.; Usala, G.; Busonero, F.; Maschio, A.; Scally, M.; Patriciu, N.; Chen, W.M. Genome-wide association scan for five major dimensions of personality. Mol. Psychiatry 2010, 15, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, A.; Tanaka, T.; Sutin, A.R.; Deiana, B.; Balaci, L.; Sanna, S.; Olla, N.; Maschio, A.; Uda, M.; Ferrucci, L. BDNF Val66Met is associated with introversion and interacts with 5-HTTLPR to influence neuroticism. Neuropsychopharmacology 2010, 35, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- De Beaumont, L.; Fiocco, A.J.; Quesnel, G.; Lupien, S.; Poirier, J. Altered declarative memory in introverted middle-aged adults carrying the BDNF Val66Met allele. Behav. Brain Res. 2013, 253, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Richter-Schmidinger, T.; Alexopoulos, P.; Horn, M.; Maus, S.; Reichel, M.; Rhein, C.; Lewczuk, P.; Sidiropoulos, C.; Kneib, T.; Perneczky, R. Influence of brain-derived neurotrophic-factor and apolipoprotein E genetic variants on hippocampal volume and memory performance in healthy young adults. J. Neural Transm. 2011, 118, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Kolb, B.; Muhammad, A.; Gibb, R. Searching for factors underlying cerebral plasticity in the normal and injured brain. J. Commun. Disord. 2011, 44, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Lommatzsch, M.; Zingler, D.; Schuhbaeck, K.; Schloetcke, K.; Zingler, C.; Schuff-Werner, P.; Virchow, J.C. The impact of age, weight and gender on BDNF levels in human platelets and plasma. Neurobiol. Aging 2005, 26, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Ziegenhorn, A.A.; Schulte-Herbrüggen, O.; Danker-Hopfe, H.; Malbranc, M.; Hartung, H.D.; Anders, D.; Lang, U.E.; Steinhagen-Thiessen, E.; Schaub, R.T.; Hellweg, R. Serum neurotrophins—A study on the time course and influencing factors in a large old age sample. Neurobiol. Aging 2007, 28, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Donnellan, M.B.; Lucas, R.E. Age differences in the Big Five across the life span: Evidence from two national samples. Psychol. Aging 2008, 23, 558–566. [Google Scholar] [CrossRef] [PubMed]
- McCrae, R.R.; Costa, P.T.; de Lima, M.P.; Simões, A.; Ostendorf, F.; Angleitner, A.; Marušić, I.; Bratko, D.; Caprara, G.V.; Barbaranelli, C. Age differences in personality across the adult life span: parallels in five cultures. Dev. Psychol. 1999, 35, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Colzato, L.S.; van der Does, A.W.; Kouwenhoven, C.; Elzinga, B.M.; Hommel, B. BDNF Val66Met polymorphism is associated with higher anticipatory cortisol stress response, anxiety, and alcohol consumption in healthy adults. Psychoneuroendocrinology 2011, 36, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.T.; McCrae, R.R. Influence of extraversion and neuroticism on subjective well-being: Happy and unhappy people. J. Personal. Soc. Psychol. 1980, 38, 668–678. [Google Scholar] [CrossRef]
- Xu, L.; Anwyl, R.; Rowan, M.J. Behavioural stress facilitates the induction of long-term depression in the hippocampus. Nature 1997, 387, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Foy, M.R.; Thompson, R.F. Behavioral stress modifies hippocampal plasticity through N-methyl-d-aspartate receptor activation. Proc. Natl. Acad. Sci. USA 1996, 93, 4750–4753. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Yang, C.H.; Hsu, K.S. Do stress and long-term potentiation share the same molecular mechanisms? Mol. Neurobiol. 2005, 32, 223–235. [Google Scholar] [CrossRef]
- Yu, H.; Wang, D.D.; Wang, Y.; Liu, T.; Lee, F.S.; Chen, Z.Y. Variant brain-derived neurotrophic factor Val66Met polymorphism alters vulnerability to stress and response to antidepressants. J. Neurosci. 2012, 32, 4092–4101. [Google Scholar] [CrossRef] [PubMed]
- Ninan, I.; Bath, K.G.; Dagar, K.; Perez-Castro, R.; Plummer, M.R.; Lee, F.S.; Chao, M.V. The BDNF Val66Met polymorphism impairs NMDA receptor-dependent synaptic plasticity in the hippocampus. J. Neurosci. 2010, 30, 8866–8870. [Google Scholar] [CrossRef] [PubMed]
- Groves, J. Is it time to reassess the BDNF hypothesis of depression? Mol. Psychiatry 2007, 12, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Jing, D.; Bath, K.G.; Ieraci, A.; Khan, T.; Siao, C.J.; Herrera, D.G.; Toth, M.; Yang, C.; McEwen, B.S. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 2006, 314, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.D.; Züchner, S.; McQuoid, D.R.; Steffens, D.C.; Blazer, D.G.; Krishnan, K.R.R. Social support in older individuals: The role of the BDNF Val66Met polymorphism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.R.; Rench, T.A.; Lyons, J.B.; Riffle, R.R. The influence of neuroticism, extraversion and openness on stress responses. Stress Health 2012, 28, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Suls, J. Affect, Stress, and Personality. In Handbook of Affect and Social Cognition; Lawrence Erlbaum Associates Publishers: Mahwah, NJ, USA, 2001; Volume 1, pp. 392–409. [Google Scholar]
- Swickert, R.J.; Rosentreter, C.J.; Hittner, J.B.; Mushrush, J.E. Extraversion, social support processes, and stress. Personal. Individ. Differ. 2002, 32, 877–891. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A. Extraversion and Its Positive Emotional Core. In Handbook of Personality Psychology; Academic Press: San Diego, CA, USA, 1997; Volume 1, pp. 767–793. [Google Scholar]
- Amirkhan, J.H.; Risinger, R.T.; Swickert, R.J. Extraversion: A “hidden” personality factor in coping? J. Personal. 1995, 63, 189–212. [Google Scholar] [CrossRef]
- David, J.P.; Suls, J. Coping efforts in daily life: Role of Big Five traits and problem appraisals. J. Personal. 1999, 67, 265–294. [Google Scholar] [CrossRef]
- Watson, D.; Hubbard, B. Adaptational style and dispositional structure: Coping in the context of the five-factor model. J. Personal. 1996, 64, 737–774. [Google Scholar] [CrossRef]
- Pavot, W.; Diener, E.; Fujita, F. Extraversion and happiness. Personal. Individ. Differ. 1990, 11, 1299–1306. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A. On traits and temperament: General and specific factors of emotional experience and their relation to the five-factor model. J. Personal. 1992, 60, 441–476. [Google Scholar] [CrossRef]
- Rusting, C.L.; Larsen, R.J. Extraversion, neuroticism, and susceptibility to positive and negative affect: A test of two theoretical models. Personal. Individ. Differ. 1997, 22, 607–612. [Google Scholar] [CrossRef]
- Jylhä, P.; Melartin, T.; Rytsälä, H.; Isometsä, E. Neuroticism, introversion, and major depressive disorder—Traits, states, or scars? Depress. Anxiety 2009, 26, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Craik, F.I.; Lockhart, R.S. Levels of processing: A framework for memory research. J. Verbal Learn. Verbal Behav. 1972, 11, 671–684. [Google Scholar] [CrossRef]
- Arbuckle, T.Y.; Gold, D.P.; Andres, D.; Schwartzman, A.; Chaikelson, J. The role of psychosocial context, age, and intelligence in memory performance of older men. Psychol. Aging 1992, 7, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Lezak, M.D. Neuropsychological Assessment, 3rd ed.; Oxford University Press: New York, NY, USA, 1995; pp. 346–352. [Google Scholar]
- Strauss, E.; Sherman, E.M.; Spreen, O. A Compendium of Neuropsychological Tests: Administration, Norms, and Commentary; Oxford University Press: New York, NY, USA, 2006; pp. 263–266. [Google Scholar]
- Fichman, H.C.; Teresa Dias, L.B.; Fernandes, C.S.; Lourenço, R.; Caramelli, P.; Nitrini, R. Normative data and construct validity of the Rey Auditory Verbal Learning Test in a Brazilian elderly population. Psychol. Neurosci. 2010, 3, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. Rey Auditory Verbal Learning Test: A Handbook; Western Psychological Services: Los Angeles, CA, USA, 1996; Volume 1, pp. 6–27. [Google Scholar]
- Costa, P.T.; McCrae, R.R. Revised Neo Personality Inventory (NEO-PI-R) and NEO Five-Factor Inventory (NEO-FFI); Psychological Assesment Resources: Lutz, FL, USA, 1992; Volume 1, pp. 36–39. [Google Scholar]
- McCrae, R.R.; Terracciano, A. Universal features of personality traits from the observer’s perspective: Data from 50 cultures. J. Personal. Soc. Psychol. 2005, 88, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Royo, J.L.; Hidalgo, M.; Ruiz, A. Pyrosequencing protocol using a universal biotinylated primer for mutation detection and SNP genotyping. Nat. Protoc. 2007, 2, 1734–1739. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Variables of Interest | BDNF Genotype | F | p | |
|---|---|---|---|---|
| BDNFMet (n = 25) | BDNFVal (n = 49) | |||
| Age (year) | 20.96 (1.94) | 20.58 (1.66) | 0.79 | 0.38 |
| Education (year) | 14.81 (1.72) | 14.16 (1.53) | 2.818 | 0.09 |
| BAI | 5.03 (4.85) | 4.79 (4.25) | 0.05 | 0.83 |
| BDI | 3.07 (2.76) | 3.73 (2.97) | 0.88 | 0.35 |
| Personality NEO-PI-R | ||||
| Neuroticism | 81.76 (17.14) | 80.21 (21.18) | 0.103 | 0.75 |
| Extraversion | 120.28 (10.87) | 131.05 (12.44) | 9.54 | 0.01 * |
| Openness | 101.04 (13.79) | 105.79 (16.31) | 1.60 | 0.21 |
| Agreeableness | 120.63 (18.21) | 121.78 (12.49) | 0.10 | 0.75 |
| Conscientiousness | 119.11 (24.19) | 118.52 (16.37) | 0.02 | 0.90 |
| RAVLT | ||||
| Word Immediate recall | 12.19 (2.77) | 11.81 (2.02) | 0.46 | 0.50 |
| Word Delayed recall | 12.06 (2.30) | 11.38 (2.56) | 1.28 | 0.26 |
| RCFT | ||||
| Immediate recall | 23.06 (6.16) | 24.03 (5.21) | 0.51 | 0.47 |
| Delayed recall | 22.92 (6.05) | 24.07 (5.27) | 0.73 | 0.40 |
| D-KEFS Verbal Fluency Test | ||||
| Condition 3 Total switching accuracy | 13.47 (2.19) | 12.71 (2.69) | 1.53 | 0.22 |
| Set-Loss Errors | 0.74 (0.85) | 0.89 (1.07) | 0.38 | 0.54 |
| D-KEFS Trail Making Test | ||||
| Condition 4 Total Time | 50.62 (12.01) | 50.78 (14.23) | 0.002 | 0.96 |
| Condition 4 Total Errors | 0.61 (0.75) | 0.64 (0.81) | 0.024 | 0.87 |
| D-KEFS Color-Word Interference Test | ||||
| Condition 4 Total Time | 49.71 (8.60) | 49.97 (12.45) | 0.009 | 0.93 |
| Condition 4 Total Errors | 1.06 (0.72) | 1.31 (1.29) | 0.84 | 0.36 |
| BDNFMet | R | p | n |
| RAVLT | |||
| Word Immediate recall | 0.479 | 0.015 * | 25 |
| Word Delayed recall | 0.542 | 0.005 * | 25 |
| RCFT | |||
| Immediate recall | 0.151 | 0.46 | 25 |
| Delayed recall | 0.182 | 0.37 | 25 |
| D-KEFS Verbal Fluency Test | |||
| Condition 3 Total switching accuracy | 0.256 | 0.21 | 25 |
| Set-Loss Errors | 0.249 | 0.22 | 25 |
| D-KEFS Trail Making Test | |||
| Condition 4 Total Time | 0.321 | 0.11 | 25 |
| Condition 4 Total Errors | −0.014 | 0.94 | 25 |
| D-KEFS Color-Word Interference Test | |||
| Condition 4 Total Time | −0.131 | 0.52 | 25 |
| Condition 4 Total Errors | −0.024 | 0.91 | 25 |
| BDNFVal | R | p | n |
| RAVLT | |||
| Word Immediate recall | −0.035 | 0.81 | 49 |
| Word Delayed recall | 0.125 | 0.39 | 49 |
| RCFT | |||
| Immediate recall | 0.068 | 0.64 | 49 |
| Delayed recall | 0.050 | 0.73 | 49 |
| D-KEFS Verbal Fluency Test | |||
| Condition 3 Total switching accuracy | 0.172 | 0.24 | 49 |
| Set-Loss Errors | 0.085 | 0.56 | 49 |
| D-KEFS Trail Making Test | |||
| Condition 4 Total Time | −0.201 | 0.17 | 49 |
| Condition 4 Total Errors | −0.053 | 0.72 | 49 |
| D-KEFS Color-Word Interference Test | |||
| Condition 4 Total Time | 0.016 | 0.91 | 49 |
| Condition 4 Total Errors | −0.216 | 0.14 | 49 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bombardier, A.; Beauchemin, M.; Gosselin, N.; Poirier, J.; De Beaumont, L. Altered Episodic Memory in Introverted Young Adults Carrying the BDNFMet Allele. Int. J. Mol. Sci. 2016, 17, 1886. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17111886
Bombardier A, Beauchemin M, Gosselin N, Poirier J, De Beaumont L. Altered Episodic Memory in Introverted Young Adults Carrying the BDNFMet Allele. International Journal of Molecular Sciences. 2016; 17(11):1886. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17111886
Chicago/Turabian StyleBombardier, Andreanne, Maude Beauchemin, Nadia Gosselin, Judes Poirier, and Louis De Beaumont. 2016. "Altered Episodic Memory in Introverted Young Adults Carrying the BDNFMet Allele" International Journal of Molecular Sciences 17, no. 11: 1886. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17111886