
Characterization of Starch Degradation Related Genes in Postharvest Kiwifruit

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
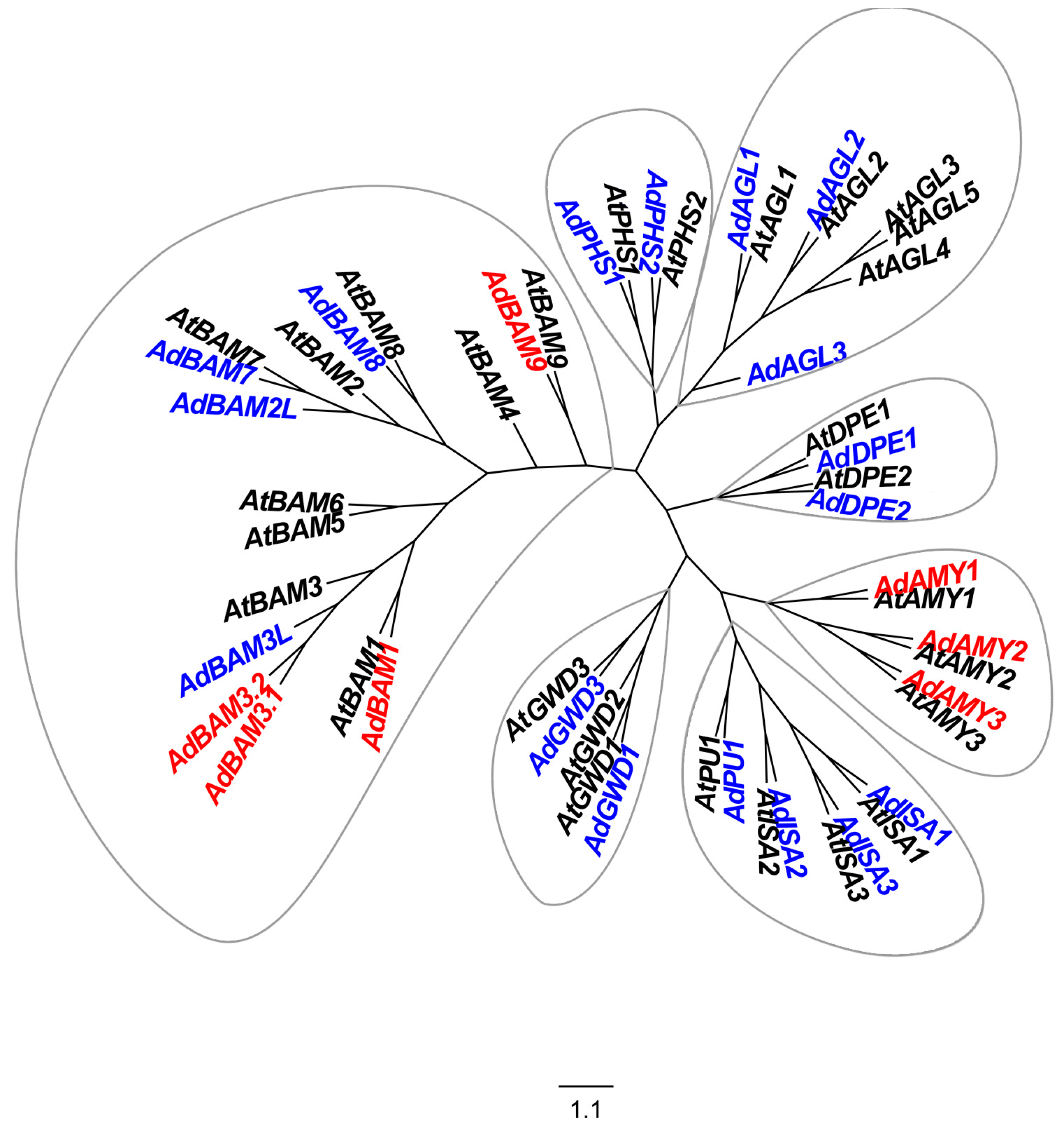
2.1. Isolation and Analysis of Starch-Related Genes
2.2. Starch Degradation during Kiwifruit Ripening and Softening at 20 °C
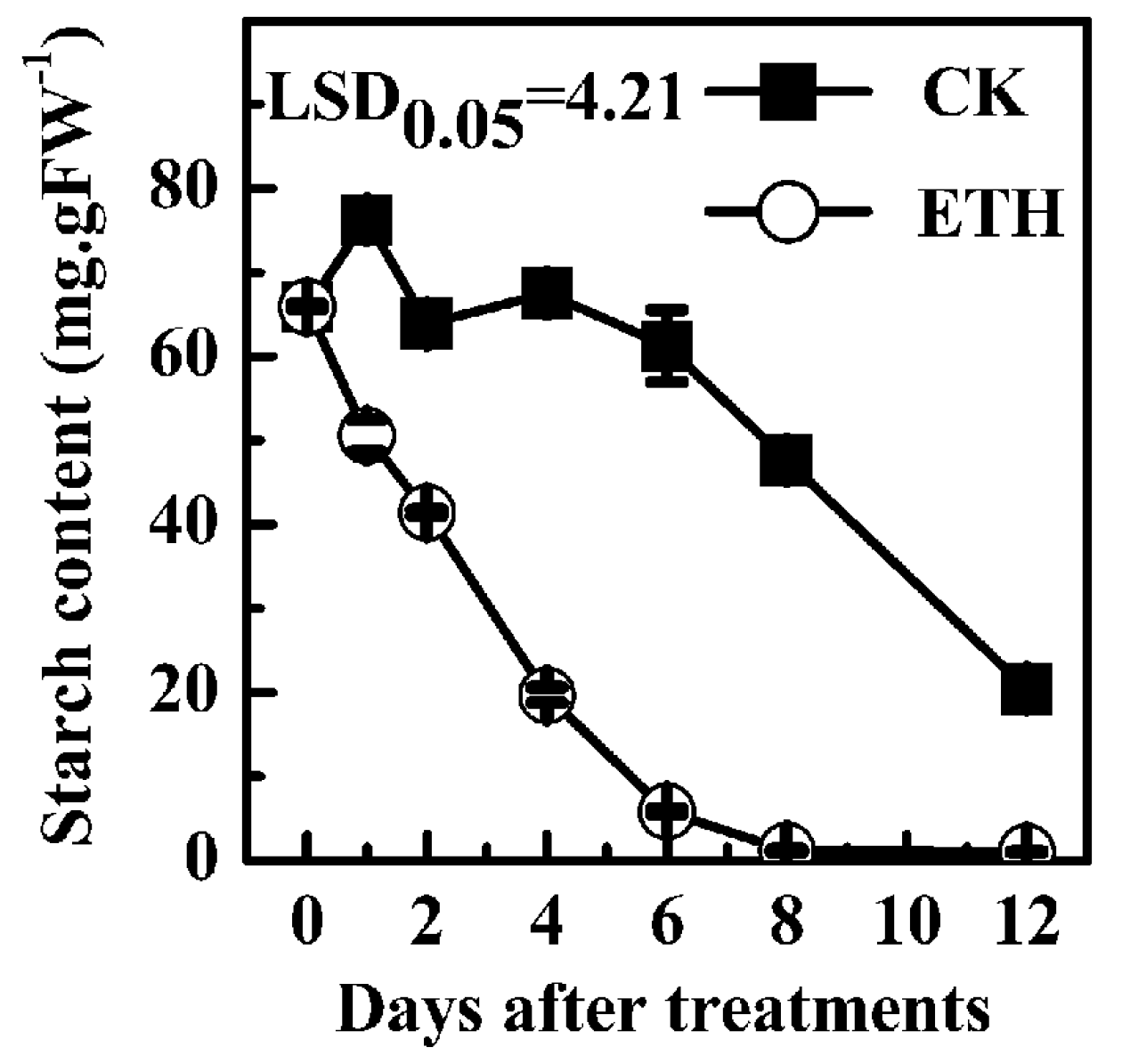
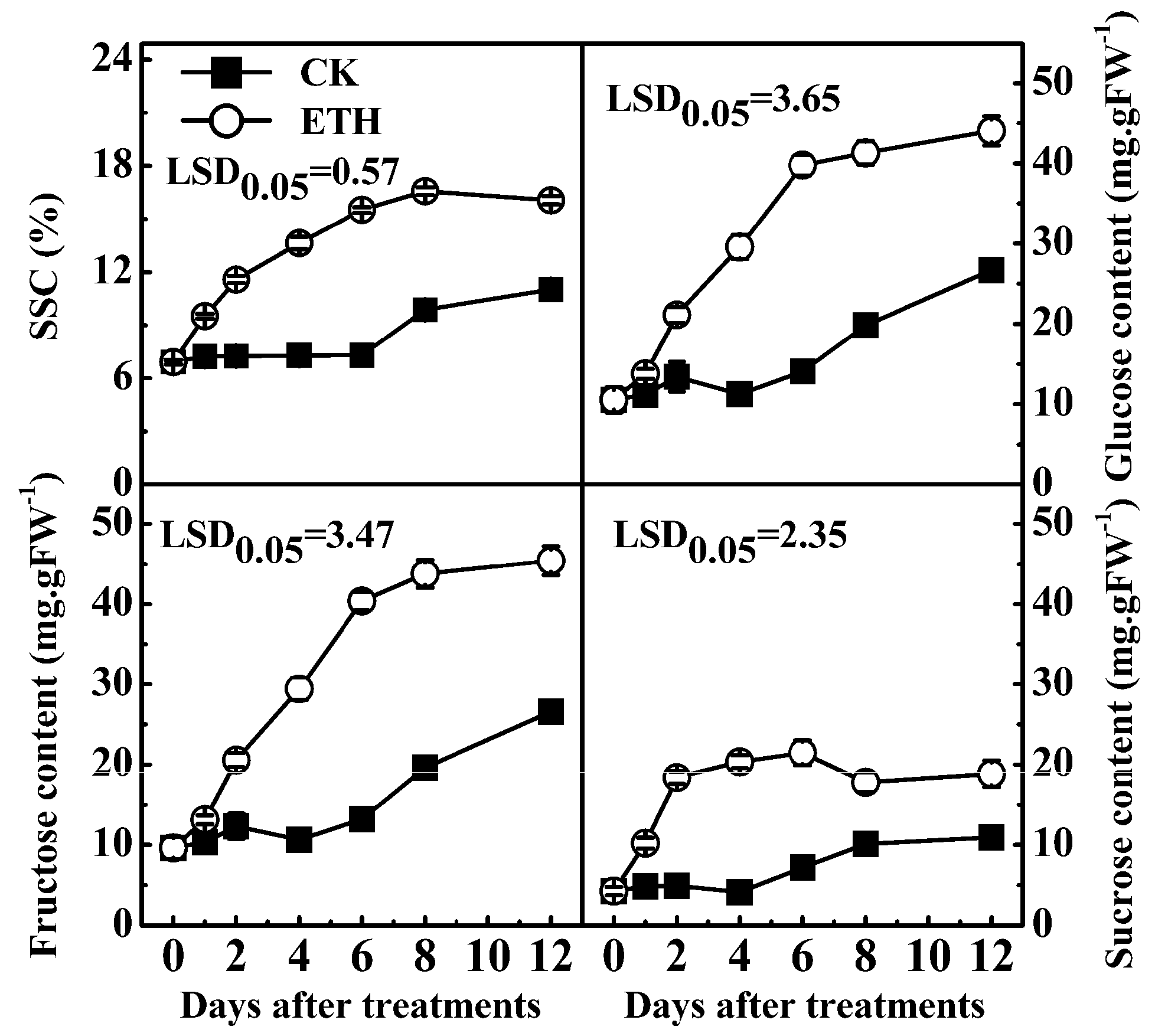
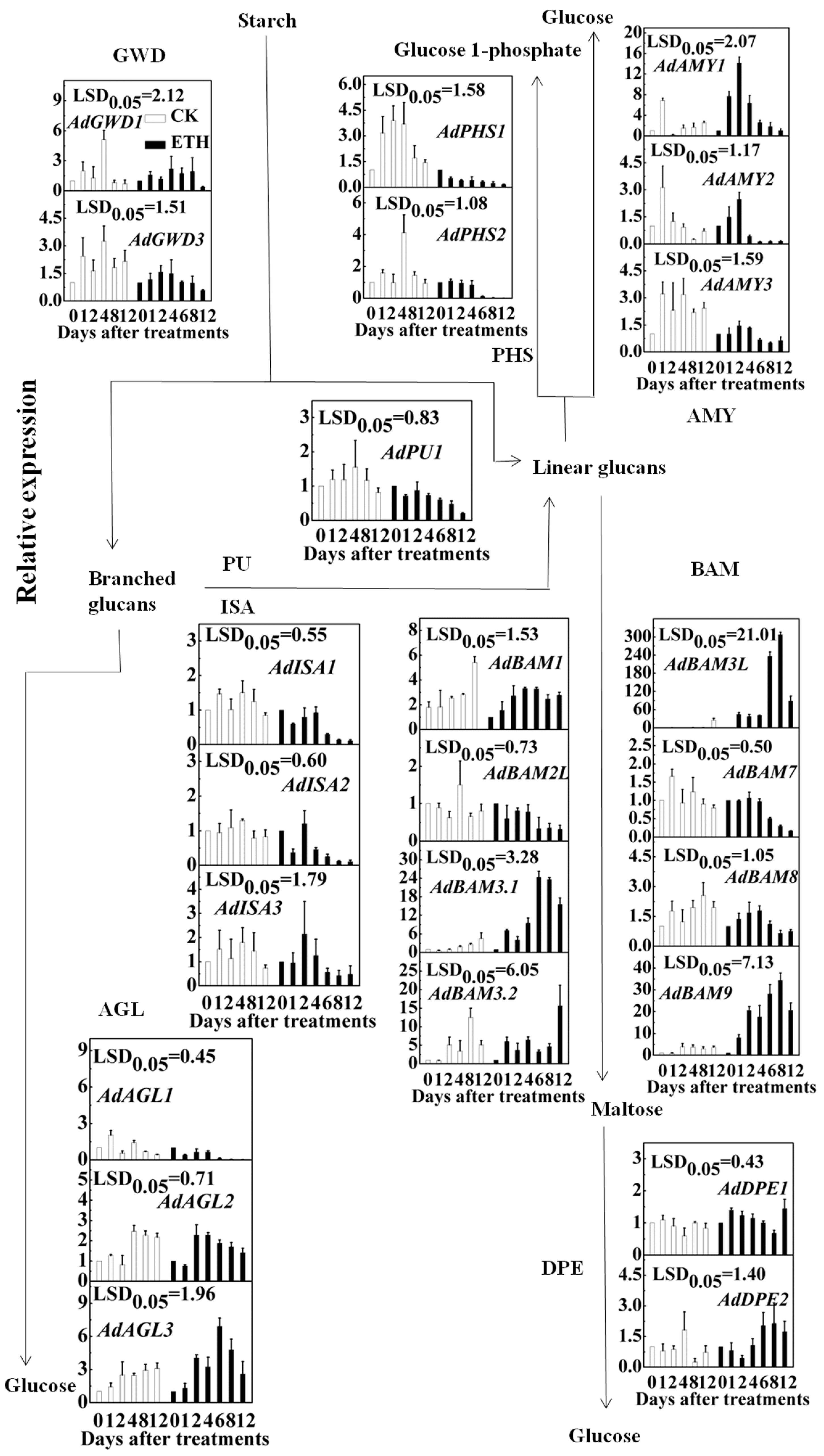
2.3. Expression of Kiwifruit Starch Related Genes in Response to Ethylene Treatment
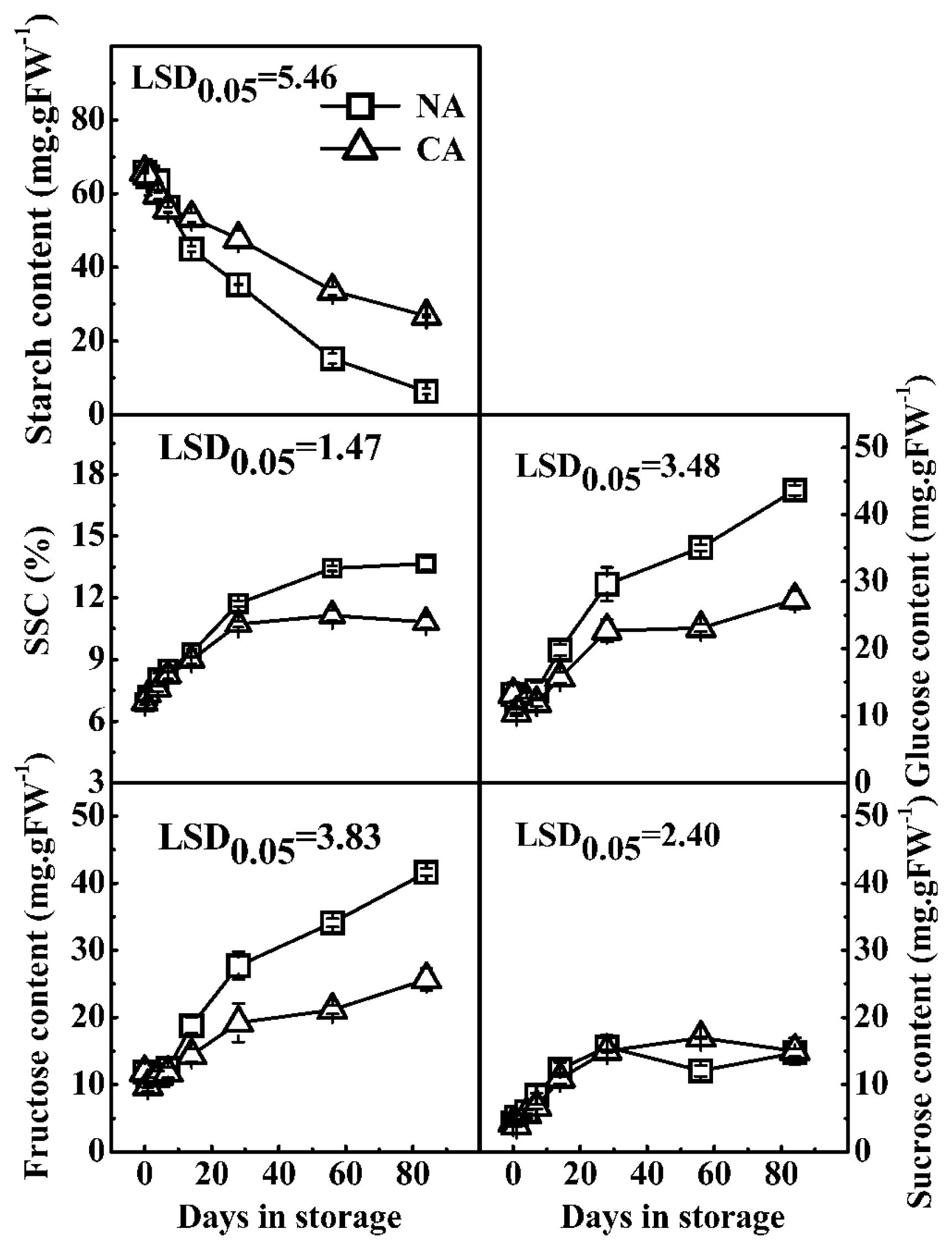
2.4. Effect of CA Storage on Starch Degradation and Related Genes in Kiwifruit at 0 °C
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Soluble Solids Content (SSC), Starch and Soluble Sugar Measurements
4.3. RNA Extraction and cDNA Synthesis
4.4. Gene Isolation and Analysis
4.5. Real-Time PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AMY | α-Amylase |
| BAM | β-Amylase |
| CA | Controlled atmosphere |
| ETH | Ethylene |
| ISA | Isoamlase |
| GWD | Glucan water dikinase |
| PHS | Glucan phosphorylase |
| DPE | 4-α-Glucanotransferase |
| PU | Pullulanase |
| AGL | α-Glucosidase |
| SSC | Soluble solids content |
| GC | Gas chromatography |
References
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant. Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Zeeman, S.C. Starch turnover: Pathways, regulation and role in growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Zeeman, S.C.; Smith, S.M. Starch degradation. Annu. Rev. Plant Biol. 2005, 56, 73–98. [Google Scholar] [CrossRef] [PubMed]
- Caspar, T.; Lin, T.P.; Kakefuda, G.; Benbow, L.; Preiss, J.; Somerville, C. Mutants of Arabidopsis with altered regulation of starch degradation. Plant Physiol. 1991, 95, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Schulze, W.; Stitt, M.; Schulze, E.D.; Neuhaus, H.E.; Fichtner, K. A quantification of the significance of assimilatory starch for growth of Arabidopsis thaliana L. Heynh. Plant Physiol. 1991, 95, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Bertin, N.; Causse, M.; Brunel, B.; Tricon, D.; Génard, M. Identification of growth processes involved in QTLs for tomato fruit size and composition. J. Exp. Bot. 2009, 60, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.G.; Domoto, P.A.; Jane, J.L. Structures and functional properties of apple (Malus. domestica Borkh) fruit starch. Carbohydr. Polym. 2006, 63, 432–441. [Google Scholar] [CrossRef]
- MacRae, E.; Quick, W.P.; Benker, C.; Stitt, M. Carbohydrate metabolism during postharvest ripening in kiwifruit. Planta 1992, 188, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Peroni, F.H.G.; Koike, C.; Louro, R.P.; Purgatto, E.; Nascimento, J.R.O.; Lajolo, F.M.; Cordenunsi, B.R. Mango starch degradation II—The binding of α-Amylase and β-Amylase to the starch granule. J. Agric. Food Chem. 2008, 56, 7416–7421. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, J.R.O.; Júnior, A.V.; Bassinello, P.Z.; Cordenunsi, B.R.; Mainardi, J.A.; Purgatto, E.; Lajolo, F.M. β-amylase expression and starch degradation during banana ripening. Postharvest Biol. Technol. 2006, 40, 41–47. [Google Scholar] [CrossRef]
- Stevenson, D.G.; Johnson, S.R.; Jane, J.L.; Ingletta, G.E. Chemical and physical properties of kiwifruit (Actinidia. deliciosa) starch. Starch 2006, 58, 323–329. [Google Scholar] [CrossRef]
- Atkinson, R.G.; Gunaseelan, K.; Wang, M.Y.; Luo, L.; Wang, T.C.; Norling, C.L.; Johnston, S.L.; Maddumage, R.; Schröder, R.; Schaffer, R.J. Dissecting the role of climacteric ethylene in kiwifruit (Actinidia. chinensis) ripening using a 1-aminocyclopropane-1-carboxylic acid oxidase knockdown line. J. Exp. Bot. 2011, 62, 3821–3835. [Google Scholar] [CrossRef] [PubMed]
- Lorberth, R.; Ritte, G.; Willmitzer, L.; Kossmann, J. Inhibition of a starch-granule-bound protein leads to modified starch and repression of cold-induced sweetening. Nat. Biotechnol. 1998, 16, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Ritte, G.; Lloyd, J.R.; Eckermann, N.; Rottmann, A.; Kossmann, J.; Steup, M. The starch-related R1 protein is an α-glucan, water dikinase. Proc. Natl. Acad. Sci. USA 2002, 99, 7166–7171. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.H.; Deiting, U.; Stitt, M. A β-Amylase in potato tubers is induced by storage at low temperature. Plant Physiol. 1997, 113, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Chia, T.; Thornetycroft, D.; Chapple, A.; Messerli, G.; Chen, J.; Zeeman, S.C.; Smith, S.M.; Smith, A.M. A cytosolic glucosyltransferase is required for conversion of starch to sucrose in Arabidopsis leaves at night. Plant J. 2004, 37, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Critchley, J.H.; Zeeman, S.C.; Takaha, T.; Smith, A.M.; Smith, S.M. A critical role for disproportionating enzyme in starch breakdown is revealed by a knock-out mutation in Arabidopsis. Plant J. 2001, 26, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.S.; Kofler, H.; Hausler, R.E.; Hille, D.; Flügge, U.I.; Zeeman, S.C.; Smith, A.M.; Kossmann, J.; Lloyd, J.; Ritte, G.; et al. The Arabidopsis sex1 mutant is defective in the R1 protein, a general regulator of starch degradation in plants, and not in the chloroplast hexose transporter. Plant Cell 2001, 13, 1907–1918. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.S.; Zeeman, S.C.; Thorneycroft, D.; Fulton, D.C.; Dunstan, H.; Lue, W.L.; Hegemann, B.; Tung, S.Y.; Umemoto, T.; Chapple, A.; et al. α-Amylase is not required for breakdown of transitory starch in Arabidopsis leaves. J. Biol. Chem. 2005, 280, 9773–9779. [Google Scholar] [CrossRef] [PubMed]
- Zeeman, S.C.; Thorneycroft, D.; Schupp, N.; Chapple, A.; Weck, M.; Dunstan, H.; Haldimann, P.; Bechtold, N.; Smith, A.M.; Smith, S.M. Plastidial α-glucan phosphorylase is not required for starch degradation in Arabidopsis leaves but has a role in the tolerance of abiotic stress. Plant Physiol. 2004, 135, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Skeffington, A.W.; Graf, A.; Duxbury, Z.; Gruissem, W.; Smith, A.M. Glucan, water dikinase exerts little control over starch degradation in Arabidopsis leaves at night. Plant Physiol. 2014, 165, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Mahlow, S.; Hejazi, M.; Kuhnert, F.; Garz, A.; Brust, H.; Baumann, O.; Fettke, J. Phosphorylation of transitory starch by α-glucan, water dikinase during starch turnover affects the surface properties and morphology of starch granules. New Phytol. 2014, 203, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Bassinello, P.Z.; Cordenuns, B.R.; Lajolo, F.M. Amylolytic activity in fruits: Comparison of different substrates and methods using banana as model. J. Agric. Food Chem. 2002, 50, 5781–5786. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.; Lajolo, F.M. Starch transformation during banana ripening: the amylase and glucosidase behavior. J. Food Sci. 1988, 53, 1181–1186. [Google Scholar] [CrossRef]
- Bierhals, J.D.; Lajolo, F.M.; Cordenunsi, B.R.; Nascimento, J.R.O. Activity, cloning, and expression of an isoamylase-type starch-debranching enzyme from banana fruit. J. Agric. Food Chem. 2004, 52, 7412–7418. [Google Scholar] [CrossRef] [PubMed]
- Da Mota, R.V.; Cordenunsi, B.R.; do Nascimento, J.R.O.; Purgatto, E.; Rosseto, M.R.M.; Lajolo, F.M. Activity and expression of banana starch phosphorylases during fruit development and ripening. Planta 2002, 216, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Bonghi, C.; Pagni, S.; Vidrih, R.; Ramina, A.; Tonutti, P. Cell wall hydrolases and amylase in kiwifruit softening. Postharvest Biol. Technol. 1996, 9, 19–29. [Google Scholar] [CrossRef]
- Wegrzyn, T.; MacRae, E. α-Amylase and starch degradation in kiwifruit. J. Plant Physiol. 1995, 147, 19–28. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Osorio, S.; Höhne, M.; Wohlers, M.; Gleave, A.P.; MacRae, E.A.; Richardson, A.C.; Atkinson, R.G.; Sulpice, R.; et al. Metabolic analysis of kiwifruit (Actinidia. deliciosa) berries from extreme genotypes reveals hallmarks for fruit starch metabolism. J. Exp. Bot. 2013, 64, 5049–5063. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Obara, T.; Luh, B.S. Changes in chemical constituents of kiwifruit during post-harvest ripening. J. Food Sci. 1983, 48, 607–611. [Google Scholar] [CrossRef]
- Zhang, A.D.; Hu, X.; Kuang, S.; Ge, H.; Yin, X.R.; Chen, K.S. Isolation, classification and transcription profiles of the Ethylene Response Factors (ERFs) in ripening kiwifruit. Sci. Hortic. 2016, 199, 209–215. [Google Scholar] [CrossRef]
- Li, X.; Xu, C.J.; Korban, S.S.; Chen, K.S. Regulatory mechanism of textural changes in ripening fruits. Crit. Rev. Plant Sci. 2010, 29, 222–243. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Prot. 2006, 1, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.R.; Zhang, Y.; Zhang, B.; Yang, S.L.; Shi, Y.N.; Ferguson, I.B.; Chen, K.S. Effects of acetylsalicylic acid on kiwifruit ethylene biosynthesis and signaling components. Postharvest Biol. Technol. 2013, 83, 27–33. [Google Scholar] [CrossRef]
- Huang, S.X.; Ding, J.; Deng, D.J.; Tang, W.; Sun, H.H.; Liu, D.Y.; Zhang, L.; Niu, X.L.; Zhang, X.; Meng, M.; et al. Draft genome of the kiwifruit Actinida chinensis. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.R.; Chen, K.S.; Allan, A.C.; Wu, R.M.; Zhang, B.; Lallu, N.; Ferguson, I.B. Ethylene-induced modulation of genes associated with the ethylene signalling pathway in ripening kiwifruit. J. Exp. Bot. 2008, 59, 2097–2108. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Kuang, S.; Zhang, A.-D.; Zhang, W.-S.; Chen, M.-J.; Yin, X.-R.; Chen, K.-S. Characterization of Starch Degradation Related Genes in Postharvest Kiwifruit. Int. J. Mol. Sci. 2016, 17, 2112. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17122112
Hu X, Kuang S, Zhang A-D, Zhang W-S, Chen M-J, Yin X-R, Chen K-S. Characterization of Starch Degradation Related Genes in Postharvest Kiwifruit. International Journal of Molecular Sciences. 2016; 17(12):2112. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17122112
Chicago/Turabian StyleHu, Xiong, Sheng Kuang, Ai-Di Zhang, Wang-Shu Zhang, Miao-Jin Chen, Xue-Ren Yin, and Kun-Song Chen. 2016. "Characterization of Starch Degradation Related Genes in Postharvest Kiwifruit" International Journal of Molecular Sciences 17, no. 12: 2112. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17122112