
Benzyl Isothiocyanate Inhibits Prostate Cancer Development in the Transgenic Adenocarcinoma Mouse Prostate (TRAMP) Model, Which Is Associated with the Induction of Cell Cycle G1 Arrest

 ,
,
Abstract
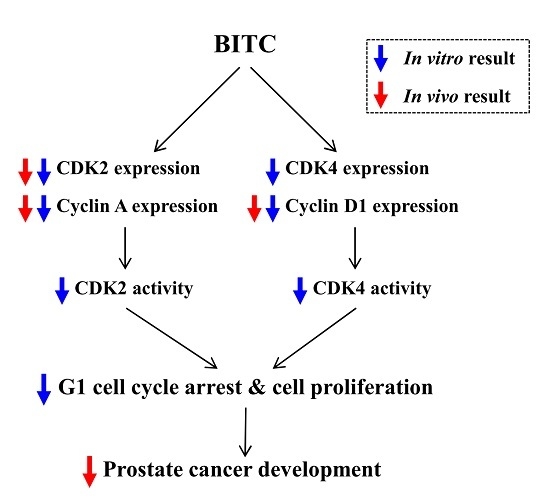
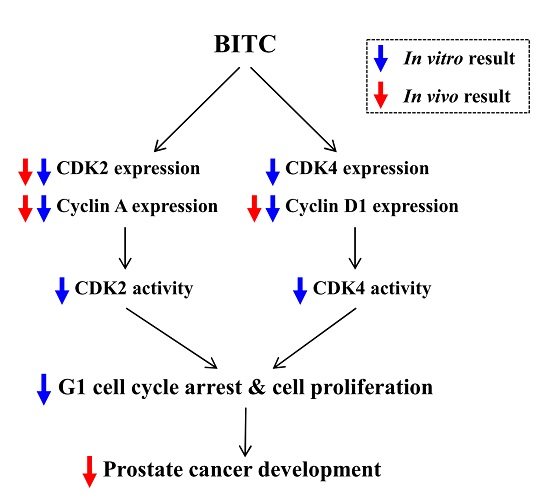
:
1. Introduction
2. Results
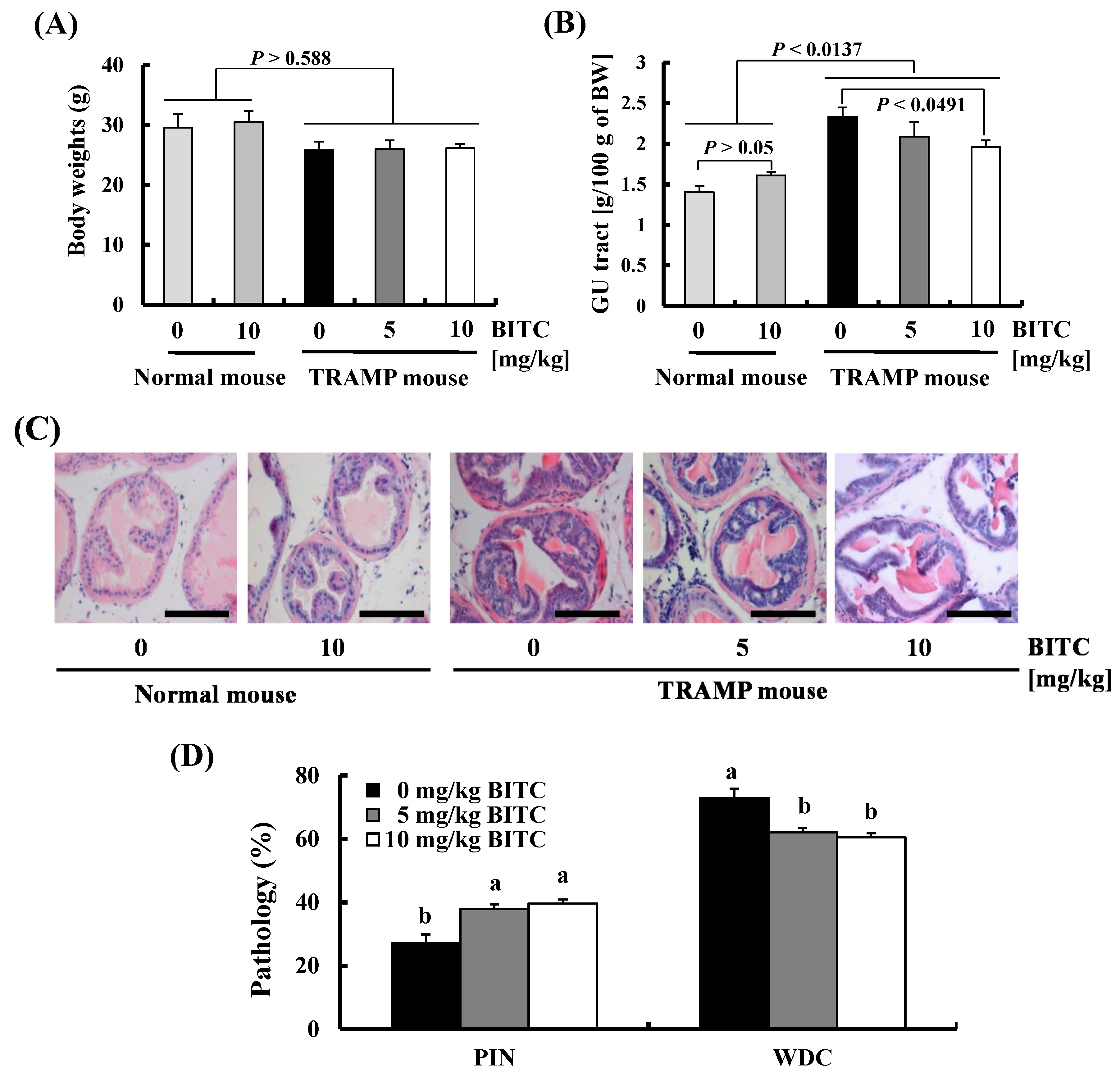
2.1. BITC Inhibits Prostate Cancer Development in TRAMP Mice
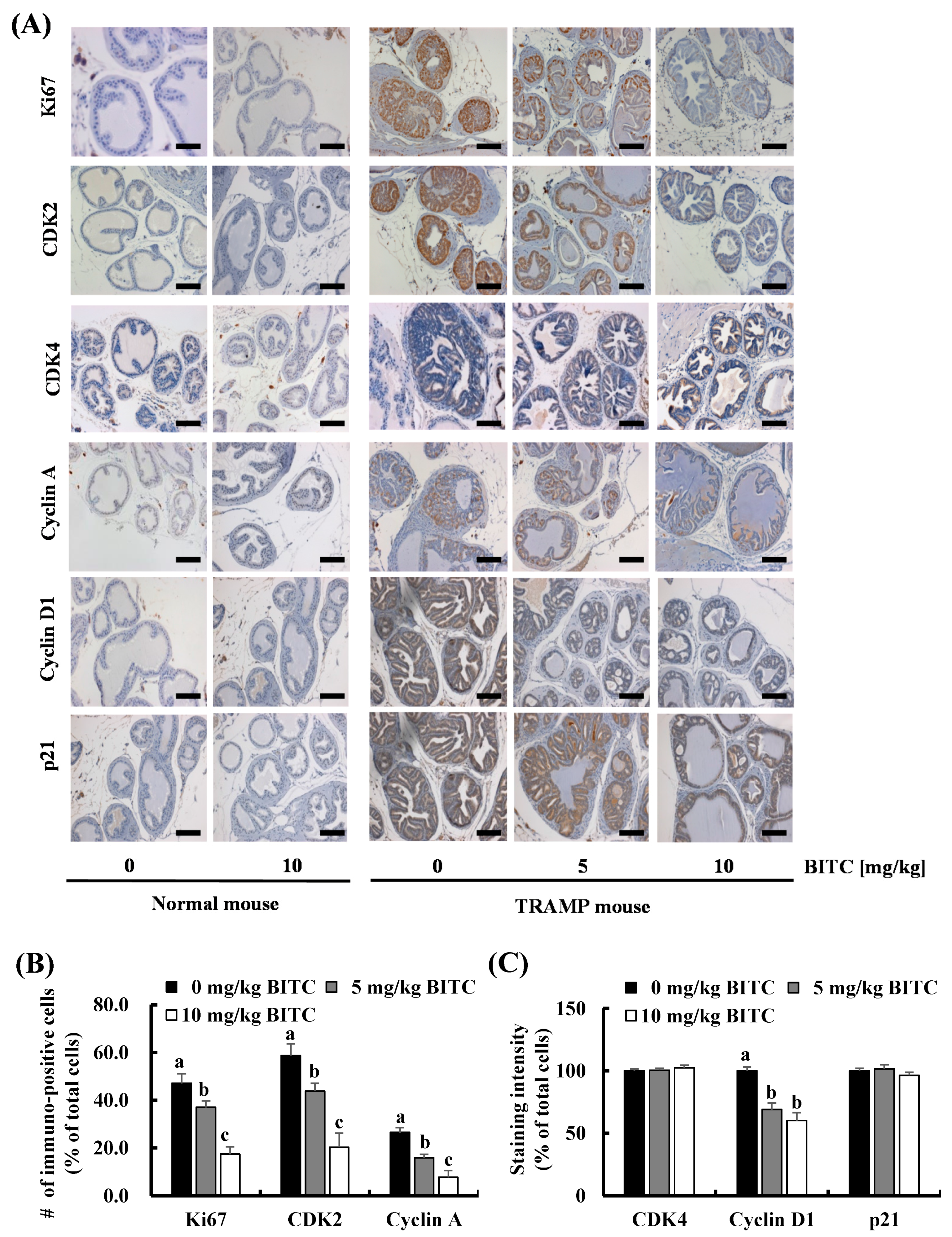
2.2. BITC Inhibits Cell Cycle Progression in the DP in TRAMP Mice
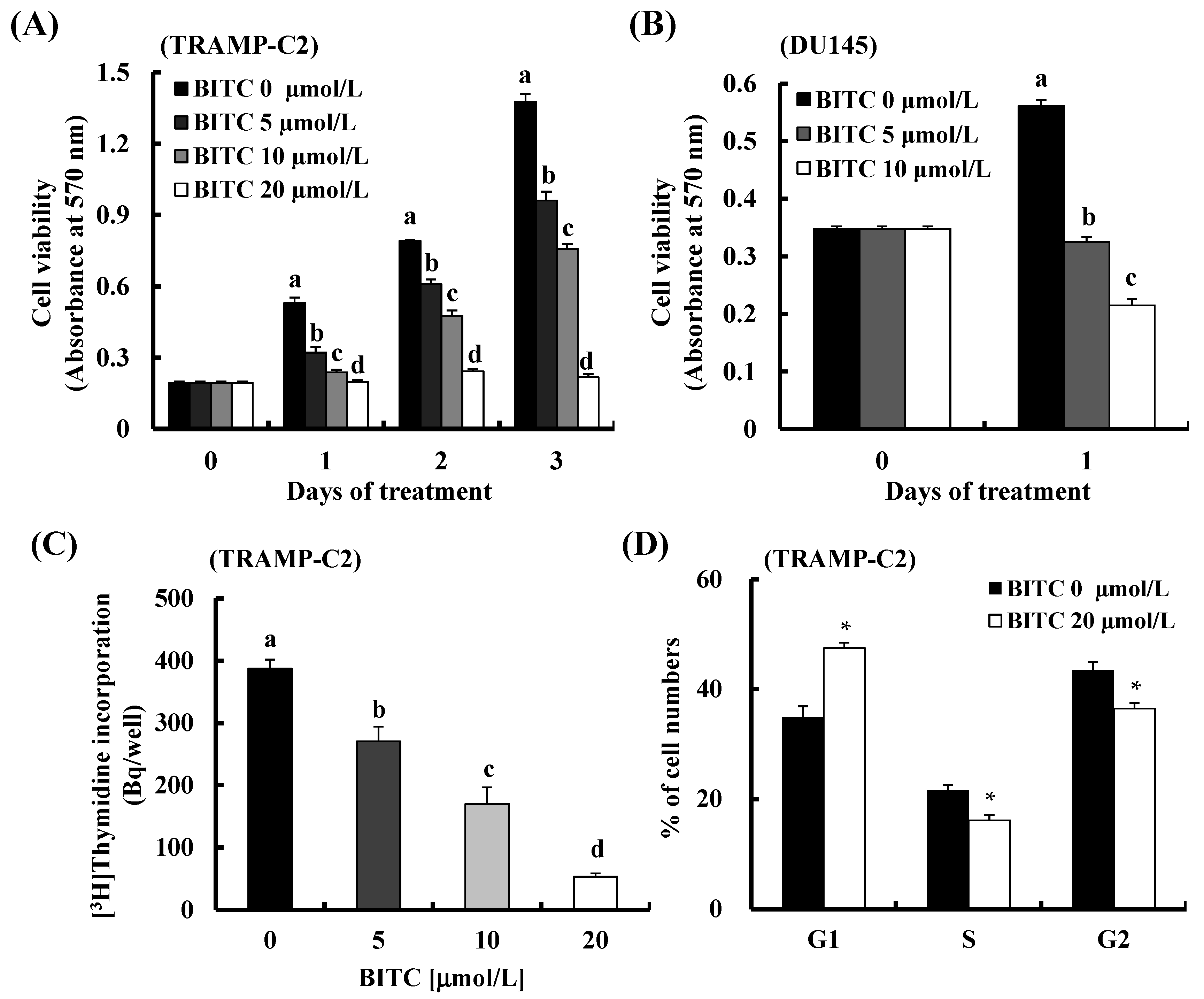
2.3. BITC Induces G1 Cell Cycle Arrest in TRAMP-C2 Cells
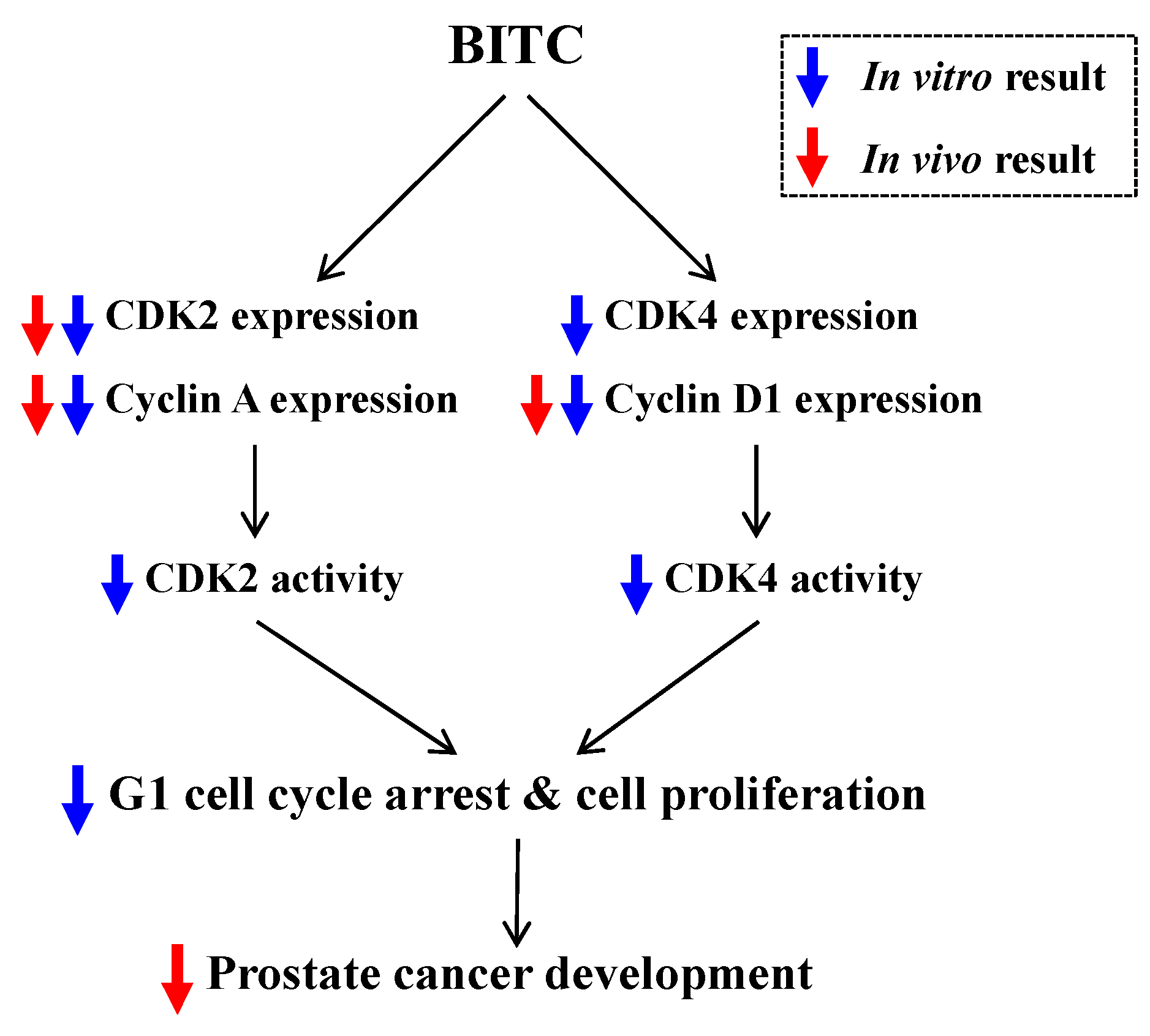
2.4. BITC Inhibits the Expression of Cyclins and the Activity of CDKs in TRAMP-C2 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Treatments
4.3. Immunohistochemical Analysis
4.4. Cell Culture
4.5. (3H)Thymidine Incorporation
4.6. Flow Cytometry Analysis of Cell Cycle Distribution
4.7. Western Blot Analyses and Determination of CDK Activity
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.K.; Sporn, M.B. Recent advances in chemoprevention of cancer. Science 1997, 278, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Kristal, A.R.; Lampe, J.W. Brassica vegetables and prostate cancer risk: A review of the epidemiological evidence. Nutr. Cancer 2002, 42, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.V.; Singh, K. Cancer chemoprevention with dietary isothiocyanates mature for clinical translational research. Carcinogenesis 2012, 33, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Wicker, C.A.; Sahu, R.P.; Kulkarni-Datar, K.; Srivastava, S.K.; Brown, T.L. BITC sensitizes pancreatic adenocarcinomas to TRAIL-induced apoptosis. Cancer Growth Metastasis 2010, 2009, 45–55. [Google Scholar] [PubMed]
- Kim, S.H.; Singh, S.V. p53-Independent apoptosis by benzyl isothiocyanate in human breast cancer cells is mediated by suppression of XIAP expression. Cancer Prev. Res. 2010, 3, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Lai, K.C.; Hsu, S.C.; Kuo, C.L.; Ma, C.Y.; Lin, M.L.; Yang, J.S.; Chung, J.G. Benzyl isothiocyanate (BITC) inhibits migration and invasion of human gastric cancer AGS cells via suppressing ERK signal pathways. Hum. Exp. Toxicol. 2011, 30, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.C.; Huang, A.C.; Hsu, S.C.; Kuo, C.L.; Yang, J.S.; Wu, S.H.; Chung, J.G. Benzyl isothiocyanate (BITC) inhibits migration and invasion of human colon cancer HT29 cells by inhibiting matrix metalloproteinase-2/-9 and urokinase plasminogen (uPA) through PKC and MAPK signaling pathway. J. Agric. Food Chem. 2010, 58, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Haldar, S. Dietary isothiocyanate mediated apoptosis of human cancer cells is associated with Bcl-xL phosphorylation. Int. J. Oncol. 2008, 33, 657–663. [Google Scholar] [PubMed]
- Lin, J.F.; Tsai, T.F.; Liao, P.C.; Lin, Y.H.; Lin, Y.C.; Chen, H.E.; Chou, K.Y.; Hwang, T.I. Benzyl isothiocyanate induces protective autophagy in human prostate cancer cells via inhibition of mTOR signaling. Carcinogenesis 2013, 34, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Cho, H.J.; Park, S.Y.; Kim, E.J.; Kim, J.K.; Park, J.H. 3,3′-Diindolylmethane inhibits prostate cancer development in the transgenic adenocarcinoma mouse prostate model. Mol. Carcinog. 2011, 50, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Y.; Saw, C.L.; Khor, T.O.; Pung, D.; Boyanapalli, S.S.; Kong, A.N. In vivo pharmacodynamics of indole-3-carbinol in the inhibition of prostate cancer in transgenic adenocarcinoma of mouse prostate (TRAMP) mice: Involvement of Nrf2 and cell cycle/apoptosis signaling pathways. Mol. Carcinog. 2012, 51, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Barve, A.; Khor, T.O.; Hao, X.; Keum, Y.S.; Yang, C.S.; Reddy, B.; Kong, A.N. Murine prostate cancer inhibition by dietary phytochemicals—Curcumin and phenyethylisothiocyanate. Pharm. Res. 2008, 25, 2181–2189. [Google Scholar] [CrossRef] [PubMed]
- Nurse, P. A long twentieth century of the cell cycle and beyond. Cell 2000, 100, 71–78. [Google Scholar] [CrossRef]
- Malumbres, M. Cyclin-dependent kinases. Genome Biol. 2014, 15. [Google Scholar] [CrossRef]
- Vermeulen, K.; van Bockstaele, D.R.; Berneman, Z.N. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003, 36, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Bhaskaran, N.; Babcook, M.A.; Fu, P.; Maclennan, G.T.; Gupta, S. Apigenin inhibits prostate cancer progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway. Carcinogenesis 2014, 35, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, N.M.; DeMayo, F.; Finegold, M.J.; Medina, D.; Tilley, W.D.; Aspinall, J.O.; Cunha, G.R.; Donjacour, A.A.; Matusik, R.J.; Rosen, J.M. Prostate cancer in a transgenic mouse. Proc. Natl. Acad. Sci. USA 1995, 92, 3439–3443. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Lefko, P.J.; Chen, T.M.; Ittmann, M.M.; Barrios, R.J.; Ayala, G.E.; Huss, W.J.; Maddison, L.A.; Foster, B.A.; Greenberg, N.M. Pathobiology of autochthonous prostate cancer in a pre-clinical transgenic mouse model. Prostate 2003, 55, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Vogel, V.; Singh, S.V. Benzyl isothiocyanate-induced apoptosis in human breast cancer cells is initiated by reactive oxygen species and regulated by Bax and Bak. Mol. Cancer Ther. 2006, 5, 2931–2945. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.N.; Yan, H.Q.; Wu, X.; Pan, Z.H.; Zhu, Y.; Meng, Z.W.; Zhou, Q.H.; Xu, K. Apoptosis induced by benzyl isothiocyanate in gefitinib-resistant lung cancer cells is associated with Akt/MAPK pathways and generation of reactive oxygen species. Cell Biochem. Biophys. 2013, 66, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Sahu, R.P.; Zhang, R.; Batra, S.; Shi, Y.; Srivastava, S.K. Benzyl isothiocyanate-mediated generation of reactive oxygen species causes cell cycle arrest and induces apoptosis via activation of MAPK in human pancreatic cancer cells. Carcinogenesis 2009, 30, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Uchida, K.; Osawa, T.; Nakamura, Y. A link between benzyl isothiocyanate-induced cell cycle arrest and apoptosis: Involvement of mitogen-activated protein kinases in the Bcl-2 phosphorylation. Cancer Res. 2004, 64, 2134–2142. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Li, G.; Cao, B.; Liu, L.; Cheng, Q.; Kong, H.; Shan, C.; Huang, X.; Chen, J.; Gao, N. Downregulation of Mcl-1 through inhibition of translation contributes to benzyl isothiocyanate-induced cell cycle arrest and apoptosis in human leukemia cells. Cell Death Dis. 2013, 4, e515. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.C.; Huang, Y.T.; Wu, P.P.; Ji, B.C.; Yang, J.S.; Yang, J.L.; Chiu, T.H.; Chueh, F.S.; Chung, J.G. The roles of AIF and Endo G in the apoptotic effects of benzyl isothiocyanate on DU 145 human prostate cancer cells via the mitochondrial signaling pathway. Int. J. Oncol. 2011, 38, 787–796. [Google Scholar] [PubMed]
- Foster, B.A.; Gingrich, J.R.; Kwon, E.D.; Madias, C.; Greenberg, N.M. Characterization of prostatic epithelial cell lines derived from transgenic adenocarcinoma of the mouse prostate (TRAMP) model. Cancer Res. 1997, 57, 3325–3330. [Google Scholar] [PubMed]
- Higdon, J.V.; Delage, B.; Williams, D.E.; Dashwood, R.H. Cruciferous vegetables and human cancer risk: Epidemiologic evidence and mechanistic basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Powolny, A.A.; Bommareddy, A.; Hahm, E.R.; Normolle, D.P.; Beumer, J.H.; Nelson, J.B.; Singh, S.V. Chemopreventative potential of the cruciferous vegetable constituent phenethyl isothiocyanate in a mouse model of prostate cancer. J. Natl. Cancer Inst. 2011, 103, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.V.; Warin, R.; Xiao, D.; Powolny, A.A.; Stan, S.D.; Arlotti, J.A.; Zeng, Y.; Hahm, E.R.; Marynowski, S.W.; Bommareddy, A.; et al. Sulforaphane inhibits prostate carcinogenesis and pulmonary metastasis in TRAMP mice in association with increased cytotoxicity of natural killer cells. Cancer Res. 2009, 69, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Warin, R.; Xiao, D.; Arlotti, J.A.; Bommareddy, A.; Singh, S.V. Inhibition of human breast cancer xenograft growth by cruciferous vegetable constituent benzyl isothiocyanate. Mol. Carcinog. 2010, 49, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.Y.; Hsiao, Y.P.; Hsu, S.C.; Hsueh, S.C.; Chang, C.H.; Ji, B.C.; Yang, J.S.; Lu, H.F.; Chung, J.G. Oral administration of benzyl-isothiocyanate inhibits in vivo growth of subcutaneous xenograft tumors of human malignant melanoma A375.S2 cells. Vivo 2013, 27, 623–626. [Google Scholar]
- Kim, M.; Cho, H.J.; Kwon, G.T.; Kang, Y.H.; Kwon, S.H.; Her, S.; Park, T.; Kim, Y.; Kee, Y.; Park, J.H. Benzyl isothiocyanate suppresses high-fat diet-stimulated mammary tumor progression via the alteration of tumor microenvironments in obesity-resistant BALB/c mice. Mol. Carcinog. 2015, 54, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Moussavi, M.; Tearle, H.; Fazli, L.; Bell, J.C.; Jia, W.; Rennie, P.S. Targeting and killing of metastatic cells in the transgenic adenocarcinoma of mouse prostate model with vesicular stomatitis virus. Mol. Ther. 2013, 21, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Warin, R.; Chambers, W.H.; Potter, D.M.; Singh, S.V. Prevention of mammary carcinogenesis in MMTV-neu mice by cruciferous vegetable constituent benzyl isothiocyanate. Cancer Res. 2009, 69, 9473–9480. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Adkins, C.; Lockman, P.; Srivastava, S.K. Metastasis of breast tumor cells to brain is suppressed by phenethyl isothiocyanate in a novel metastasis model. PLoS ONE 2013, 8, e67278. [Google Scholar] [CrossRef] [PubMed]
- Olvera-Caltzontzin, P.; Delgado, G.; Aceves, C.; Anguiano, B. Iodine uptake and prostate cancer in the TRAMP mouse model. Mol. Med. 2013, 19, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Rizzo, P.; Grimley, P.M.; Procopio, A.; Mew, D.J.; Shridhar, V.; de Bartolomeis, A.; Esposito, V.; Giuliano, M.T.; Steinberg, S.M.; et al. Simian virus-40 large-T antigen binds p53 in human mesotheliomas. Nat. Med. 1997, 3, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Zilfou, J.T.; Lowe, S.W. Tumor suppressive functions of p53. Cold Spring Harb. Perspect. Biol. 2009, 1, a001883. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, P.G.; Woodhead, A.J.; Berdini, V.; Boulstridge, J.A.; Carr, M.G.; Cross, D.M.; Davis, D.J.; Devine, L.A.; Early, T.R.; Feltell, R.E.; et al. Identification of N-(4-piperidinyl)-4-(2,6-dichlorobenzoylamino)-1H-pyrazole-3-carboxamide (AT7519), a novel cyclin dependent kinase inhibitor using fragment-based X-ray crystallography and structure based drug design. J. Med. Chem. 2008, 51, 4986–4999. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, N.M.; DeMayo, F.J.; Sheppard, P.C.; Barrios, R.; Lebovitz, R.; Finegold, M.; Angelopoulou, R.; Dodd, J.G.; Duckworth, M.L.; Rosen, J.M.; et al. The rat probasin gene promoter directs hormonally and developmentally regulated expression of a heterologous gene specifically to the prostate in transgenic mice. Mol. Endocrinol. 1994, 8, 230–239. [Google Scholar] [PubMed]
- Gingrich, J.R.; Barrios, R.J.; Foster, B.A.; Greenberg, N.M. Pathologic progression of autochthonous prostate cancer in the TRAMP model. Prostate Cancer Prostatic Dis. 1999, 2, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.Y.; Jeong, Y.; Tyner, A.L.; Park, J.H. Induction of cell cycle arrest and apoptosis in HT-29 human colon cancer cells by the dietary compound luteolin. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G66–G75. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Kim, W.K.; Kim, E.J.; Jung, K.C.; Park, S.; Lee, H.S.; Tyner, A.L.; Park, J.H. Conjugated linoleic acid inhibits cell proliferation and ErbB3 signaling in HT-29 human colon cell line. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G996–G1005. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BITC (mg/kg) | Normal Mouse | TRAMP Mouse | |||
|---|---|---|---|---|---|
| 0 | 10 | 0 | 5 | 10 | |
| Liver weight | 4.06 ± 0.05 | 4.1 ± 0.10 | 4.39 ± 0.17 | 3.99 ± 0.21 | 3.96 ± 0.22 |
| Lung weight | 0.58 ± 0.06 | 0.54 ± 0.02 | 0.59 ± 0.02 | 0.61 ± 0.02 | 0.61 ± 0.04 |
| Spleen weight | 0.27 ± 0.04 | 0.21 ± 0.02 | 0.27 ± 0.03 | 0.31 ± 0.03 | 0.29 ± 0.03 |
| BITC (mg/kg) | Normal Mouse | TRAMP Mouse | |||
|---|---|---|---|---|---|
| 0 | 10 | 0 | 5 | 10 | |
| Creatinine (nmol/L) | 0.20 ± 0.02 | 0.23 ± 0.05 | 0.25 ± 0.03 | 0.20 ± 0.01 | 0.18 ± 0.01 |
| AST (U/L) | 81.1 ± 6.47 | 59.9 ± 9.73 | 59.5 ± 3.69 | 52.8 ± 8.62 | 58.6 ± 8.89 |
| ALT(U/L) | 30.8 ± 14.0 | 9.50 ± 0.50 | 20.6 ± 5.13 | 21.3 ± 7.06 | 13.2 ± 1.74 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.J.; Lim, D.Y.; Kwon, G.T.; Kim, J.H.; Huang, Z.; Song, H.; Oh, Y.S.; Kang, Y.-H.; Lee, K.W.; Dong, Z.; et al. Benzyl Isothiocyanate Inhibits Prostate Cancer Development in the Transgenic Adenocarcinoma Mouse Prostate (TRAMP) Model, Which Is Associated with the Induction of Cell Cycle G1 Arrest. Int. J. Mol. Sci. 2016, 17, 264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17020264
Cho HJ, Lim DY, Kwon GT, Kim JH, Huang Z, Song H, Oh YS, Kang Y-H, Lee KW, Dong Z, et al. Benzyl Isothiocyanate Inhibits Prostate Cancer Development in the Transgenic Adenocarcinoma Mouse Prostate (TRAMP) Model, Which Is Associated with the Induction of Cell Cycle G1 Arrest. International Journal of Molecular Sciences. 2016; 17(2):264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17020264
Chicago/Turabian StyleCho, Han Jin, Do Young Lim, Gyoo Taik Kwon, Ji Hee Kim, Zunnan Huang, Hyerim Song, Yoon Sin Oh, Young-Hee Kang, Ki Won Lee, Zigang Dong, and et al. 2016. "Benzyl Isothiocyanate Inhibits Prostate Cancer Development in the Transgenic Adenocarcinoma Mouse Prostate (TRAMP) Model, Which Is Associated with the Induction of Cell Cycle G1 Arrest" International Journal of Molecular Sciences 17, no. 2: 264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17020264