
The Role of Sialyl-Tn in Cancer

Institute of Genetic Medicine, Newcastle University, Newcastle-upon-Tyne NE1 3BZ, UK
Int. J. Mol. Sci. 2016, 17(3), 275; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030275
Submission received: 5 February 2016
/
Revised: 17 February 2016
/
Accepted: 18 February 2016
/
Published: 24 February 2016
(This article belongs to the Special Issue Glycan–Receptor Interaction)
{kind=link}
Abstract
:Activation of an aberrant glycosylation pathway in cancer cells can lead to expression of the onco-foetal sialyl-Tn (sTn) antigen. STn is a truncated O-glycan containing a sialic acid α-2,6 linked to GalNAc α-O-Ser/Thr and is associated with an adverse outcome and poor prognosis in cancer patients. The biosynthesis of the sTn antigen has been linked to the expression of the sialytransferase ST6GalNAc1, and also to mutations in and loss of heterozygosity of the COSMC gene. sTn neo- or over-expression occurs in many types of epithelial cancer including gastric, colon, breast, lung, oesophageal, prostate and endometrial cancer. sTn is believed to be carried by a variety of glycoproteins and may influence protein function and be involved in tumour development. This review discusses how the role of sTn in cancer development and tumour cell invasiveness might be organ specific and occur through different mechanisms depending on each cancer type or subtype. As the sTn-antigen is expressed early in carcinogenesis targeting sTn in cancer may enable the targeting of tumours from the earliest stage.
1. Introduction
Aberrant Glycosylation can play an important role in key fundamental processes occurring in cancer and changes in O-glycosylation are frequently observed [1]. O-glycosylation of proteins is a diverse and abundant form of post-translational modification that occurs in the Golgi apparatus and depends on the sequential action of several glycosylation enzymes. The biosynthesis of O-glycans is initiated by a family of up to 20 GalNAc-transferases that transfer N-Acetylgalactosamine (GalNAc) to serine threonine residues on proteins to produce the Tn antigen. The Tn antigen is then further branched and capped in subsequent processing steps by a large number of different glycosyltransferases. In normal cells, O-glycosylation proceeds to mature elongated and branched O-glycans, which are frequently modified with sialic acid. An increase in global sialylation has been closely associated with cancer and can play a fundamental role in cell adhesion, cellular recognition and cell signalling [1,2].
The pattern of glycans expressed in a cell depends on the glycosyltransferases expressed, their substrate specificity, and localisation. In cancer cells the processing of O-glycans to mature branched structures is often disrupted, and they express only early biosynthetic intermediates [3,4]. The sialyl-Tn antigen (Neu5Acα2-6GalNAcα-O-Ser/Thr), known as sTn, is a truncated O-glycan containing a sialic acid α-2,6 linked to GalNAc α-O-Serine/Threonine (Ser/Thr). sTn is expressed by more than 80% of human carcinomas and is linked to poor prognosis in cancer patients. The sTn antigen was first discovered as a cancer marker >30 years ago. Early studies identified the sTn as a marker for diagnosis and prognosis in cancer, and later work focused on using sTn as a potential therapeutic target. sTn neo- or over-expression has been described in many types of epithelial cancer including: gastric [5], colon [6], breast [7,8], lung [9], oesophageal [10], pancreatic [11], prostate [12,13], bladder [14], and endometrial cancer [15]. Studies suggest that sTn expression has primarily been detected at the apical or luminal surface of tissue and tends to occur in early carcinogenesis [16].
2. Synthesis of the Sialyl-Tn (sTn) Antigen
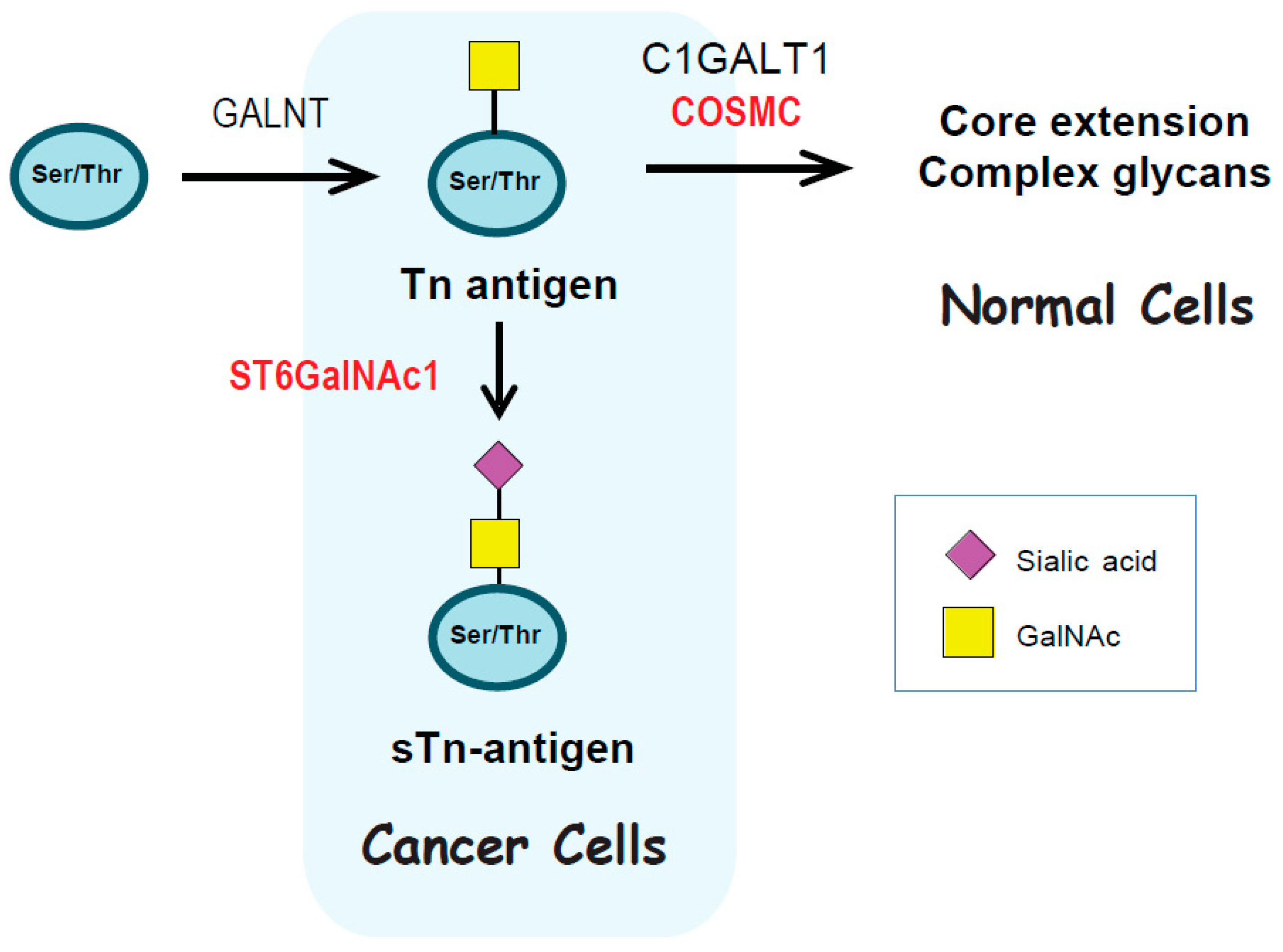
The expression of truncated O-glycans in cancer has been proposed to result from altered expression of glycosyltransferases [17], hypermethylation or mutations in COSMC (core 1 β3-Gal-T-specific molecular chaperone) [18,19], mis-localisation of GalNAc-transferases from the Golgi to the endoplasmic reticulum (ER) [20], and fluctuations in pH levels [21]. Specifically, biosynthesis of the sTn antigen has been linked to the expression of the sialytransferase ST6GalNAc1, and to mutations in and loss of heterozygosity of the COSMC gene (Figure 1). The biosynthesis of sTn can be mediated in vitro by the specific sialyltransferases ST6GalNAc1 and ST6GalNAc2 [22,23]. However, in a cellular context only ST6GalNAc1 is able to synthesise sTn [17,22], and over-expression of ST6GalNAc1 in gastric, breast and prostate cell lines has been shown to induce expression of sTn, indicating a crucial role for this enzyme in sTn biosynthesis [17,24,25,26].
The enzyme C1Ga1T1 catalyses the common first elongation step of the Tn antigen to produce the core 1 O-glycan structure (T antigen) and requires the COSMC chaperone protein for correct folding and activity [18,27,28,29]. Mutations in and loss of heterozygosity of the COSMC gene have been identified in sTn positive colon and melanoma cell lines, and also in sTn positive cervical cancer tissue [18]. Although these events are thought to be rare in breast and colon cancer [30], up to 40% of cases of pancreatic cancer have been found to have hypermethylation of the COSMC gene, and a corresponding increase in truncated O-glycans [19]. Increased expression of sTn in cancer may also be enhanced by increased synthesis of the O-GalNAc precursor and decreased competition due to reduced core-synthesis.
3. sTn Is an Onco-Foetal Antigen
Monoclonal antibodies specific to sTn have been used to show that overall expression of sTn is rare in normal compared to cancer tissues, leading to sTn being described as an onco-foetal antigen [16]. However, while there are some studies describing sTn detection in foetal organs and amniotic fluids [7,31,32,33], a role for sTn in embryonic development remains to be identified. In adult healthy tissues, low sTn expression has been detected in the salivary gland, oesophagus and stomach, and appears to be restricted to the upper digestive tract [7,34,35]. Expression of sTn has also been linked to inflammatory diseases of the digestive tract and may be regulated by ƴ-interferon inflammatory signals [36,37,38]. sTn is believed to be carried by a number of glycoproteins which are differentially expressed in different cell types [16]. In mucinous cells, sTn is carried by glycoproteins such as mucins which can be either membrane bound and secreted [39,40]. Squamous cells mainly have intensive staining in the cytoplasm and some staining on the cell membrane [10,41]. MUC1 (mucin 1, cell surface associated), CD44 (CD44 molecule, Indian blood group), integrin β1 and osteopontin proteins have been identified as sTn carrier proteins [24,42,43,44], all of which have roles in cell adhesion, migration or chemotaxis. It is thought that the addition of sTn to these glycoproteins may influence protein function and play a role in specific mechanisms involved in tumour development [45].
4. sTn as a Prognostic Biomarker
Detection of serum tumour markers is a non-invasive, simple method for diagnosis and post-surgery analysis. The sTn antigen enters serum through O-glycoprotein shedding or secretion from tumours into the bloodstream. This requires a large amount of tumour mass, which is more likely found in advanced cancers and is therefore associated with poor prognosis. A high level of sTn (>38 U/mL) has been detected in the sera of patients with gastric, colorectal, pancreatic, cervical, endometrial and ovarian cancers [15,46,47,48,49,50,51,52,53], and is associated with tumour size and metastasis [46,47,54]. In prostate cancer, sTn is detected in up to half of all high grade tumours [12,13], and sTn-MUC1 has been correlated with survival outcome and higher serum Prostate Specific Antigen (PSA) levels [55]. A recent study has also linked elevated serum sTn levels to histological grade and lymph node metastasis in endometrial cancer [15].
While sTn has been associated with decreased overall survival of patients, this may depend on the type of cancer studied [16]. For example, in lung cancer, no link to survival was found [56], and in prostate cancer the STGalNAc1 enzyme was found to be up-regulated in primary prostate cancer tissue but significantly down-regulated in metastatic lesions [25]. Studies in these cancer types have been limited, but is hoped that additional investigations will help further determine the prognostic value of sTn in different cancer types.
5. The Role of sTn in Cancer Progression
Truncated O-glycans occur on the majority of epithelial cancers and in many pre-malignant lesions [11,57,58]. A recent study by Radhakrishnan et al. showed that immature truncated O-glycans can directly induce oncogenic features in cancer cells, including increased proliferation, loss of tissue architecture, disruption of basement membrane adhesion and invasive growth [19]. In breast and gastric cancer cell lines, sTn up-regulation is linked to increased tumour growth and metastases in in vivo models [24,42]. Although induction of sTn inhibited the formation of stable tumour masses in prostate cancer, potentially promoting cancer cell dissemination, there was no effect upon metastasis [25]. Consistent with this, sTn positive ovarian cancer cells are more frequently observed at the invasive front of tumours but less often in metastatic lesions [59,60], and breast ductal invasive carcinomas have lower levels of sTn than primary ductal carcinomas [61]. Studies have suggested that sTn can play a role in protecting metastatic cells in the blood stream from degradation by the immune system [62]. Alterations in glycoprotein composition on the cell surface can induce or prevent the recognition by lectin molecules such as selectins, siglecs and galectins, which can play a role in cell–cell and cell–matrix interactions, and may influence cancer progression [63]. As the biosynthesis of sTn gives rise to negative charges, sTn expression is implicated in the interaction of cells with their surrounding environment [64]. It is likely that the sTn-antigen facilitates the release of individual cells from the primary tumour mass by reducing cell–cell aggregation, for example by disrupting the interaction of galectins with terminal galactose residues [65,66]. Thus, in some cases, sTn may enhance the dissemination of cancer cells, promoting primary tumour transition, but may not improve the settlement of metastatic cells at secondary sites [25]. Cells expressing sTn may have an improved ability to migrate and invade underlying tissue and eventually reach blood or lymph vessels [67]. As specific adhesive properties are needed for successful extravasation of metastatic cells and invasion of target tissue, one possibility is that sTn can sometimes play a transient role in cancer progression [25].
Targeting sTn carrier proteins is a potential cancer therapeutic option. It remains open whether sTn might be carried on cancer stem cell marker proteins but as these proteins can be modified with O-glycans this seems a likely possibility [68]. The expression of sTn is generally reported to be heterogeneous within tumours [39,69,70,71,72,73] and could be regulated in the tumour via the expression of carrier proteins. As sTn can be carried by different glycoproteins in different compartments, its role in cancer progression may be different in different tumour types, and the effect of sTn expression on tumour cell invasiveness might be organ specific. Together these findings suggest that the biological function of sTn in promoting cancer development may occur through different mechanisms depending on each cancer type or subtype. Future studies using animal models expressing sTn in multiple cancer types and cell backgrounds will help further elucidate the role in sTn in cancer cell invasion and metastasis in different cancers.
Immunotherapy, or using the patient’s own immune system to target cancer cells, is an attractive approach to treat cancer, and the side-effects are mild compared with traditional therapies. Anti-tumour antibodies could delay tumour growth by antibody dependent cytotoxicity and inhibition of function. Since healthy adult epithelial cells do not normally expose the sTn antigen to the immune system and bloodstream, the use of sTn as an immunisation antigen is an attractive option [16,74,75]. Clinical studies have demonstrated that immunisation with anti-sTn vaccines can induce production of sTn specific IgGs [76,77,78], and in murine models sTn vaccines have been found to induce antibody mediated tumour protection [44]. Additionally, in early stage breast cancer, the detection of antibodies against MUC1-sTn is associated with increased time to metastasis suggesting a protective role of anti-sTn antibodies [79]. sTn antibodies have been detected in healthy females who did not go on to develop cancer 25–30 years after blood sampling, suggesting a possible functional role or these antibodies in suppressing tumour development [79].
The Theratope anti-sTn vaccine produces an anti-sTn immune response and is well tolerated by patients with little toxicity [80,81,82]. In phase II clinical trials for patients with metastatic breast cancer there was a significant improvement in survival by 12 months [80,81,82,83,84]. These promising results led to a randomised, double blind phase III trial involving over 1000 women with metastatic breast cancer, but unfortunately in this trial Theratope failed to demonstrate improved patient survival or time to disease progression [76]. One possible explanation for the failure of the Theratope phase III clinical trial is that the patients were not pre-evaluated for sTn expression, meaning that heterogeneous sTn expression between patients could have masked any benefit of the vaccine [16]. A concurrent study showed that Theratope did increase survival in a pre-stratified subset of patients who were receiving hormonal therapy [85]. The reasons why anti-sTn immunisation seems to only be effective in a subset of patients remain unclear but the overall results point to a relative efficiency and safety of the Theratope vaccine.
In recent years, there has been rapid progress in antibody production methodology, including the immortalisation of human B cells, phage display technology and high throughput screening, transgenic mice and more recently molecular-engineered antibodies [86]. It is hoped these new technologies may have the potential to develop novel antibodies against sTn for use in cancer treatment [86]. An immunotherapy approach targeting sTn carrier proteins may offer the possibility to target more specific mechanisms in tumour development. For example, vaccines targeting MUC1-sTn have been evaluated in clinical trials (Bradbury and Shepherd, 2008, Beatson et al., 2010). MUC1 is expressed on the surface of epithelial cells where it is involved in cell adhesion and signalling (Hollingsworth and Swanson, 2004), and can be aberrantly glycosylated with sTn (Burdick et al., 1997). It may also be possible to create cell therapy via training dendritic cells (DCs) or T cells with sTn or sTn carrier proteins as a tool for cancer therapy. For anti-sTn therapy to advance further it will be vital to fully understand the role that the sTn antigen plays in the function of sTn expressing proteins and in overall tumour progression. Once this is achieved, it may also be possible to combine sTn with other cancer associated antigens in order to trigger multi-antigenic responses in patients to create new platforms for delivery of anti-tumour therapeutics based on the patient’s personal tumour profile (Niederhafner et al., 2008, Slovin, 2007). As the sTn-antigen is expressed early in carcinogenesis in all epithelial cancers investigated, targeting sTn in cancer may enable the targeting of tumours from the earliest stage.
6. Future Perspectives
Many proteins with central roles in cell–cell adhesion, differentiation and development are O-glycosylated and known to have roles in cancer [19,87]. Induction of sTn in cancer could potentially produce global effects on O-glycosylation of proteins, and simultaneously affect multiple systems to promote tumour formation and growth, meaning that deciphering the molecular mechanisms underlying the role of sTn in cancer will not be straightforward. Site specific modification of proteins with O-glycans can have profound effects on protein functions such as pro-protein processing, modulation of ligand binding properties of receptors and regulation of cell signalling [88,89,90]. Future studies determining the specific effects of sTn on specific glycoproteins will help further dissect the role of sTn in cancer.
Acknowledgments
This work was funded by Prostate Cancer UK (PG12-34), The John George William Patterson Foundation, The Wellcome Trust (grant numbers WT080368MA and WT089225/Z/09/Z) and BBSRC (grant numbers BB/1006923/1 and BB/J007293/1).
Conflicts of Interest
The author declares no conflict of interest.
References
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Varki, A. Perspectives on the significance of altered glycosylation of glycoproteins in cancer. Glycoconj. J. 1997, 14, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Tarp, M.A.; Clausen, H. Mucin-type O-glycosylation and its potential use in drug and vaccine development. Biochim. Biophys. Acta 2008, 1780, 546–563. [Google Scholar] [CrossRef] [PubMed]
- Brockhausen, I.; Yang, J.; Dickinson, N.; Ogata, S.; Itzkowitz, S.H. Enzymatic basis for sialyl-Tn expression in human colon cancer cells. Glycoconj. J. 1998, 15, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Ohuchi, N.; Thor, A.; Nose, M.; Fujita, J.; Kyogoku, M.; Schlom, J. Tumor-associated glycoprotein (TAG-72) detected in adenocarcinomas and benign lesions of the stomach. Int. J. Cancer 1986, 38, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Itzkowitz, S.H.; Yuan, M.; Montgomery, C.K.; Kjeldsen, T.; Takahashi, H.K.; Bigbee, W.L.; Kim, Y.S. Expression of Tn, sialosyl-Tn, and T antigens in human colon cancer. Cancer Res. 1989, 49, 197–204. [Google Scholar] [PubMed]
- Thor, A.; Ohuchi, N.; Szpak, C.A.; Johnston, W.W.; Schlom, J. Distribution of oncofetal antigen tumor-associated glycoprotein-72 defined by monoclonal antibody B72.3. Cancer Res. 1986, 46, 3118–3124. [Google Scholar] [PubMed]
- Lottich, S.C.; Johnston, W.W.; Szpak, C.A.; Delong, E.R.; Thor, A.; Schlom, J. Tumor-associated antigen TAG-72: Correlation of expression in primary and metastatic breast carcinoma lesions. Breast Cancer Res. Treat. 1985, 6, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Johnston, W.W.; Szpak, C.A.; Thor, A.; Schlom, J. Phenotypic characterization of lung cancers in fine needle aspiration biopsies using monoclonal antibody B72.3. Cancer Res. 1986, 46, 6462–6470. [Google Scholar] [PubMed]
- Flucke, U.; Zirbes, T.K.; Schroder, W.; Monig, S.P.; Koch, V.; Schmitz, K.; Thiele, J.; Dienes, H.P.; Holscher, A.H.; Baldus, S.E. Expression of mucin-associated carbohydrate core antigens in esophageal squamous cell carcinomas. Anticancer Res. 2001, 21, 2189–2193. [Google Scholar] [PubMed]
- Ching, C.K.; Holmes, S.W.; Holmes, G.K.; Long, R.G. Comparison of two sialosyl-Tn binding monoclonal antibodies (MLS102 and B72.3) in detecting pancreatic cancer. Gut 1993, 34, 1722–1725. [Google Scholar] [CrossRef] [PubMed]
- Genega, E.M.; Hutchinson, B.; Reuter, V.E.; Gaudin, P.B. Immunophenotype of high-grade prostatic adenocarcinoma and urothelial carcinoma. Modern Pathol. 2000, 13, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.B.; Meredith, R.F.; Schlom, J.; LoBuglio, A.F.; Bueschen, A.J.; Wheeler, R.H.; Stockard, C.R.; Grizzle, W.E. Tumor associated glycoprotein-72 is highly expressed in prostatic adenocarcinomas. J. Urol. 1994, 152, 243–246. [Google Scholar] [PubMed]
- Ferreira, J.A.; Videira, P.A.; Lima, L.; Pereira, S.; Silva, M.; Carrascal, M.; Severino, P.F.; Fernandes, E.; Almeida, A.; Costa, C.; et al. Overexpression of tumour-associated carbohydrate antigen sialyl-Tn in advanced bladder tumours. Mol. Oncol. 2013, 7, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Kasai, M.; Fukuda, T.; Ichimura, T.; Yasui, T.; Sumi, T. Serum sialyl-Tn (sTn) as a tumor marker in patients with endometrial cancer. Pathol. Oncol. Res. POR 2015. [Google Scholar] [CrossRef] [PubMed]
- Julien, S.; Videira, P.A.; Delannoy, P. Sialyl-Tn in cancer: (How) did we miss the target? Biomolecules 2012, 2, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Sewell, R.; Backstrom, M.; Dalziel, M.; Gschmeissner, S.; Karlsson, H.; Noll, T.; Gatgens, J.; Clausen, H.; Hansson, G.C.; Burchell, J.; et al. The ST6GalNAc-I sialyltransferase localizes throughout the Golgi and is responsible for the synthesis of the tumor-associated sialyl-Tn O-glycan in human breast cancer. J. Biol. Chem. 2006, 281, 3586–3594. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Lanneau, G.S.; Gautam, T.; Wang, Y.; Xia, B.; Stowell, S.R.; Willard, M.T.; Wang, W.; Xia, J.Y.; Zuna, R.E.; et al. Human tumor antigens Tn and sialyl Tn arise from mutations in COSMC. Cancer Res. 2008, 68, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, P.; Dabelsteen, S.; Madsen, F.B.; Francavilla, C.; Kopp, K.L.; Steentoft, C.; Vakhrushev, S.Y.; Olsen, J.V.; Hansen, L.; Bennett, E.P.; et al. Immature truncated O-glycophenotype of cancer directly induces oncogenic features. Proc. Natl. Acad. Sci. USA 2014, 111, E4066–E4075. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.J.; Chia, J.; Senewiratne, J.; Bard, F. Regulation of O-glycosylation through Golgi-to-ER relocation of initiation enzymes. J. Cell Biol. 2010, 189, 843–858. [Google Scholar] [CrossRef] [PubMed]
- Hassinen, A.; Pujol, F.M.; Kokkonen, N.; Pieters, C.; Kihlstrom, M.; Korhonen, K.; Kellokumpu, S. Functional organization of Golgi N- and O-glycosylation pathways involves pH-dependent complex formation that is impaired in cancer cells. J. Biol. Chem. 2011, 286, 38329–38340. [Google Scholar] [CrossRef] [PubMed]
- Marcos, N.T.; Bennett, E.P.; Gomes, J.; Magalhaes, A.; Gomes, C.; David, L.; Dar, I.; Jeanneau, C.; DeFrees, S.; Krustrup, D.; et al. ST6GalNAc-I controls expression of sialyl-Tn antigen in gastrointestinal tissues. Front. Biosci. 2011, 3, 1443–1455. [Google Scholar] [CrossRef]
- Harduin-Lepers, A.; Vallejo-Ruiz, V.; Krzewinski-Recchi, M.A.; Samyn-Petit, B.; Julien, S.; Delannoy, P. The human sialyltransferase family. Biochimie 2001, 83, 727–737. [Google Scholar] [CrossRef]
- Julien, S.; Adriaenssens, E.; Ottenberg, K.; Furlan, A.; Courtand, G.; Vercoutter-Edouart, A.S.; Hanisch, F.G.; Delannoy, P.; Le Bourhis, X. ST6GalNAc I expression in MDA-MB-231 breast cancer cells greatly modifies their O-glycosylation pattern and enhances their tumourigenicity. Glycobiology 2006, 16, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J.; Oltean, S.; Vodak, D.; Wilson, B.T.; Livermore, K.E.; Zhou, Y.; Star, E.; Floros, V.I.; Johannessen, B.; Knight, B.; et al. The androgen receptor controls expression of the cancer-associated sTn antigen and cell adhesion through induction of ST6GalNAc1 in prostate cancer. Oncotarget 2015, 6, 34358–34374. [Google Scholar] [PubMed]
- Marcos, N.T.; Pinho, S.; Grandela, C.; Cruz, A.; Samyn-Petit, B.; Harduin-Lepers, A.; Almeida, R.; Silva, F.; Morais, V.; Costa, J.; et al. Role of the human ST6GalNAc-I and ST6GalNAc-II in the synthesis of the cancer-associated sialyl-Tn antigen. Cancer Res. 2004, 64, 7050–7057. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Aryal, R.P.; Stowell, C.J.; Cummings, R.D. Regulation of protein O-glycosylation by the endoplasmic reticulum-localized molecular chaperone COSMC. J. Cell Biol. 2008, 182, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Cummings, R.D. A unique molecular chaperone COSMC required for activity of the mammalian core 1 β3-galactosyltransferase. Proc. Natl. Acad. Sci. USA 2002, 99, 16613–16618. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ju, T.; Ding, X.; Xia, B.; Wang, W.; Xia, L.; He, M.; Cummings, R.D. COSMC is an essential chaperone for correct protein O-glycosylation. Proc. Natl. Acad. Sci. USA 2010, 107, 9228–9233. [Google Scholar] [CrossRef] [PubMed]
- Yoo, N.J.; Kim, M.S.; Lee, S.H. Absence of COSMC gene mutations in breast and colorectal carcinomas. APMIS 2008, 116, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Stanick, D.; Schuss, A.; Mishriki, Y.; Chao, S.; Thor, A.; Lundy, J. Reactivity of the monoclonal antibody B72.3 with fetal antigen: Correlation with expression of TAG-72 in human carcinomas. Cancer Investig. 1988, 6, 279–287. [Google Scholar] [CrossRef]
- Pistolesi, S.; Nuti, M.; Castagna, M.; Pingitore, R. Mammary fetal gland: Identification of new oncofetal antigens by monoclonal antibodies B72.3, MM1.80 and 4.36. Tumori 2001, 87, 252–255. [Google Scholar] [PubMed]
- Baldus, S.E.; Hanisch, F.G.; Schwonzen, M.; Nakahara, Y.; Iijima, H.; Ogawa, T.; Peter-Katalinic, J.; Uhlenbruck, G. Monoclonal antibody SP-21 defines a sialosyl-Tn antigen expressed on carcinomas and K562 erythroleukemia cells. Anticancer Res. 1992, 12, 1935–1940. [Google Scholar] [PubMed]
- Cao, Y.; Stosiek, P.; Springer, G.F.; Karsten, U. Thomsen-friedenreich-related carbohydrate antigens in normal adult human tissues: A systematic and comparative study. Histochem. Cell Biol. 1996, 106, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Yonezawa, S.; Nomoto, M.; Ueno, K.; Kim, Y.S.; Sato, E. Expression of mucin antigens and lewis X-related antigens in carcinomas and dysplasia of the pharynx and larynx. Pathol. Int. 1996, 46, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Ta, A.; Harpaz, N.; Bodian, C.; Roston, A.; Oberman, L.; Chen, A.; Itzkowitz, S. Sialyl-Tn antigen expression in Crohn’s colitis. Inflamm. Bowel Dis. 1997, 3, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Barresi, G.; Giuffre, G.; Vitarelli, E.; Grosso, M.; Tuccari, G. The immunoexpression of Tn, sialyl-Tn and T antigens in chronic active gastritis in relation to helicobacter pylori infection. Pathology 2001, 33, 298–302. [Google Scholar] [PubMed]
- Cohen, M.; Drut, R.; Cueto Rua, E. Sialyl-Tn antigen distribution in helicobacter pylori chronic gastritis in children: An immunohistochemical study. Pediatr. Pathol. Mol. Med. 2003, 22, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Federici, M.F.; Kudryashov, V.; Saigo, P.E.; Finstad, C.L.; Lloyd, K.O. Selection of carbohydrate antigens in human epithelial ovarian cancers as targets for immunotherapy: Serous and mucinous tumors exhibit distinctive patterns of expression. Int. J. Cancer 1999, 81, 193–198. [Google Scholar] [CrossRef]
- Pinto, R.; Carvalho, A.S.; Conze, T.; Magalhaes, A.; Picco, G.; Burchell, J.M.; Taylor-Papadimitriou, J.; Reis, C.A.; Almeida, R.; Mandel, U.; et al. Identification of new cancer biomarkers based on aberrant mucin glycoforms by in situ proximity ligation. J. Cell. Mol. Med. 2012, 16, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ferrer, A.; Barranco, C.; de Bolos, C. Differences in the O-glycosylation patterns between lung squamous cell carcinoma and adenocarcinoma. Am. J. Clin. Pathol. 2002, 118, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, H.; Matsuzaki, H.; Ando, H.; Kaji, H.; Nakanishi, H.; Ikehara, Y.; Narimatsu, H. Enhancement of metastatic ability by ectopic expression of ST6GalNAcI on a gastric cancer cell line in a mouse model. Clin. Exp. Metastasis 2012, 29, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Rocher, J.; Loirand, G.; Le Pendu, J. Expression of sialyl-Tn epitopes on β1 integrin alters epithelial cell phenotype, proliferation and haptotaxis. J. Cell Sci. 2004, 117, 5059–5069. [Google Scholar] [CrossRef] [PubMed]
- Julien, S.; Picco, G.; Sewell, R.; Vercoutter-Edouart, A.S.; Tarp, M.; Miles, D.; Taylor-Papadimitriou, J.; Clausen, H.; Burchell, J.M. Sialyl-Tn vaccine induces antibody-mediated tumour protection in a relevant murine model. Br. J. Cancer 2009, 100, 1746–1754. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.J.; Swindall, A.F.; Bellis, S.L. Regulation of the metastatic cell phenotype by sialylated glycans. Cancer Metastasis Rev. 2012, 31, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, I.; Maehara, Y.; Kusumoto, T.; Yoshida, M.; Kakeji, Y.; Kusumoto, H.; Furusawa, M.; Sugimachi, K. Predictive value of preoperative serum sialyl Tn antigen levels in prognosis of patients with gastric cancer. Cancer 1993, 72, 1836–1840. [Google Scholar] [CrossRef]
- Nakagoe, T.; Sawai, T.; Tsuji, T.; Jibiki, M.A.; Nanashima, A.; Yamaguchi, H.; Yasutake, T.; Ayabe, H.; Arisawa, K.; Ishikawa, H. Predictive factors for preoperative serum levels of sialy lewis(x), sialyl lewis(a) and sialyl Tn antigens in gastric cancer patients. Anticancer Res. 2002, 22, 451–458. [Google Scholar] [PubMed]
- Motoo, Y.; Kawakami, H.; Watanabe, H.; Satomura, Y.; Ohta, H.; Okai, T.; Makino, H.; Toya, D.; Sawabu, N. Serum sialyl-Tn antigen levels in patients with digestive cancers. Oncology 1991, 48, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Nishimura, G.; Nonomura, A.; Miwa, K.; Miyazaki, I. Serological studies on CEA, CA 19–9, sTn and SLX in colorectal cancer. Hepatogastroenterology 1999, 46, 914–919. [Google Scholar] [PubMed]
- Nanashima, A.; Yamaguchi, H.; Nakagoe, T.; Matsuo, S.; Sumida, Y.; Tsuji, T.; Sawai, T.; Yamaguchi, E.; Yasutake, T.; Ayabe, H. High serum concentrations of sialyl Tn antigen in carcinomas of the biliary tract and pancreas. J. Hepato-Biliary-Pancreat. Surg. 1999, 6, 391–395. [Google Scholar] [CrossRef]
- Numa, F.; Tsunaga, N.; Michioka, T.; Nawata, S.; Ogata, H.; Kato, H. Tissue expression of sialyl Tn antigen in gynecologic tumors. J. Obstet. Gynaecol. 1995, 21, 385–389. [Google Scholar] [CrossRef]
- Kobayashi, H.; Terao, T.; Kawashima, Y. Sialyl Tn as a prognostic marker in epithelial ovarian cancer. Br. J. Cancer 1992, 66, 984–985. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Ogawa, H.; Nakanishi, K.; Tanizawa, O.; Karino, K.; Endo, J. Clinical value of sialyl Tn antigen in patients with gynecologic tumors. Obstet. Gynecol. 1990, 75, 1032–1036. [Google Scholar] [PubMed]
- Imada, T.; Rino, Y.; Takahashi, M.; Hatori, S.; Shiozawa, M.; Amano, T.; Kondo, J. Serum CA 19–9, SLX, sTn and CEA levels of the peripheral and the draining venous blood in gastric cancer. Hepatogastroenterology 1999, 46, 2086–2090. [Google Scholar] [PubMed]
- Arai, T.; Fujita, K.; Fujime, M.; Irimura, T. Expression of sialylated muc1 in prostate cancer: Relationship to clinical stage and prognosis. Int. J. Urol. 2005, 12, 654–661. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, T.A.; Aloia, T.A.; Moore, M.B.; Herndon, J.E., II; Brooks, K.R.; Lau, C.L.; Harpole, D.H., Jr. Molecular biologic substaging of stage i lung cancer according to gender and histology. Ann. Thorac. Surg. 2000, 69, 882–886. [Google Scholar]
- Itzkowitz, S.; Kjeldsen, T.; Friera, A.; Hakomori, S.; Yang, U.S.; Kim, Y.S. Expression of Tn, sialosyl Tn, and T antigens in human pancreas. Gastroenterology 1991, 100, 1691–1700. [Google Scholar] [PubMed]
- Kim, G.E.; Bae, H.I.; Park, H.U.; Kuan, S.F.; Crawley, S.C.; Ho, J.J.; Kim, Y.S. Aberrant expression of MUC5AC and MUC6 gastric mucins and sialyl Tn antigen in intraepithelial neoplasms of the pancreas. Gastroenterology 2002, 123, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Berner, A.; Nesland, J.M.; Risberg, B.; Kristensen, G.B.; Trope, C.G.; Bryne, M. Carbohydrate antigen expression in primary tumors, metastatic lesions, and serous effusions from patients diagnosed with epithelial ovarian carcinoma: Evidence of up-regulated Tn and sialyl Tn antigen expression in effusions. Hum. Pathol. 2000, 31, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Gotlieb, W.H.; Ben-Baruch, G.; Kopolovic, J.; Goldberg, I.; Nesland, J.M.; Berner, A.; Bjamer, A.; Bryne, M. Expression of carbohydrate antigens in advanced-stage ovarian carcinomas and their metastases—A clinicopathologic study. Gynecol. Oncol. 2000, 77, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, F.C.; Figueiredo, P.; Lacerda, M. Simple mucin-type carbohydrate antigens (T, sialosyl-T, Tn and sialosyl-Tn) in breast carcinogenesis. Virchows Arch. 1995, 427, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Ogata, S.; Maimonis, P.J.; Itzkowitz, S.H. Mucins bearing the cancer-associated sialosyl-Tn antigen mediate inhibition of natural killer cell cytotoxicity. Cancer Res. 1992, 52, 4741–4746. [Google Scholar] [PubMed]
- Hauselmann, I.; Borsig, L. Altered tumor-cell glycosylation promotes metastasis. Front. Oncol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J.; Elliott, D.J. Sugars and cell adhesion: The role of ST6GalNAc1 in prostate cancer progression. Cancer Cell Microenviron. 2016, 3, e1174. [Google Scholar]
- Takenaka, Y.; Fukumori, T.; Raz, A. Galectin-3 and metastasis. Glycoconj. J. 2004, 19, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Beatson, R.; Maurstad, G.; Picco, G.; Arulappu, A.; Coleman, J.; Wandell, H.H.; Clausen, H.; Mandel, U.; Taylor-Papadimitriou, J.; Sletmoen, M.; et al. The breast cancer-associated glycoforms of MUC1, MUC1-Tn and sialyl-Tn, are expressed in COSMC wild-type cells and bind the C-type lectin MGL. PLoS ONE 2015, 10, e0125994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinho, S.; Marcos, N.T.; Ferreira, B.; Carvalho, A.S.; Oliveira, M.J.; Santos-Silva, F.; Harduin-Lepers, A.; Reis, C.A. Biological significance of cancer-associated sialyl-Tn antigen: Modulation of malignant phenotype in gastric carcinoma cells. Cancer Lett. 2007, 249, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U.; Goletz, S. What makes cancer stem cell markers different? SpringerPlus 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Victorzon, M.; Nordling, S.; Nilsson, O.; Roberts, P.J.; Haglund, C. Sialyl Tn antigen is an independent predictor of outcome in patients with gastric cancer. Int. J. Cancer 1996, 65, 295–300. [Google Scholar] [CrossRef]
- Cao, Y.; Schlag, P.M.; Karsten, U. Immunodetection of epithelial mucin (MUC1, MUC3) and mucin-associated glycotopes (TF, Tn, and sialosyl-Tn) in benign and malignant lesions of colonic epithelium: Apolar localization corresponds to malignant transformation. Virchows Arch. 1997, 431, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Carrilho, C.; Cantel, M.; Gouveia, P.; David, L. Simple mucin-type carbohydrate antigens (Tn, sialosyl-Tn, T and sialosyl-T) and gp 230 mucin-like glycoprotein are candidate markers for neoplastic transformation of the human cervix. Virchows Arch. 2000, 437, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Kinney, A.Y.; Sahin, A.; Vernon, S.W.; Frankowski, R.F.; Annegers, J.F.; Hortobagyi, G.N.; Buzdar, A.U.; Frye, D.K.; Dhingra, K. The prognostic significance of sialyl-Tn antigen in women treated with breast carcinoma treated with adjuvant chemotherapy. Cancer 1997, 80, 2240–2249. [Google Scholar] [CrossRef]
- Miles, D.W.; Happerfield, L.C.; Smith, P.; Gillibrand, R.; Bobrow, L.G.; Gregory, W.M.; Rubens, R.D. Expression of sialyl-Tn predicts the effect of adjuvant chemotherapy in node-positive breast cancer. Br. J. Cancer 1994, 70, 1272–1275. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, L.A.; Sandmaier, B.M. Theratope vaccine (sTn-KLH). Expert Opin. Biol. Ther. 2001, 1, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.A. Technology evaluation: Theratope, biomira inc. Curr. Opin. Mol. Ther. 2000, 2, 453–458. [Google Scholar] [PubMed]
- Miles, D.; Roche, H.; Martin, M.; Perren, T.J.; Cameron, D.A.; Glaspy, J.; Dodwell, D.; Parker, J.; Mayordomo, J.; Tres, A.; et al. Phase III multicenter clinical trial of the sialyl-Tn (sTn)-keyhole limpet hemocyanin (KLH) vaccine for metastatic breast cancer. Oncologist 2011, 16, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.K.; Murray, J.L. Clinical development of the sTn-KLH vaccine (Theratope®). Clin. Breast Cancer 2003, 3 (Suppl. 4), S139–S143. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.; Papazisis, K. Rationale for the clinical development of sTn-KLH (Theratope®) and anti-MUC-1 vaccines in breast cancer. Clin. Breast Cancer 2003, 3 (Suppl. 4), S134–S138. [Google Scholar] [CrossRef] [PubMed]
- Blixt, O.; Bueti, D.; Burford, B.; Allen, D.; Julien, S.; Hollingsworth, M.; Gammerman, A.; Fentiman, I.; Taylor-Papadimitriou, J.; Burchell, J.M. Autoantibodies to aberrantly glycosylated MUC1 in early stage breast cancer are associated with a better prognosis. Breast Cancer Res. 2011, 13, R25. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.W.; Towlson, K.E.; Graham, R.; Reddish, M.; Longenecker, B.M.; Taylor-Papadimitriou, J.; Rubens, R.D. A randomised phase II study of sialyl-Tn and DETOX-B adjuvant with or without cyclophosphamide pretreatment for the active specific immunotherapy of breast cancer. Br. J. Cancer 1996, 74, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- MacLean, G.D.; Reddish, M.A.; Koganty, R.R.; Longenecker, B.M. Antibodies against mucin-associated sialyl-Tn epitopes correlate with survival of metastatic adenocarcinoma patients undergoing active specific immunotherapy with synthetic sTn vaccine. J. Immunother. Emphas. Tumor Immunol. 1996, 19, 59–68. [Google Scholar] [CrossRef]
- Holmberg, L.A.; Sandmaier, B.M. Vaccination with Theratope® (sTn-KLH) as treatment for breast cancer. Expert Rev. Vaccines 2004, 3, 655–663. [Google Scholar] [CrossRef] [PubMed]
- MacLean, G.D.; Miles, D.W.; Rubens, R.D.; Reddish, M.A.; Longenecker, B.M. Enhancing the effect of Theratope® sTn-KLH cancer vaccine in patients with metastatic breast cancer by pretreatment with low-dose intravenous cyclophosphamide. J. Immunother. Emphas. Tumor Immunol. 1996, 19, 309–316. [Google Scholar] [CrossRef]
- Reddish, M.A.; MacLean, G.D.; Poppema, S.; Berg, A.; Longenecker, B.M. Pre-immunotherapy serum CA27.29 (MUC-1) mucin level and CD69+ lymphocytes correlate with effects of Theratope® sialyl-Tn-KLH cancer vaccine in active specific immunotherapy. Cancer Immunol. Immunother. 1996, 42, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Mayordomo, J.; Tres, A.; Miles, D.; Finke, L.; Jenkins, H. Long-term follow-up of patients concomitantly treated with hormone therapy in a prospective controlled randomized multicenter clinical study comparing sTn-KLH vaccine with KLH control in stage IV breast cancer following first-line chemotherapy. J. Clin. Oncol. 2004, 22, 2603. [Google Scholar]
- Loureiro, L.R.; Carrascal, M.A.; Barbas, A.; Ramalho, J.S.; Novo, C.; Delannoy, P.; Videira, P.A. Challenges in antibody development against Tn and sialyl-Tn antigens. Biomolecules 2015, 5, 1783–1809. [Google Scholar] [CrossRef] [PubMed]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, K.T.; Clausen, H. Site-specific protein O-glycosylation modulates proprotein processing—Deciphering specific functions of the large polypeptide galnac-transferase gene family. Biochim. Biophys. Acta 2012, 1820, 2079–2094. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, K.; Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Biosynthesis of the Sialyl-Tn (sTn) antigen in cancer cells.

© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030275
AMA Style
Munkley J. The Role of Sialyl-Tn in Cancer. International Journal of Molecular Sciences. 2016; 17(3):275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030275
Chicago/Turabian StyleMunkley, Jennifer. 2016. "The Role of Sialyl-Tn in Cancer" International Journal of Molecular Sciences 17, no. 3: 275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030275
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.