
Effects of Two Sublethal Concentrations of Mercury Chloride on the Morphology and Metallothionein Activity in the Liver of Zebrafish (Danio rerio)

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
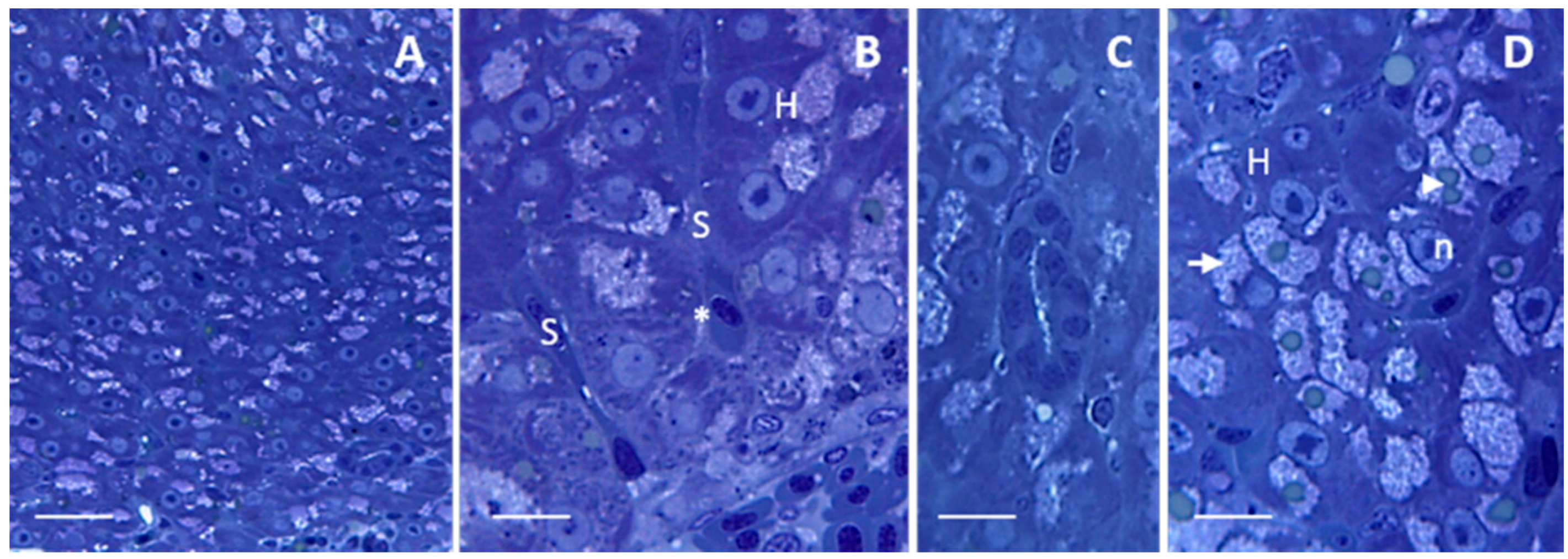
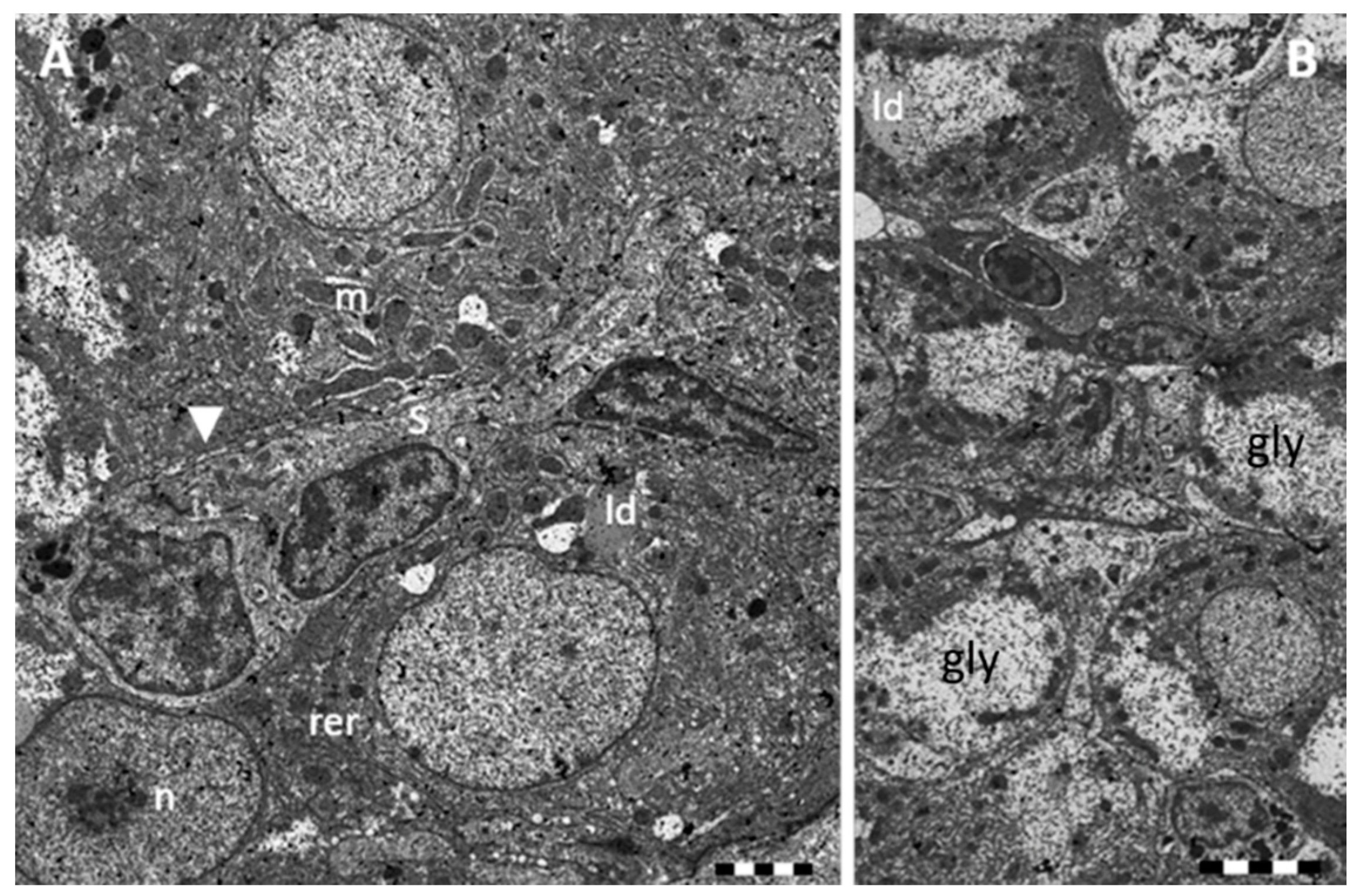
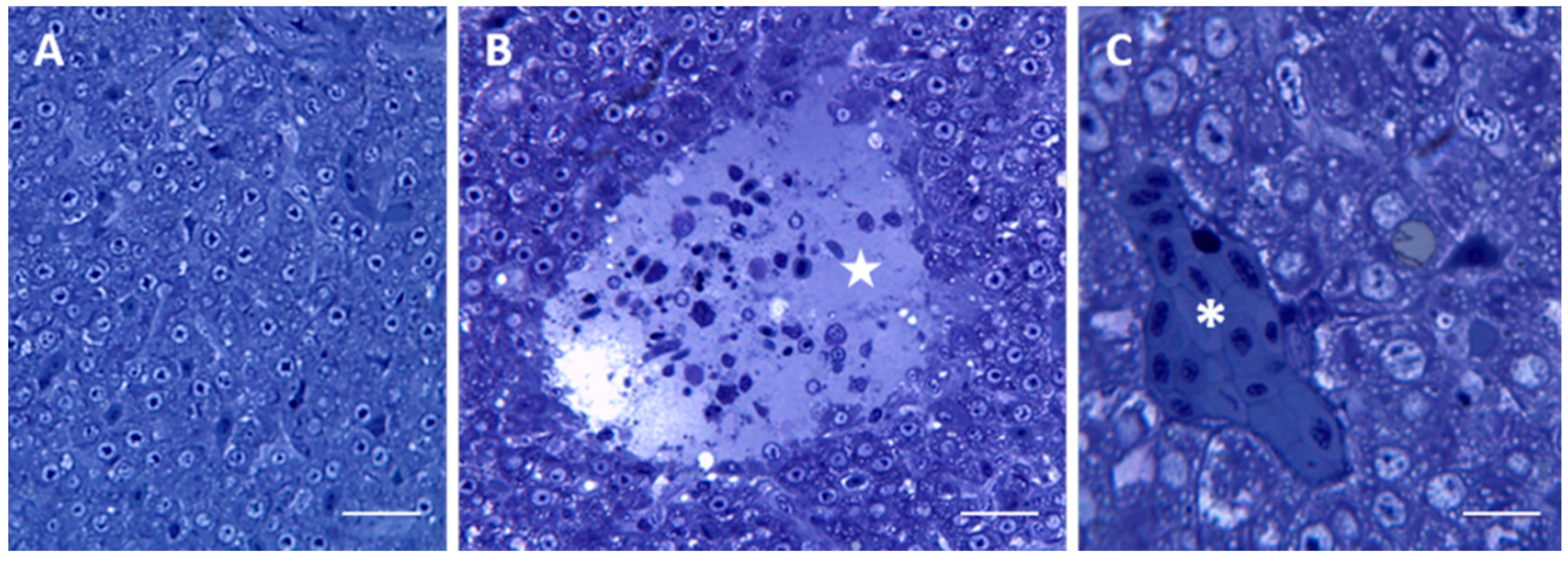
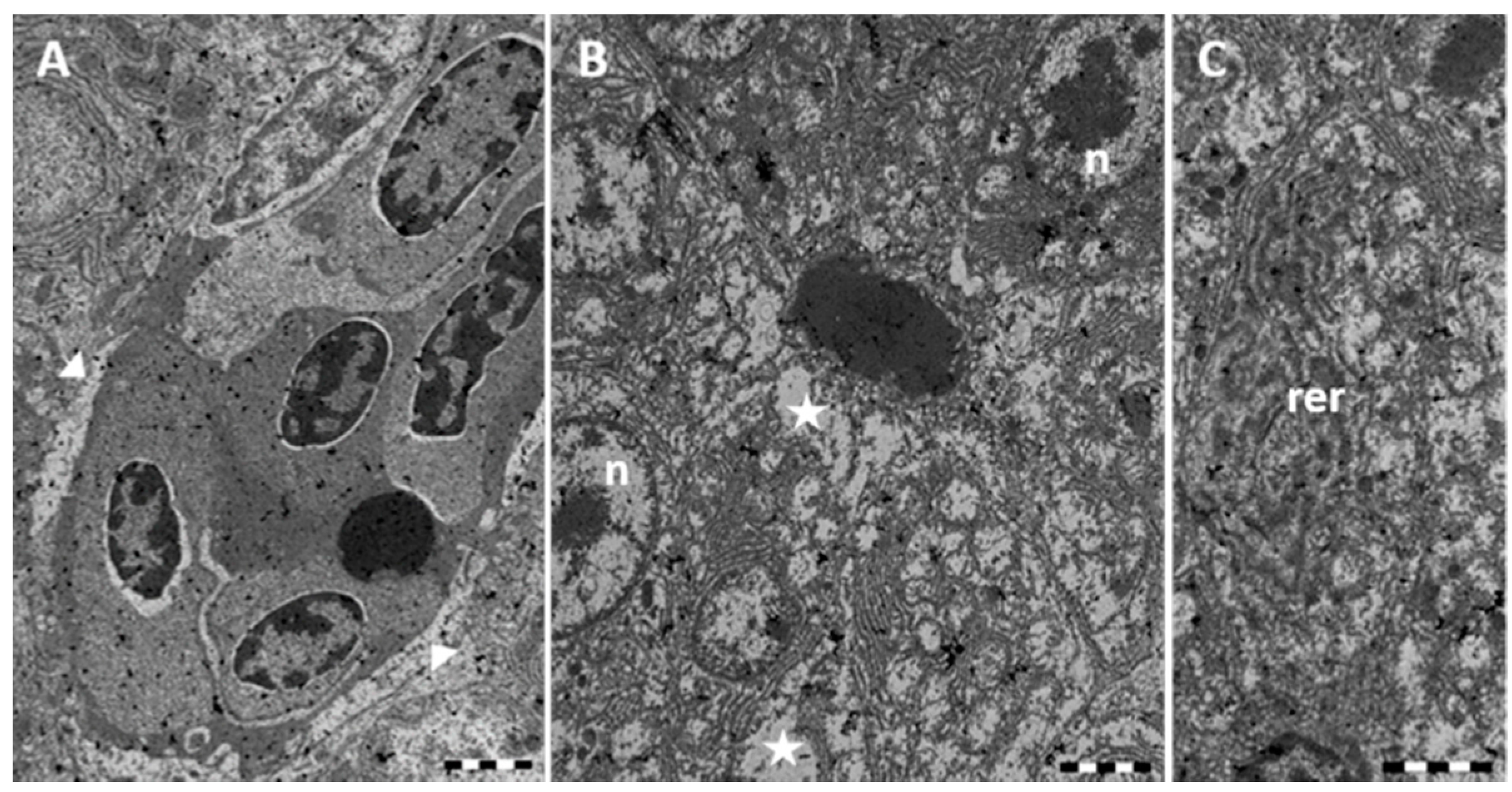
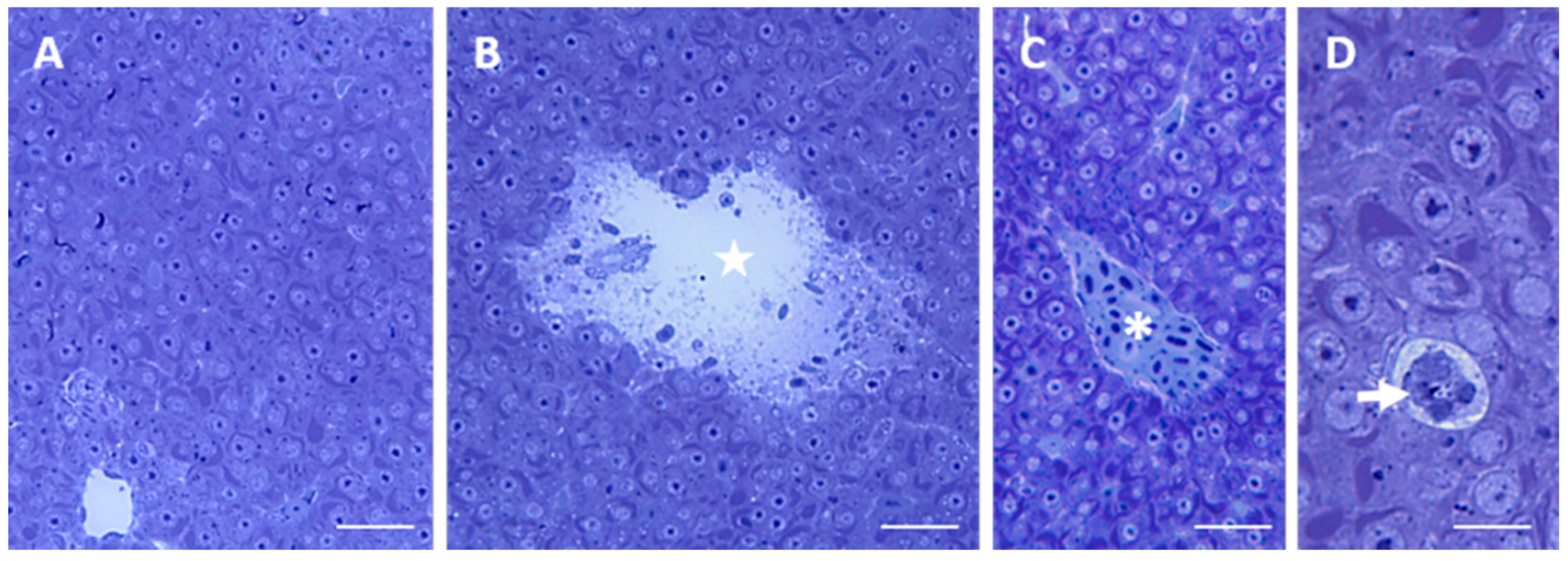
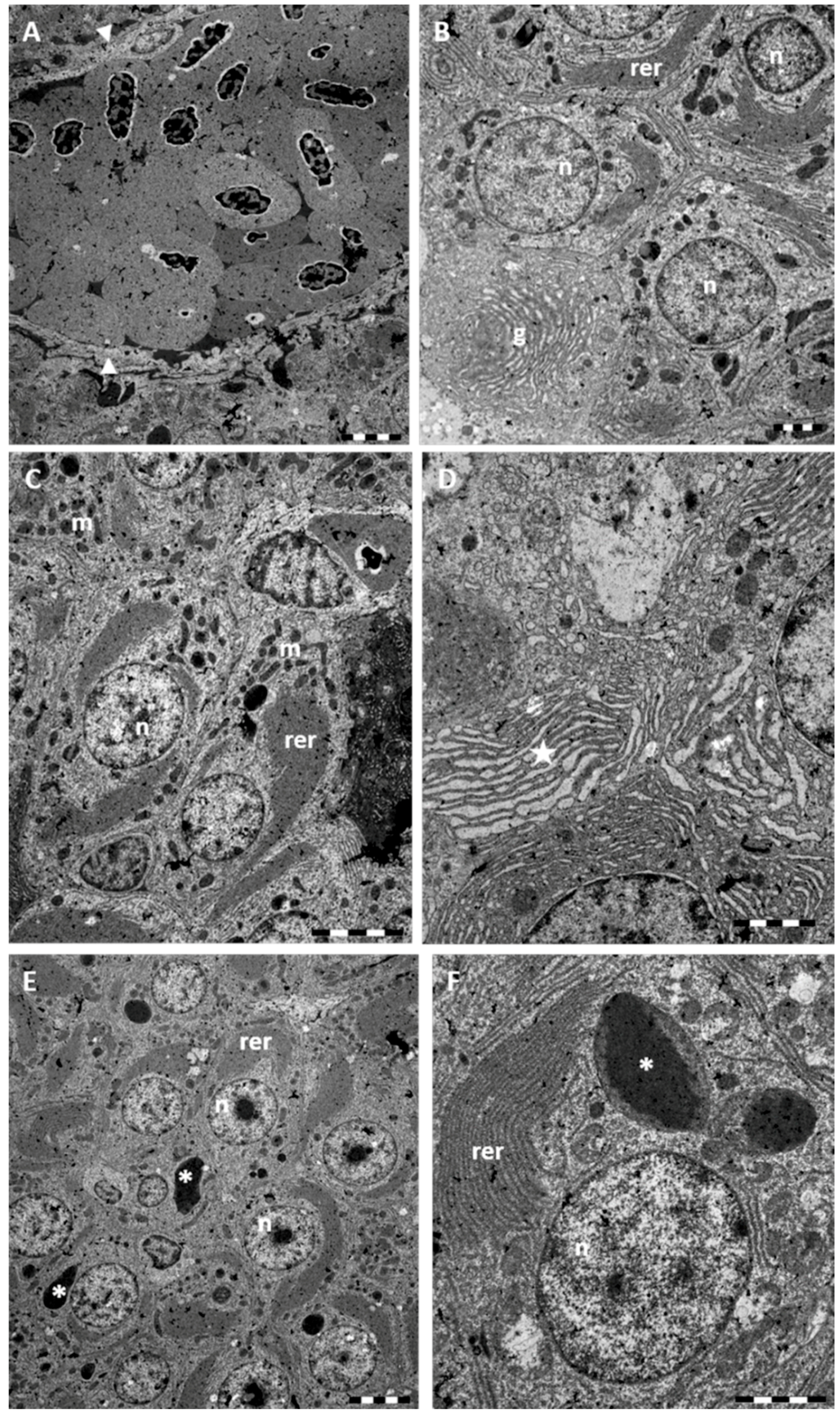
2.1. Morphology and Ultrastructure
2.1.1. Control
2.1.2. Exposed Fishes
2.2. In Situ Hybridization
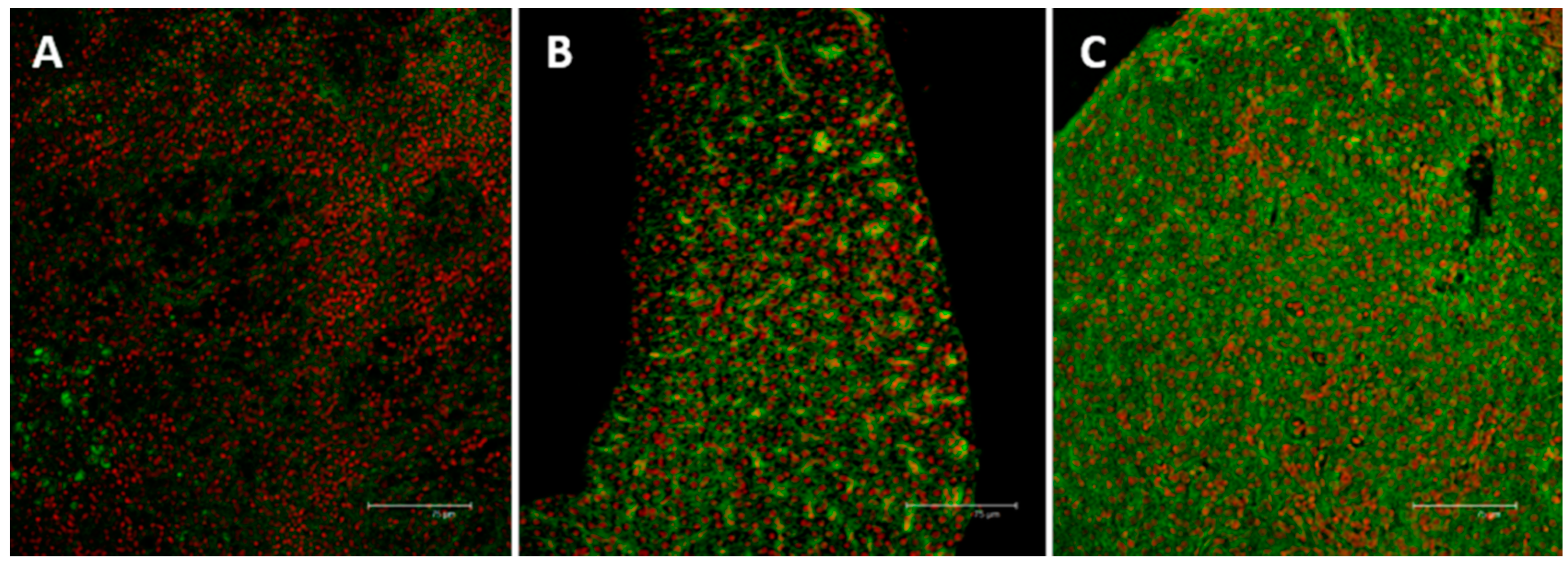
2.3. Immunolocalization of MTs
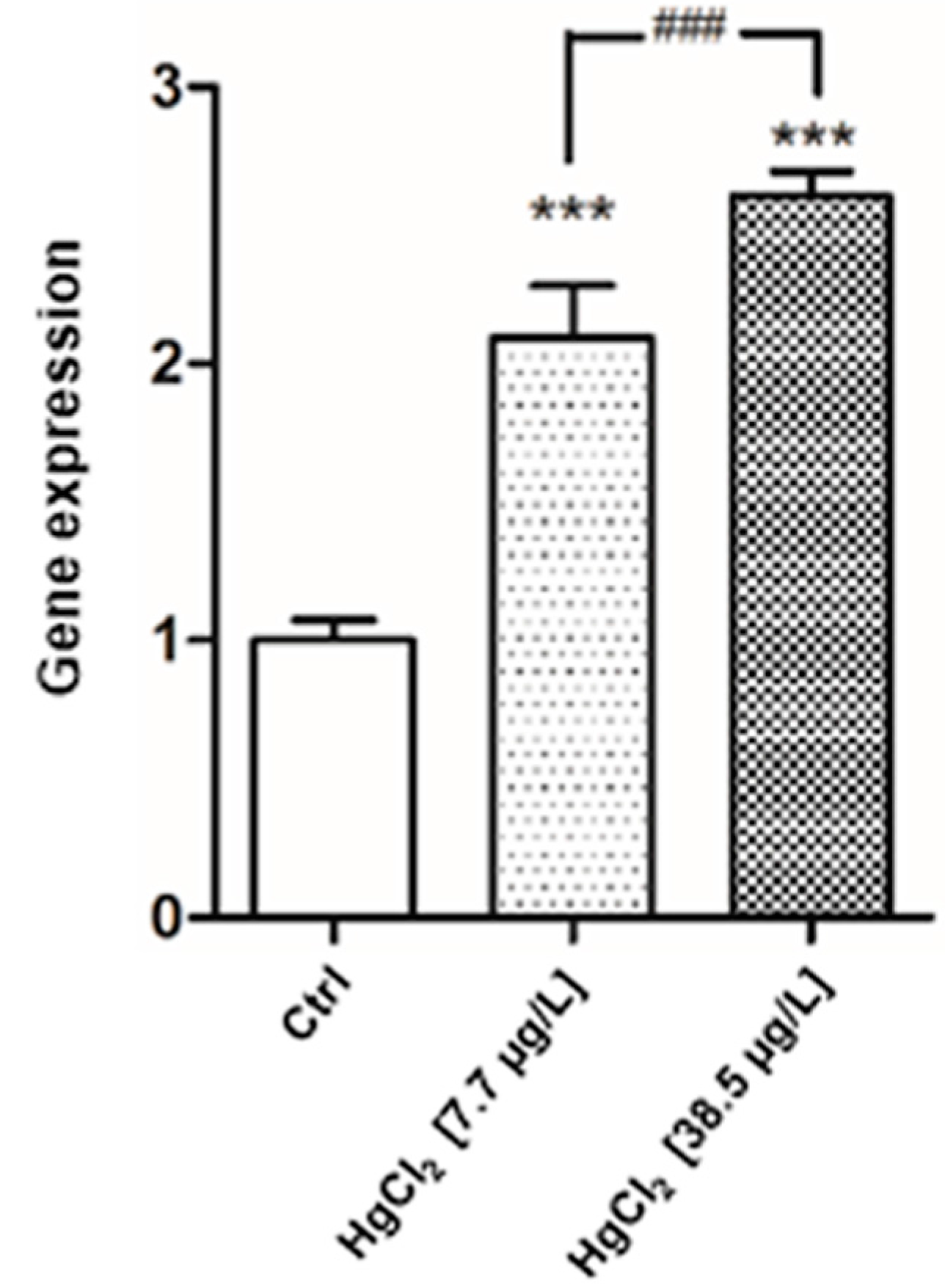
2.4. Gene Expression
3. Discussion
4. Materials and Methods
4.1. Test Organism and Experimental Design
4.2. Light Microscopy and Transmission Electron Microscopy
4.3. Immunohistochemistry
4.4. Mt mRNA in situ Hybridization
4.5. Quantitative RT-PCR
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, G.; Cai, Y.; O’Driscoll, N.; Feng, X.; Jiang, G. Overview of mercury in the environment. In Environmental Chemistry and Toxicology of Mercury, 1st ed.; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 1–12. [Google Scholar]
- Chen, C.W.; Chen, C.F.; Dong, C.D. Distribution and accumulation of mercury in sediments of Kaohsiung River Mouth, Taiwan. APCBEE Procedia 2012, 1, 153–158. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a global pollutant: Sources, pathways, and effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef] [PubMed]
- Wiener, J.G. Mercury exposed: Advances in environmental analysis and ecotoxicology of a highly toxic metal. Environ. Toxicol. Chem. 2013, 32, 2175–2178. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organization). Mercury and health, 2013. Available online: http://www.who.int/mediacentre/factsheets/fs361/en/ (accessed on 11 October 2013).
- Sharma, M.K.; Sharma, A.; Kumar, A.; Kumar, M. Evaluation of protective efficacy of Spirulina fusiformis against mercury induced nephrotoxicity in Swiss albino mice. Food Chem. Toxicol. 2007, 45, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, T.W.; Magos, L.; Myers, G.J. The toxicology of mercury—Current exposures and clinical manifestations. N. Engl. J. Med. 2003, 349, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Bando, I.; Reus, M.I.S.; Andrés, D.; Cascales, M. Endogenous antioxidant defence system in rat liver following mercury chloride oral intoxication. J. Biochem. Mol. Toxic. 2003, 19, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Gentès, S.; Maury-Brachet, R.; Feng, C.; Pedrero, Z.; Tessier, E.; Legeay, A.; Mesmer-Dudons, N.; Baudrimont, M.; Maurice, L.; Amouroux, D.; et al. Specific effects of dietary methylmercury and inorganic mercury in zebrafish (Danio rerio) determined by genetic, histological, and metallothionein responses. Environ. Sci. Technol. 2015, 49, 14560–14569. [Google Scholar] [CrossRef]
- Rao, M.V.; Chhunchha, B. Protective role of melatonin against the mercury induced oxidative stress in the rat thyroid. Food Chem. Toxicol. 2010, 48, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Senger, M.R.; Rosemberg, D.B.; Seibt, K.J.; Dias, R.D.; Bogo, M.R.; Bonan, C.D. Influence of mercury chloride on adenosine deaminase activity and gene expression in zebrafish (Danio rerio) brain. Neurotoxicology 2010, 31, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.A.; Moussa, E.A.; Abbott, L.C. The effect of methylmercury exposure on early central nervous system development in the zebrafish (Danio rerio) embryo. J. Appl. Toxicol. 2012, 32, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Ho, N.Y.; Yang, L.; Legradi, J.; Armant, O.; Takamiya, M.; Rastegar, S.; Strähle, U. Gene responses in the central nervous system of zebrafish embryos exposed to the neurotoxicant methyl mercury. Environ. Sci. Technol. 2013, 47, 3316–3325. [Google Scholar] [CrossRef] [PubMed]
- Cambier, S.; Benard, G.; Mesmer-Dudons, N.; Gonzalez, P.; Rossignol, R.; Brethes, D.; Bourdineaud, J.P. At environmental doses, dietary methylmercury inhibits mitochondrial energy metabolism in skeletal muscles of the zebrafish (Danio rerio). Int. J. Biochem. Cell B 2009, 41, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Cambier, S.; Gonzalez, P.; Durrieu, G.; Maury-Brachet, R.; Boudou, A.; Bourdineaud, J.P. Serial analysis of gene expression in the skeletal muscles of zebrafish fed with a methylmercury-contaminated diet. Environ. Sci. Technol. 2009, 44, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Aboud, O.A.S.A. Impact of pollution with lead, mercury and cadmium on the immune response of Oreochromis niloticus. N.Y. Sci. J. 2010, 3, 9–16. [Google Scholar]
- Penglase, S.; Hamre, K.; Ellingsen, S. Selenium and mercury have a synergistic negative effect on fish reproduction. Aquat. Toxicol. 2014, 149, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.; Puntervoll, P.; Valdersnes, S.; Goksøyr, A. Responses in the brain proteome of Atlantic cod (Gadus morhua) exposed to methylmercury. Aquat. Toxicol. 2010, 100, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Berntssen, M.H.; Aatland, A.; Handy, R.D. Chronic dietary mercury exposure causes oxidative stress, brain lesions, and altered behaviour in Atlantic salmon (Salmo salar) parr. Aquat. Toxicol. 2003, 65, 55–72. [Google Scholar] [CrossRef]
- Brandão, F.; Cappello, T.; Raimundo, J.; Santos, M.A.; Maisano, M.; Mauceri, A.; Pacheco, M.; Pereira, P. Unravelling the mechanisms of mercury hepatotoxicity in wild fish (Liza aurata) through a triad approach: Bioaccumulation, metabolomic profiles and oxidative stress. Metallomics 2015, 7, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Ung, C.Y.; Lam, S.H.; Hlaing, M.M.; Winata, C.L.; Korzh, S.; Mathavan, S.; Gong, Z. Mercury-induced hepatotoxicity in zebrafish: In vivo mechanistic insights from transcriptome analysis, phenotype anchoring and targeted gene expression validation. BMC Genom. 2010, 11, 212. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.; Dominique, Y.; Massabuau, J.C.; Boudou, A.; Bourdineaud, J.P. Comparative effects of dietary methylmercury on gene expression in liver, skeletal muscle, and brain of the zebrafish (Danio rerio). Environ. Sci. Technol. 2005, 39, 3972–3980. [Google Scholar] [CrossRef] [PubMed]
- Mela, M.; Neto, F.F.; Yamamoto, F.Y.; Almeida, R.; Grötzner, S.R.; Ventura, D.F.; de Oliveira Ribeiro, C.A. Mercury distribution in target organs and biochemical responses after subchronic and trophic exposure to Neotropical fish Hoplias malabaricus. Fish Physiol. Biochem. 2014, 40, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Bleau, H.; Daniel, C.; Chevalier, G.; van Tra, H.; Hontela, A. Effects of acute exposure to mercuric chloride and methylmercury on plasma cortisol, T3, T4, glucose, and liver glycogen in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 1996, 34, 221–235. [Google Scholar] [CrossRef]
- Jesus, T.B.D.; Almeida, P.G.A.D.; Vergílio, C.D.S.; Machado, A.L.D.S.; Carvalho, C.E.V.D. Acute intraperitoneal mercury chloride contamination and distribution in liver, muscle and gill of a neotropical fish Hoplias malabaricus (BLOCK, 1794). Braz. Arch. Biol. Technol. 2011, 54, 379–386. [Google Scholar] [CrossRef]
- Roy George, K.; Malini, N.A.; Sandhya Rani, G.O. Biochemical changes in liver and muscle of the cichlid, Oreochromis mossambicus (Peters, 1852) exposed to sub-lethal concentration of mercuric chloride. Indian J. Fish. 2012, 59, 147–152. [Google Scholar]
- Karlsen, O.A.; Sheehan, D.; Goksøyr, A. Alterations in the Atlantic cod (Gadus morhua) hepatic thiol-proteome after methylmercury exposure. J. Toxicol. Environ. Heal. A 2014, 77, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Yadetie, F.; Karlsen, O.A.; Lanzén, A.; Berg, K.; Olsvik, P.; Hogstrand, C.; Goksøyr, A. Global transcriptome analysis of Atlantic cod (Gadus morhua) liver after in vivo methylmercury exposure suggests effects on energy metabolism pathways. Aquat. Toxicol. 2013, 126, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Mela, M.; Randi, M.A.F.; Ventura, D.F.; Carvalho, C.E.V.; Pelletier, E.; de Oliveira Ribeiro, C.A. Effects of dietary methylmercury on liver and kidney histology in the neotropical fish Hoplias malabaricus. Ecotoxicol. Environ. Saf. 2007, 68, 426–435. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Ribeiro, C.A.; Belger, L.; Pelletier, E.; Rouleau, C. Histopathological evidence of inorganic mercury and methyl mercury toxicity in the arctic charr (Salvelinus alpinus). Environ. Res. 2002, 90, 217–225. [Google Scholar] [CrossRef]
- Régine, M.B.; Gilles, D.; Yannick, D.; Alain, B. Mercury distribution in fish organs and food regimes: Significant relationships from twelve species collected in French Guiana (Amazonian basin). Sci. Total Environ. 2006, 368, 262–270. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Ribeiro, C.O.; Pelletier, E.; Pfeiffer, W.C.; Rouleau, C. Comparative uptake, bioaccumulation, and gill damages of inorganic mercury in tropical and nordic freshwater fish. Environ. Res. 2000, 83, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dörra, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Korbas, M.; Blechinger, S.R.; Krone, P.H.; Pickering, I.J.; George, G.N. Localizing organomercury uptake and accumulation in zebrafish larvae at the tissue and cellular level. Proc. Natl. Acad. Sci. USA 2008, 105, 12108–12112. [Google Scholar] [CrossRef] [PubMed]
- Korbas, M.; Krone, P.H.; Pickering, I.J.; George, G.N. Dynamic accumulation and redistribution of methylmercury in the lens of developing zebrafish embryos and larvae. J. Biol. Inorg. Chem. 2010, 15, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Korbas, M.; MacDonald, T.C.; Pickering, I.J.; George, G.N.; Krone, P.H. Chemical form matters: Differential accumulation of mercury following inorganic and organic mercury exposures in zebrafish larvae. ACS Chem. Biol. 2011, 7, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, E.; Talarico, E.; Corapi, B.; Perrotta, I.; Tripepi, S. Effects of a sublethal concentration of sodium lauryl sulphate on the morphology and Na+/K+ ATPase activity in the gill of the ornate wrasse (Thalassoma pavo). Ecotoxicol. Environ. Saf. 2008, 71, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, E.; Mauceri, A.; Maisano, M.; Bernabò, I.; Giannetto, A.; de Domenico, E.; Corapi, B.; Tripepi, S.; Fasulo, S. Ultrastructural and immunohistochemical investigation on the gills of the teleost, Thalassoma pavo L., exposed to cadmium. Acta Histochem. 2011, 113, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Ivanković, D.; Pavičić, J.; Erk, M.; Filipović-Marijić, V.; Raspor, B. Evaluation of the Mytilus galloprovincialis Lam. Digestive gland metallothionein as a biomarker in a long-term field study: Seasonal and spatial variability. Mar. Pollut. Bull. 2005, 50, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Sinaie, M.; Bastami, K.D.; Ghorbanpour, M.; Najafzadeh, H.; Shekari, M.; Haghparast, S. Metallothionein biosynthesis as a detoxification mechanism in mercury exposure in fish, spotted scat (Scatophagus argus). Fish Physiol. Biochem. 2010, 36, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Hamza-Chaffai, A.; Amiard, J.C.; Pellerin, J.; Joux, L.; Berthet, B. The potential use of metallothionein in the clam Ruditapes decussatus as a biomarker of in situ metal exposure. Comp. Biochem. Phys. C 2000, 127, 185–197. [Google Scholar] [CrossRef]
- De Boeck, G.; Ngo, T.T.H.; van Campenhout, K.; Blust, R. Differential metallothionein induction patterns in three freshwater fish during sublethal copper exposure. Aquat. Toxicol. 2003, 65, 413–424. [Google Scholar] [CrossRef]
- Bebianno, M.J.; Santos, C.; Canário, J.; Gouveia, N.; Sena-Carvalho, D.; Vale, C. Hg and metallothionein-like proteins in the black scabbardfish Aphanopus carbo. Food Chem. Toxicol. 2007, 45, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Braunbeck, T.; Storch, V.; Bresch, H. Species-specific reaction of liver ultrastructure in zebrafish (Brachydanio rerio) and trout (Salmo gairdneri) after prolonged exposure to 4-chloroaniline. Arch. Environ. Contam. Toxicol. 1990, 19, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Lin, J.; Yang, P.; Chen, Q.; Chu, X.; Gao, C.; Hu, J. Fine structure, enzyme histochemistry, and immunohistochemistry of liver in zebrafish. Anat. Rec. 2012, 295, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Paris-Palacios, S.; Biagianti-Risbourg, S.; Vernet, G. Biochemical and (ultra) structural hepatic perturbations of Brachydanio rerio (Teleostei, Cyprinidae) exposed to two sublethal concentrations of copper sulfate. Aquat. Toxicol. 2000, 50, 109–124. [Google Scholar] [CrossRef]
- Gogbold, D.L. Mercury in forest ecosystem: Risk and research needs. In Mercury Pollution: Integration and synthesis; Watras, C.J., Huckabee, J.W., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 295–304. [Google Scholar]
- Bais, U.E.; Lokhande, M.V. Effect of cadmium chloride on histopathological changes in the freshwater fish Ophiocephalus striatus (Channa). Int. J. Zool. Res. 2012, 8, 23–32. [Google Scholar] [CrossRef]
- Khidr, B.M.; Mekkawy, I.A.; Harabawy, A.S.; Ohaida, A.S. Effect of lead nitrate on the liver of the cichlid fish (Oreochromis niloticus): A light microscope study. Pak. J. Biol. Sci. 2012, 15, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Drevnick, P.E.; Roberts, A.P.; Otter, R.R.; Hammerschmidt, C.R.; Klaper, R.; Oris, J.T. Mercury toxicity in livers of northern pike (Esox lucius) from Isle Royale, USA. Comp. Biochem. Phys. C 2008, 147, 331–338. [Google Scholar] [CrossRef] [PubMed]
- De Lisle, R.C.; Sarras, M.P.; Hidalgo, J.; Andrews, G.K. Metallothionein is a component of exocrine pancreas secretion: Implications for zinc homeostasis. Am. J. Physiol Cell Physiol. 1996, 271, 1103–1110. [Google Scholar]
- Pearce, L.L.; Wasserloos, K.; Croix, C.M.S.; Gandley, R.; Levitan, E.S.; Pitt, B.R. Metallothionein, nitric oxide and zinc homeostasis in vascular endothelial cells. J. Nutr. 2000, 130, 1467S–1470S. [Google Scholar] [PubMed]
- Ecker, D.J.; Butt, T.R.; Sternberg, E.J.; Neeper, M.P.; Debouck, C.; Gorman, J.A.; Crooke, S.T. Yeast metallothionein function in metal ion detoxification. J. Biol. Chem. 1986, 261, 16895–16900. [Google Scholar] [PubMed]
- Huang, P.C.; Morris, S.; Dinman, J.; Pine, R.; Smith, B. Role of metallothionein in detoxification and tolerance to transition metals. Exp. Suppl. 1987, 52, 439–446. [Google Scholar] [CrossRef]
- Li, X.; Chen, H.; Epstein, P.N. Metallothionein protects islets from hypoxia and extends islet graft survival by scavenging most kinds of reactive oxygen species. J. Biol. Chem. 2004, 279, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.S.; Patel, K.M.; Roesijadi, G. Oyster metallothionein as an oxyradical scavenger: Implications for hemocyte defense responses. Dev. Comp. Immunol. 1999, 23, 443–449. [Google Scholar] [CrossRef]
- Morris, C.A.; Stürzenbaum, S.; Nicolaus, B.; Morgan, A.J.; Harwood, J.L.; Kille, P. Identification and characterisation of metallothioneins from environmental indicator species as potential biomonitors. In Metallothionein IV; Klaassen, C.D., Ed.; Birkhäuser Vargel: Basel, Switzerland, 1999; pp. 621–627. [Google Scholar]
- Hamza-Chaffai, A.; Amiard, J.C.; Cosson, R.P. Relationship between metallothioneins and metals in a natural population of the clam Ruditapes decussatus from Sfax coast: A non-linear model using Box-Cox transformation. Comp. Biochem. Phys. C 1999, 123, 153–163. [Google Scholar] [CrossRef]
- Cosson, R.P.; Amiard, J.C. Use of metallothionein as biomarkers of exposure to metals. In Use of Biomarkers for Environmental Quality Assessment; Lagadic, L., Caquet, T., Amiard, J.C., Ramade, F., Eds.; Science Publishers, Inc.: Enfield, NH, USA, 2000; pp. 79–111. [Google Scholar]
- Geffard, A.; Amiard-Triquet, C.; Amiard, J.C. Do seasonal changes affect metallothionein induction by metals in mussels, Mytilus edulis? Ecotoxicol. Environ. Saf. 2005, 61, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Knapen, D.; Reynders, H.; Bervoets, L.; Verheyen, E.; Blust, R. Metallothionein gene and protein expression as a biomarker for metal pollution in natural gudgeon populations. Aquat. Toxicol. 2007, 82, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, L.; Bai, Q.; Zhu, X.; Zhang, J.; Wei, Q.; Li, D.; Gao, C.; Li, J.; Zhang, Z.; et al. Heavy metal-induced metallothionein expression is regulated by specific protein phosphatase 2A complexes. J. Biol. Chem. 2014, 289, 22413–22426. [Google Scholar] [CrossRef] [PubMed]
- Thophon, S.; Pokethitiyook, P.; Chalermwat, K.; Upatham, E.S.; Sahaphong, S. Ultrastructural alterations in the liver and kidney of white sea bass, Lates calcarifer, in acute and subchronic cadmium exposure. Environ. Toxicol. 1993, 19, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kondoh, M. Recent studies on metallothionein: Protection against toxicity of heavy metals and oxygen free radicals. Tohoku J. Exp. Med. 2002, 196, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Woo, E.S.; Lazo, J.S. Nucleocytoplasmic functionality of metallothionein. Cancer Res. 1997, 57, 4236–4241. [Google Scholar] [PubMed]
- Ivanina, A.V.; Cherkasov, A.S.; Sokolova, I.M. Effects of cadmium on cellular protein and glutathione synthesis and expression of stress proteins in eastern oysters, Crassostrea virginica Gmelin. J. Exp. Biol. 2008, 211, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Vutukuru, S.S.; Basani, K. Acute effects of mercuric chloride on glycogen and protein content of Zebra fish, Danio rerio. J. Environ. Biol. 2013, 34, 277. [Google Scholar] [PubMed]
- Coons, A.H.; Leduc, E.H.; Connolly, J.M. Studies on antibody. I. A method for thehistochemical demonstration of specific antibody and its application to a studyof the hyperimmune rabbit. J. Exp. Med. 1955, 102, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Ebbesson, L.O.E.; Nilsen, T.O.; Helvik, J.V.; Tronci, V.; Stefansson, S.O. Corticotropin-releasing factor neurogenesis during midlife development in salmon: Genetic, environmental and thyroid hormone regulation. J. Neuroendocrinol. 2011, 23, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; John, J.A.C.; Lin, C.H.; Lin, H.F.; Wu, S.C.; Lin, C.H.; Chang, C.Y. Expression of metallothionein gene during embryonic and early larval development in zebrafish. Aquat. Toxicol. 2004, 69, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macirella, R.; Guardia, A.; Pellegrino, D.; Bernabò, I.; Tronci, V.; Ebbesson, L.O.E.; Sesti, S.; Tripepi, S.; Brunelli, E. Effects of Two Sublethal Concentrations of Mercury Chloride on the Morphology and Metallothionein Activity in the Liver of Zebrafish (Danio rerio). Int. J. Mol. Sci. 2016, 17, 361. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030361
Macirella R, Guardia A, Pellegrino D, Bernabò I, Tronci V, Ebbesson LOE, Sesti S, Tripepi S, Brunelli E. Effects of Two Sublethal Concentrations of Mercury Chloride on the Morphology and Metallothionein Activity in the Liver of Zebrafish (Danio rerio). International Journal of Molecular Sciences. 2016; 17(3):361. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030361
Chicago/Turabian StyleMacirella, Rachele, Antonello Guardia, Daniela Pellegrino, Ilaria Bernabò, Valentina Tronci, Lars O. E. Ebbesson, Settimio Sesti, Sandro Tripepi, and Elvira Brunelli. 2016. "Effects of Two Sublethal Concentrations of Mercury Chloride on the Morphology and Metallothionein Activity in the Liver of Zebrafish (Danio rerio)" International Journal of Molecular Sciences 17, no. 3: 361. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030361