
Chemical Elicitor-Induced Modulation of Antioxidant Metabolism and Enhancement of Secondary Metabolite Accumulation in Cell Suspension Cultures of Scrophularia kakudensis Franch




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
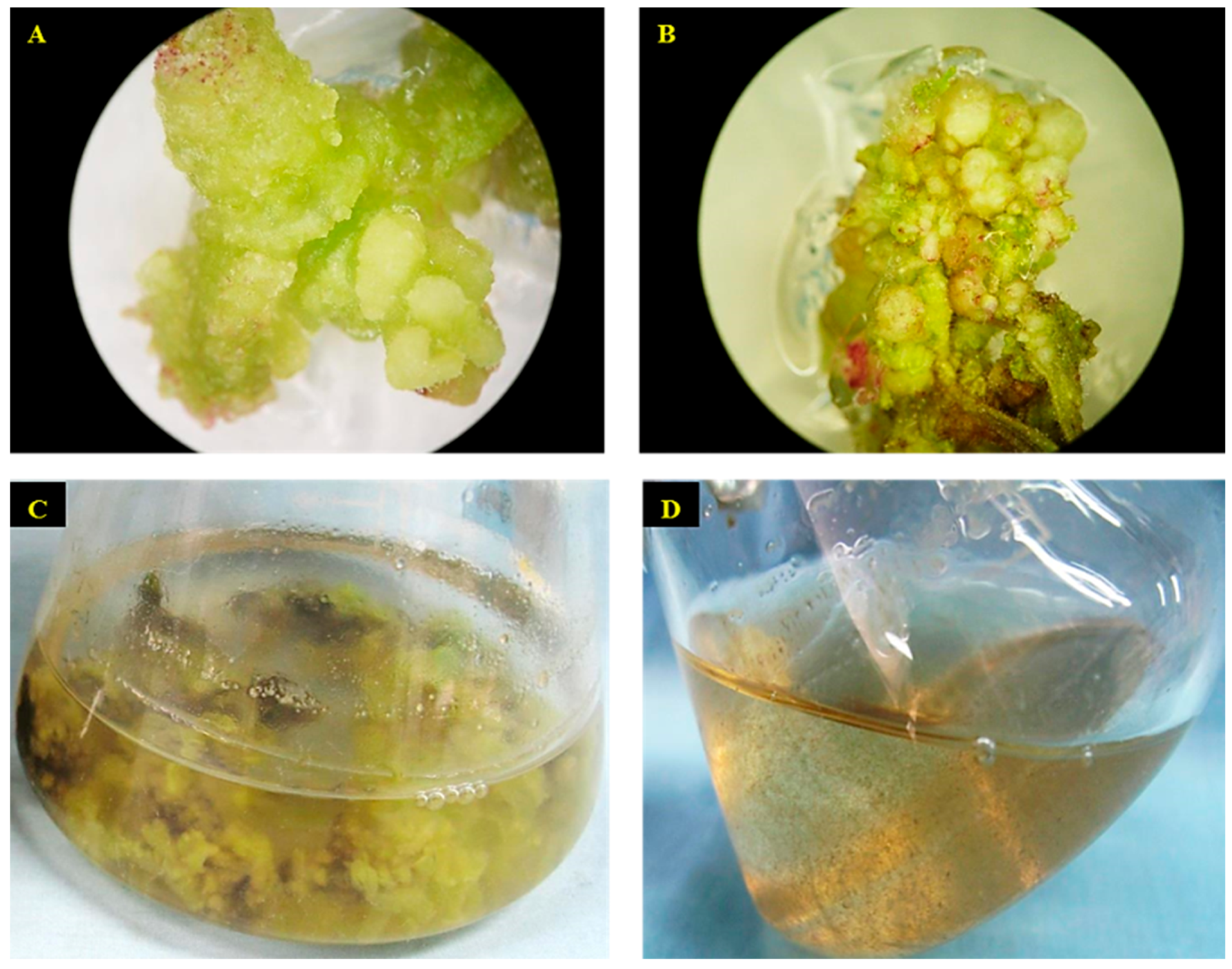
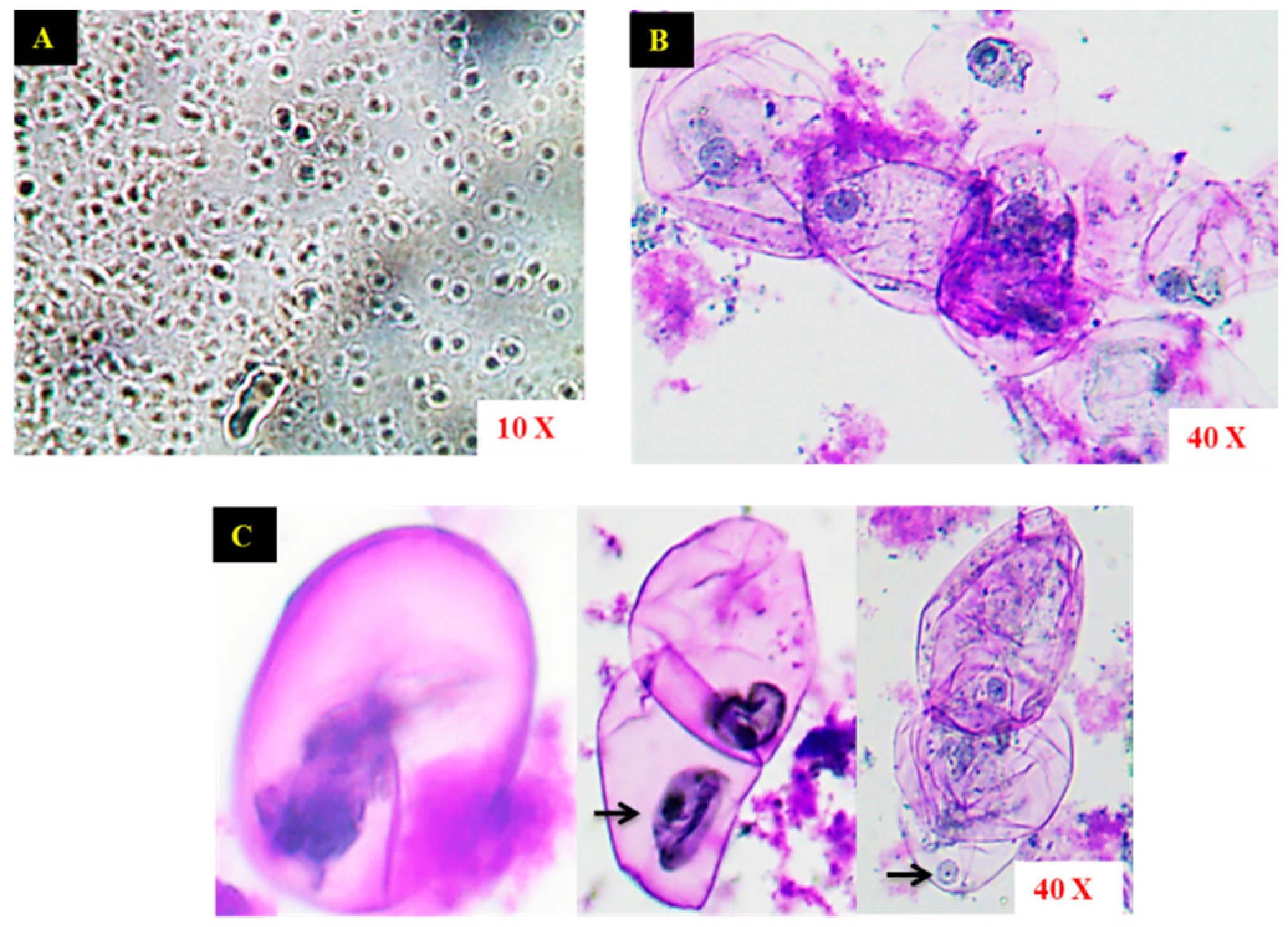
2.1. Establishment of Cell Suspension Culture and Morphological Observation of Cells
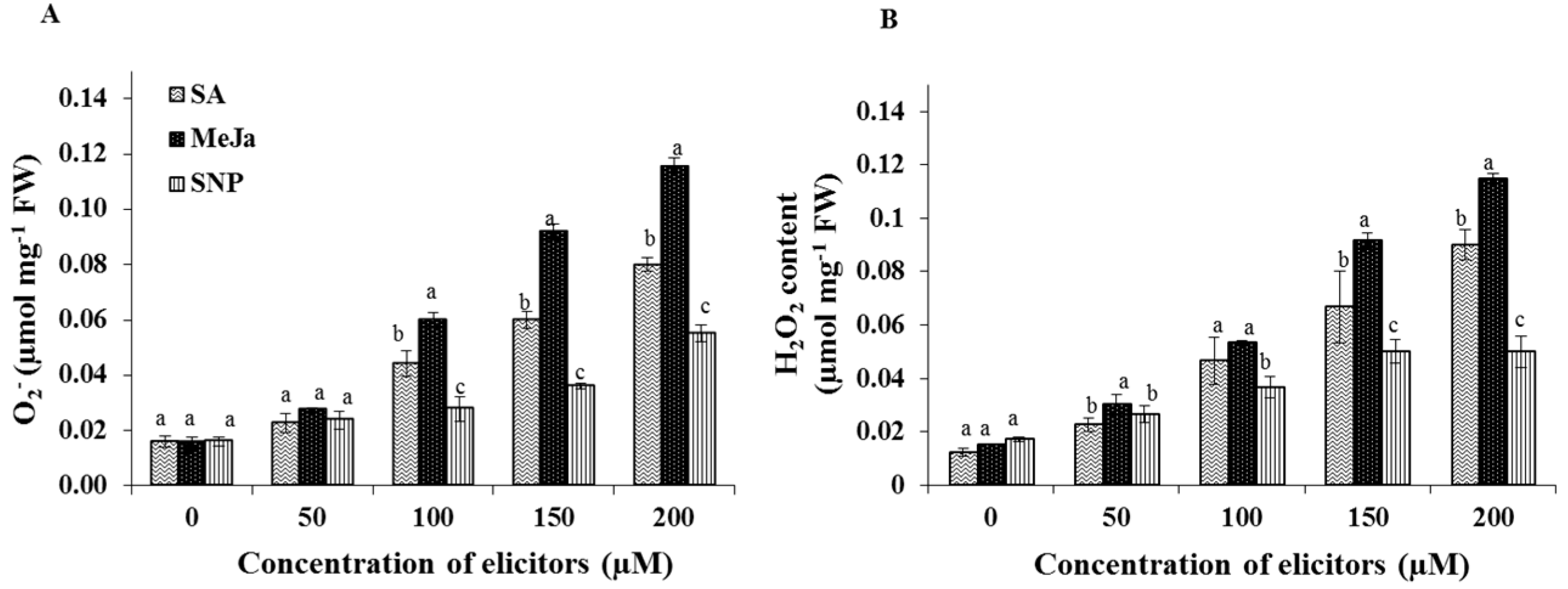
2.2. Effect of Chemical Elicitors on the Contents of Endogenous Free Radicals
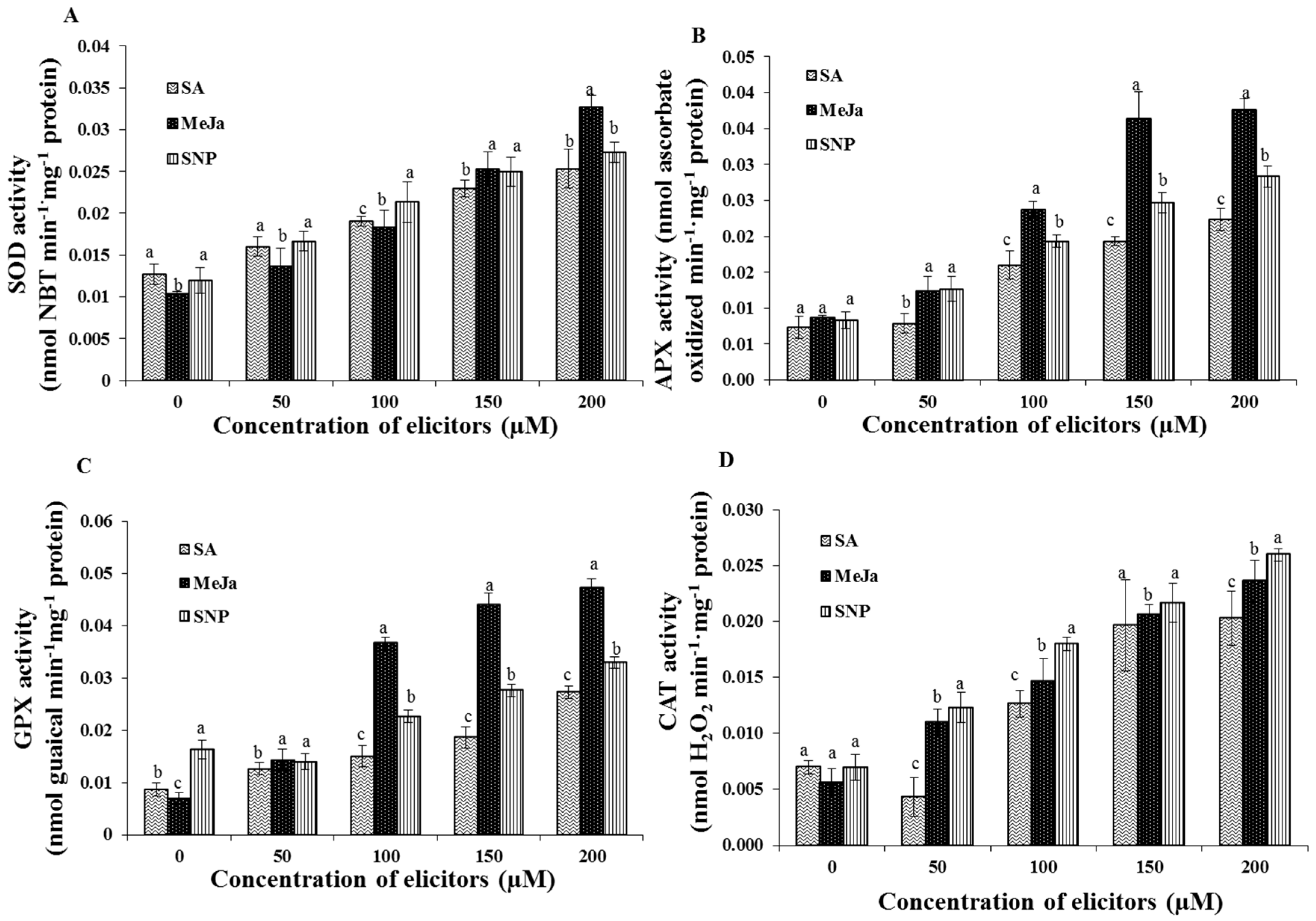
2.3. Effect of Chemical Elicitors on the Antioxidant Enzyme Activities of the Cells
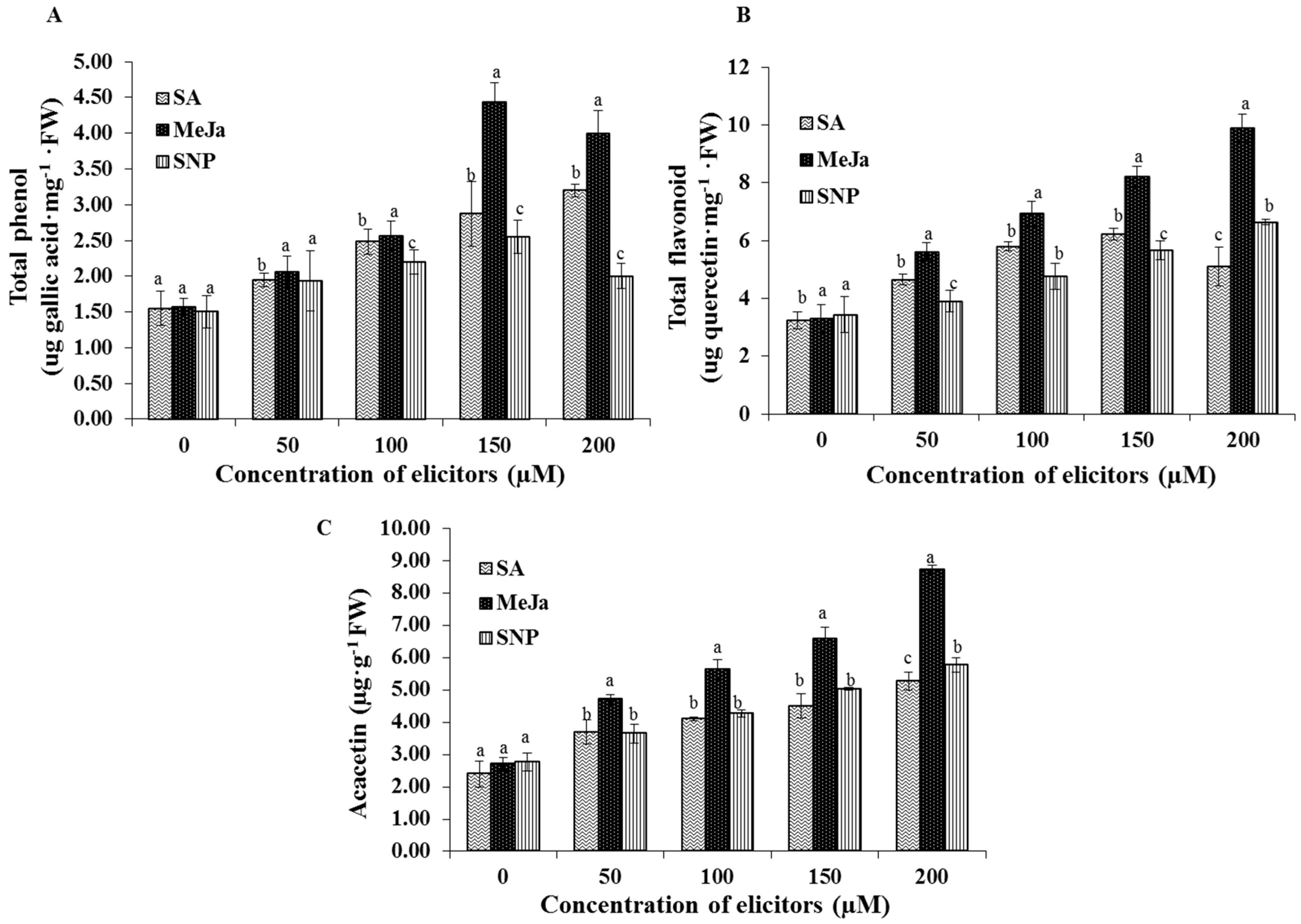
2.4. Effect of Chemical Elicitors on the Accumulation of Bioactive Compounds
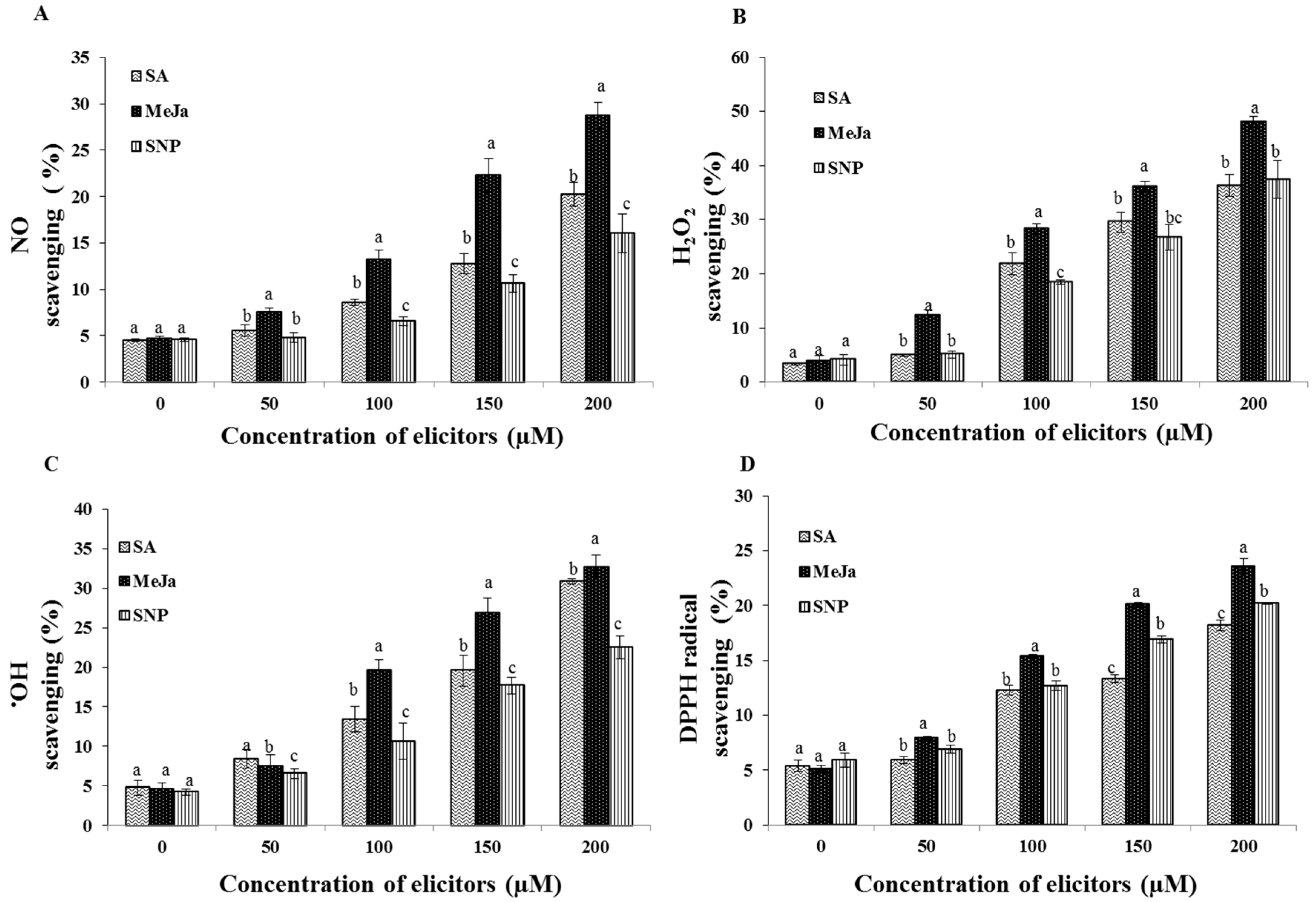
2.5. Effect of Chemical Elicitors on the Free Radical Scavenging Potential of the Cell Extracts
3. Discussion
4. Experimental Section
4.1. Plant Materials and Culture Conditions
4.2. Establishment of Cell Suspension and Elicitor Treatments
4.3. Microscopic Observation of Cells
4.4. Estimation of Superoxide (O2−) and Hydrogen Peroxide (H2O2)
4.5. Estimation of Antioxidant Enzyme Activities
4.6. Extract Preparation for Phytochemical Analysis
4.7. Estimation of Bioactive Compounds
4.7.1. Estimation of Total Phenols and Flavonoids
4.7.2. Quantification of Acacetin Using High Performance Liquid Chromatography (HPLC)
4.8. Assessment of Free Radicals Scavenging Potential of Cell Extracts
4.8.1. Superoxide (O2−) Radical Scavenging Assay
4.8.2. Nitric Oxide (NO) Radical Scavenging Assay
4.8.3. Hydrogen Peroxide (H2O2) Radical Scavenging Assay
4.8.4. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ahmad, S.; Garg, M.; Tamboli, E.T.; Abdin, M.Z.; Ansari, S.H. In vitro production of alkaloids: Factors, approaches, challenges and prospects. Pharmacogn. Rev. 2013, 7, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Rischer, H.; Hakkinen, S.T.; Ritala, A.; Seppanen-Laakso, T.; Miralpeix, B.; Capell, T.; Christou, P.; Oksman-Caldentey, K. Plant cells as pharmaceutical factories. Curr. Pharm. Des. 2013, 19, 5640–5660. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Ming, Q.L.; Lin, B.; Rahman, K.; Zheng, C.J.; Han, T.; Qin, L.P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.S.; Fareed, S.; Ansari, S.; Rahman, M.A.; Ahmad, I.Z.; Saeed, M. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Sivanandhan, G.; Dev, G.K.; Jeyaraj, M.; Rajesh, M.; Arjunan, A.; Muthuselvam, M.; Manickavasagam, M.; Selvaraj, N.; Ganapathi, A. Increased production of withanolide A, withanone, and withaferin A in hairy root cultures of Withania somnifera (L.) Dunal elicited with methyl jasmonate and salicylic acid. Plant Cell Tissue Organ Cult. 2013, 114, 121–129. [Google Scholar] [CrossRef]
- Wu, C.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Nitric oxide elicitation induces the accumulation of secondary metabolites and antioxidant defense in adventitious roots of Echinacea purpurea. J. Plant Biol. 2007, 6, 636–643. [Google Scholar] [CrossRef]
- Boonsnongcheep, P.; Korsangruang, S.; Soonthornchareonnon, N.; Chintapakorn, Y.; Saralamp, P.; Prathanturarug, S. Growth and isoflavonoid accumulation of Pueraria candollei var. candollei and P. candollei var. mirifica cell suspension cultures. Plant Cell Tissue Organ Cult. 2010, 101, 119–126. [Google Scholar] [CrossRef]
- Frankfater, C.R.; Dowd, M.K.; Triplett, B.A. Effect of elicitors on the production of gossypol and methylated gossypol in cotton hairy roots. Plant Cell Tissue Organ Cult. 2009, 98, 341–349. [Google Scholar] [CrossRef]
- Bonfill, M.; Mangas, S.; Moyano, E.; Cusido, R.M.; Palazoon, J. Production of centellosides and phytosterols in cell suspension cultures of Centella asiatica. Plant Cell Tissue Organ Cult. 2011, 104, 61–67. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 201, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ahn, M.; Lee, S. Isolation and identification of phytochemical constituents from Scrophularia. takesimensis. J. Med. Plants Res. 2012, 6, 3923–3930. [Google Scholar]
- Yamamoto, A.; Miyase, T.; Ueno, A.; Maeda, T. Scrophulasaponins II-IB, new saikosaponin homologs from Scrophularia. kakudensis FRANCH. Chem. Pharm. Bull. 1993, 41, 1780–1783. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, A.; Soundararajan, P.; Park, Y.G.; Jeong, B.R. In vitro propagation, phytochemical analysis, and evaluation of free radical scavenging property of Scrophularia kakudensis Franch tissue extracts. Biomed. Res. Int. 2015, 2015, 480564. [Google Scholar] [CrossRef] [PubMed]
- Sesterhenn, K.; Distl, M.; Wink, M. Occurrence of iridoid glycosides in in vitro cultures and intact plants of Scrophularia nodosa L. Plant Cell Rep. 2007, 26, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Khanpour-Ardestani, N.; Sharifi, M.; Behmanesh, M. Establishment of callus and cell suspension culture of Scrophularia striata Boiss: An in vitro approach for acteoside production. Cytotechnology 2015, 67, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Oksman-Caldentey, K.M.; Inzé, D. Plant cell factories in the post-genomic era: New ways to produce designer secondary metabolites. Trends Plant Sci. 2004, 9, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [PubMed]
- Subhashini, P.; Raja, S.; Thangaradjou, T. Establishment of cell suspension culture protocol for a seagrass (Halodule pinifolia): Growth kinetics and histomorphological characterization. Aquat. Bot. 2014, 117, 33–40. [Google Scholar] [CrossRef]
- Nagella, P.; Murthy, H.N. Establishment of cell suspension cultures of Withania somnifera for the production of with anolide A. Bioresour. Technol. 2010, 101, 6735–6739. [Google Scholar] [CrossRef] [PubMed]
- Lopes, S.O.; Moreno, P.R.; Henriques, A.T. Growth characteristics and chemical analysis of Psychotria carthagenensis cell suspension cultures. Enzyme Microb. Technol. 2000, 26, 259–264. [Google Scholar] [CrossRef]
- Sangduen, N.; Prasertsongskun, S. Regeneration and Application: From suspension cultured derived inflorescences of Vetiveria zizanioides (L.) Nash to selection of herbicide resistant cell. AU. J. Technol. 2009, 12, 135–148. [Google Scholar]
- Delledonne, M.; Xia, Y.; Dixon, R.A.; Lamb, C. Nitric oxide functions as a signal in plant disease resistance. Nature 1998, 394, 585–588. [Google Scholar] [PubMed]
- Lamb, C.; Dixon, R.A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Biol. 1997, 48, 251–275. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lawrence, C.D.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Kauss, H.; Jeblick, W.; Ziegler, J.; Krabler, W. Pretreatment of parsley (Petroselinum crispum L.) suspension cultures with methyl jasmonate enhances elicitation of activated oxygen species. Plant Physiol. 1994, 105, 89–94. [Google Scholar] [PubMed]
- Wang, S.Y.; Bowman, L.; Ding, M. Methyl jasmonate enhances antioxidant activity and flavonoid content in blackberries (Rubus sp.) and promotes antiproliferation of human cancer cells. Food Chem. 2008, 107, 1261–1269. [Google Scholar] [CrossRef]
- Zhang, L.; Da, X. Methyl jasmonate induces production of reactive oxygen species and alterations in mitochondrial dynamics that precede photosynthetic dysfunction and subsequent cell death. Plant Cell Physiol. 2008, 49, 1092–1111. [Google Scholar]
- Mur, L.A.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid elicitation induces ginsenosides accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Rep. 2006, 25, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.T.; Yi, T.H.; Kao, C.H. Hydrogen peroxide is involved in methyl jasmonate-induced senescence of rice leaves. Physiol. Plant. 2006, 127, 293–303. [Google Scholar] [CrossRef]
- Orozco-Cárdenas, M.L.; Narváez-Vásquez, J.; Ryan, C.A. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin, and methyl jasmonate. Plant Cell 2001, 13, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Joseph, C.; Hagege, D. Jasmonic acid elicitation of Hypericum perforatum L. cell suspensions and effects on the production of phenylpropanoids and naphtodianthrones. Plant Cell Tissue Organ Cult. 2007, 89, 1–13. [Google Scholar] [CrossRef]
- Creelman, R.A.; Tierney, M.L.; Mullet, J.E. Jasmonic acid/methyl jasmonate accumulate in wounded soybean hypocotyls and modulate wound gene expression. Proc. Natl. Acad. Sci. USA 1992, 89, 4938–4941. [Google Scholar] [CrossRef] [PubMed]
- Mirjalili, N.; Linden, J.C. Methyl jasmonate induced production of taxol in suspension cultures of Taxus cuspidata: Ethylene interaction and induction models. Biotechnol. Prog. 1996, 12, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Tamari, G.; Borochov, A.; Atzorn, R.; Weiss, D. Methyl jasmonate induces pigmentation and flavonoid gene expression in petunia corollas: A possible role in wound response. Physiol. Plant. 1995, 94, 45–50. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. LWT Food Sci. Technol. 2007, 40, 344–352. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: Oxford, UK, 2007; p. 3. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium of rapid growth and bioassay with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Manivannan, A.; Prabhakaran, S.; Laras, S.A.; Chung, H.K.; Muneer, S.; Jeong, B.R. Silicon-mediated enhancement of physiological and biochemical characteristics of Zinnia elegans “Dreamland Yellow” grown under salinity stress. Hortic. Environ. Biotechnol. 2015, 56, 721–731. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yang, W.J.; Liu, C.; Gu, Z.Y.; Zhang, X.Y.; Cheng, B.; Mao, Y.; Xue, G.P. Protective effects of acacetin isolated from Ziziphora clinopodioides Lam. (Xintahua) on neonatal rat cardiomyocytes. Chin. Med. 2014, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manivannan, A.; Soundararajan, P.; Park, Y.G.; Jeong, B.R. Chemical Elicitor-Induced Modulation of Antioxidant Metabolism and Enhancement of Secondary Metabolite Accumulation in Cell Suspension Cultures of Scrophularia kakudensis Franch. Int. J. Mol. Sci. 2016, 17, 399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030399
Manivannan A, Soundararajan P, Park YG, Jeong BR. Chemical Elicitor-Induced Modulation of Antioxidant Metabolism and Enhancement of Secondary Metabolite Accumulation in Cell Suspension Cultures of Scrophularia kakudensis Franch. International Journal of Molecular Sciences. 2016; 17(3):399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030399
Chicago/Turabian StyleManivannan, Abinaya, Prabhakaran Soundararajan, Yoo Gyeong Park, and Byoung Ryong Jeong. 2016. "Chemical Elicitor-Induced Modulation of Antioxidant Metabolism and Enhancement of Secondary Metabolite Accumulation in Cell Suspension Cultures of Scrophularia kakudensis Franch" International Journal of Molecular Sciences 17, no. 3: 399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030399