
Koumine Attenuates Lipopolysaccaride-Stimulated Inflammation in RAW264.7 Macrophages, Coincidentally Associated with Inhibition of NF-κB, ERK and p38 Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
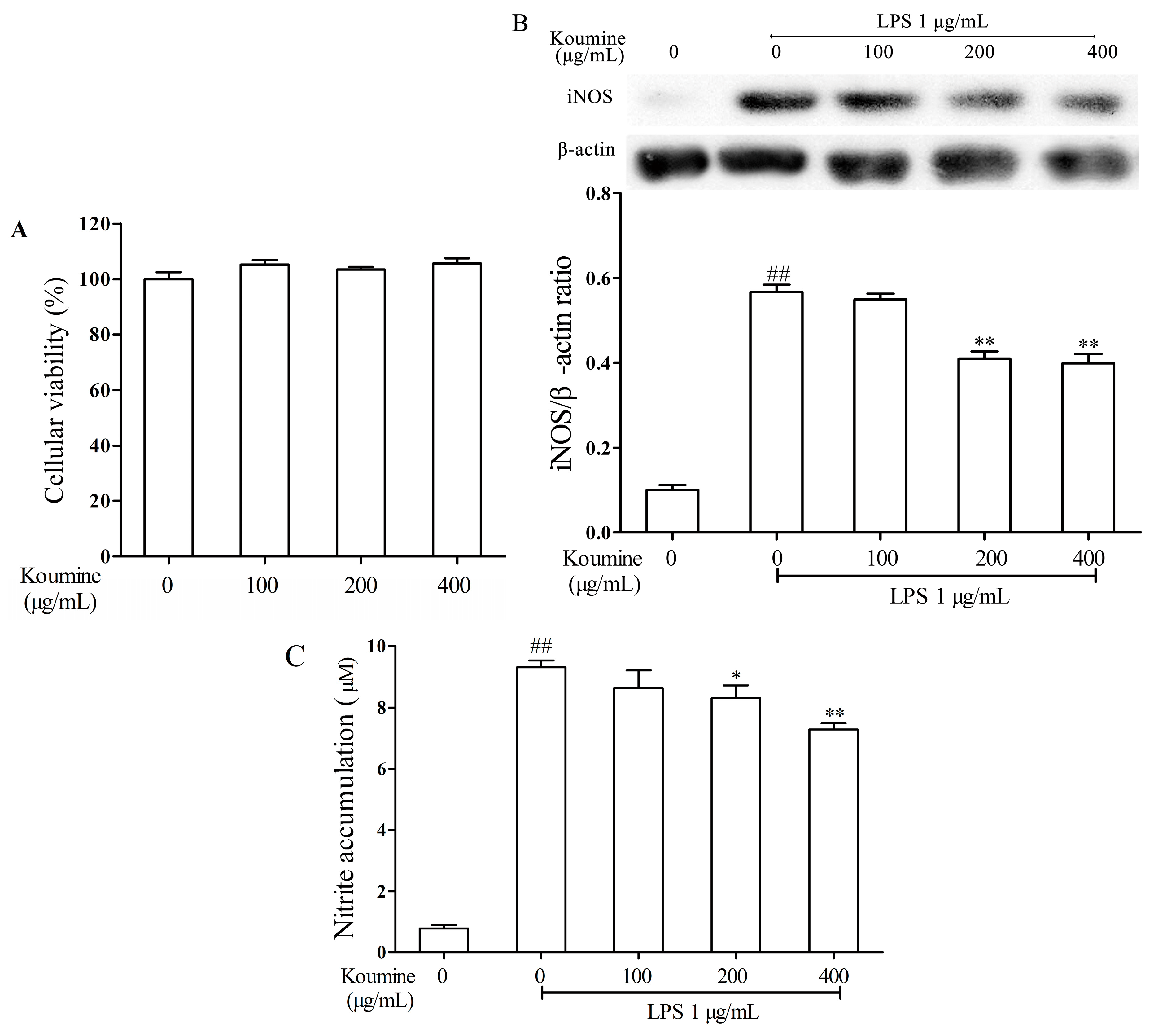
2.1. Koumine Suppresses the Levels of NO and iNOS in LPS-Stimulated RAW264.7 Cells
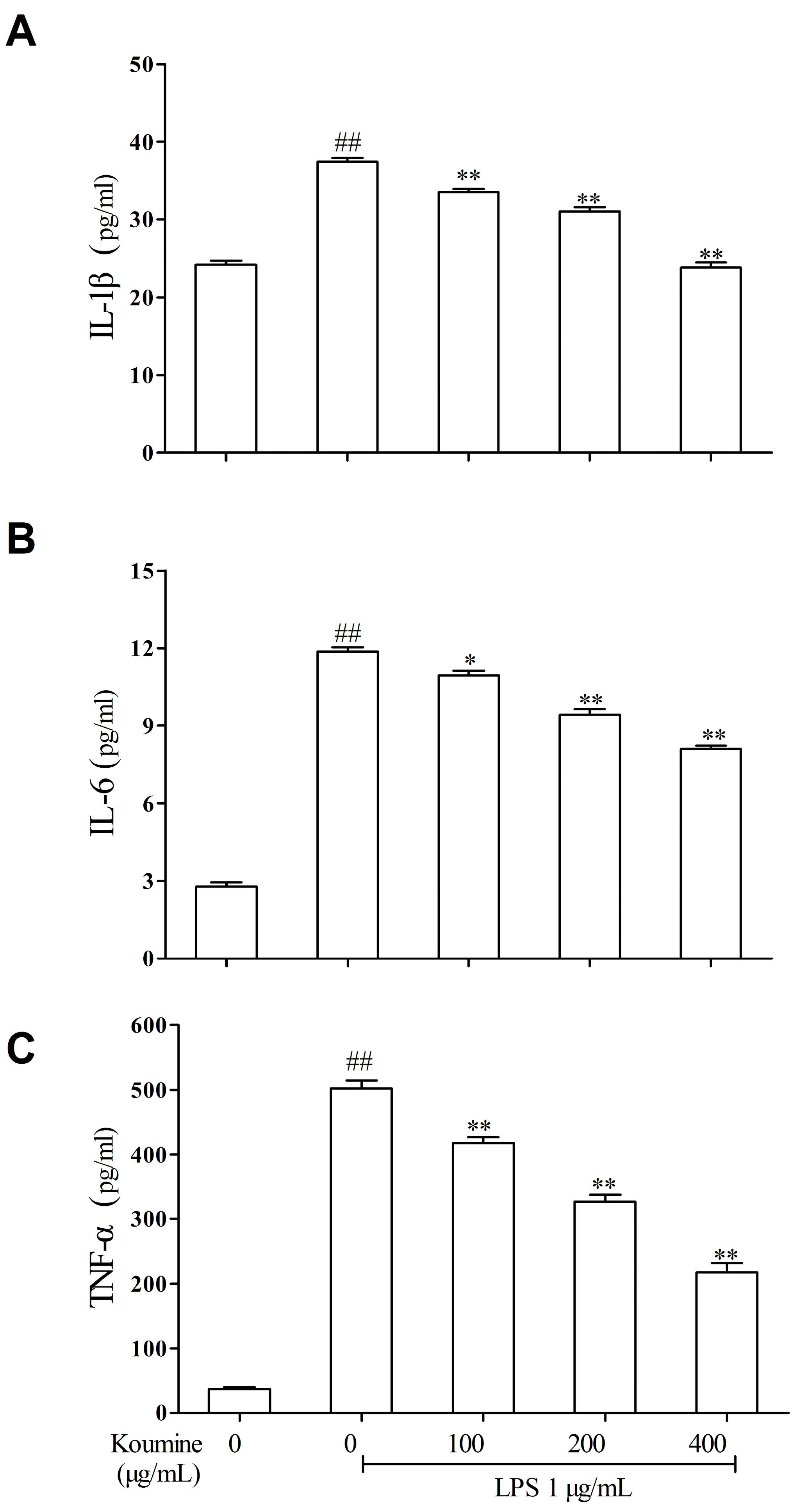
2.2. Koumine Attenuates the Production of Pro-Inflammatory Cytokines in LPS-Treated RAW264.7 Macrophages
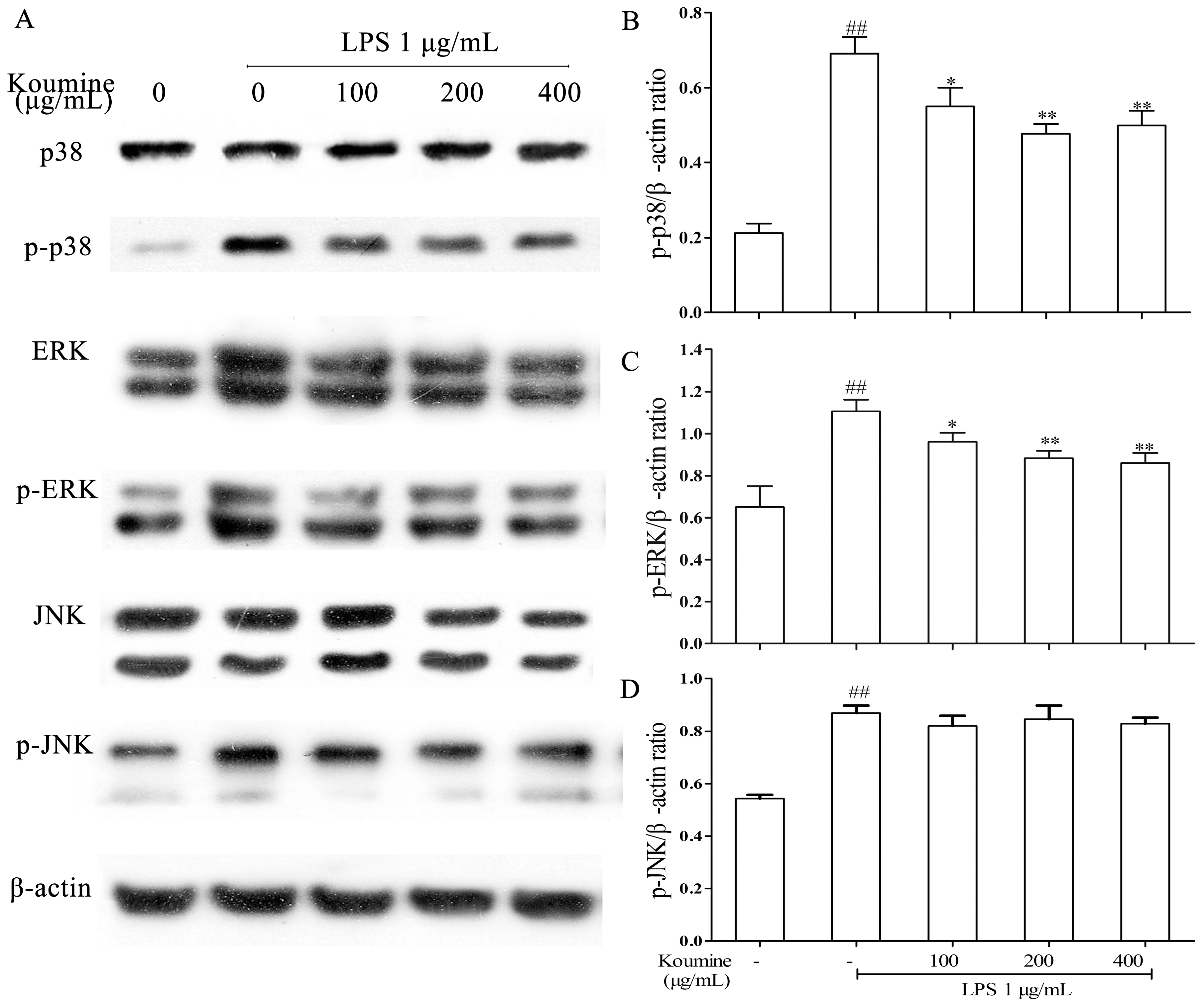
2.3. Koumine Inhibits p38 and ERK Phosphorylation in LPS-Stimulated RAW264.7 Macrophages
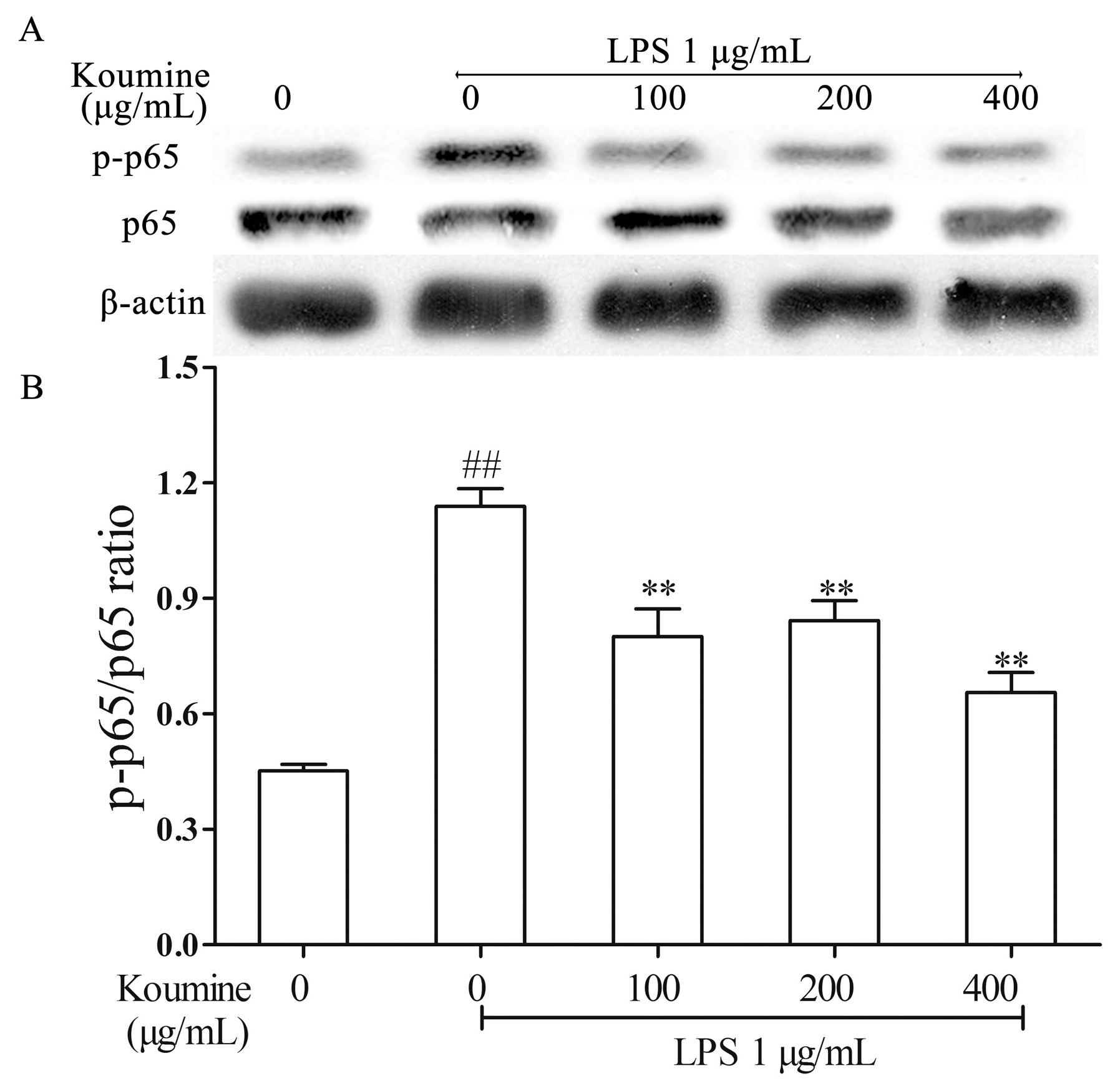
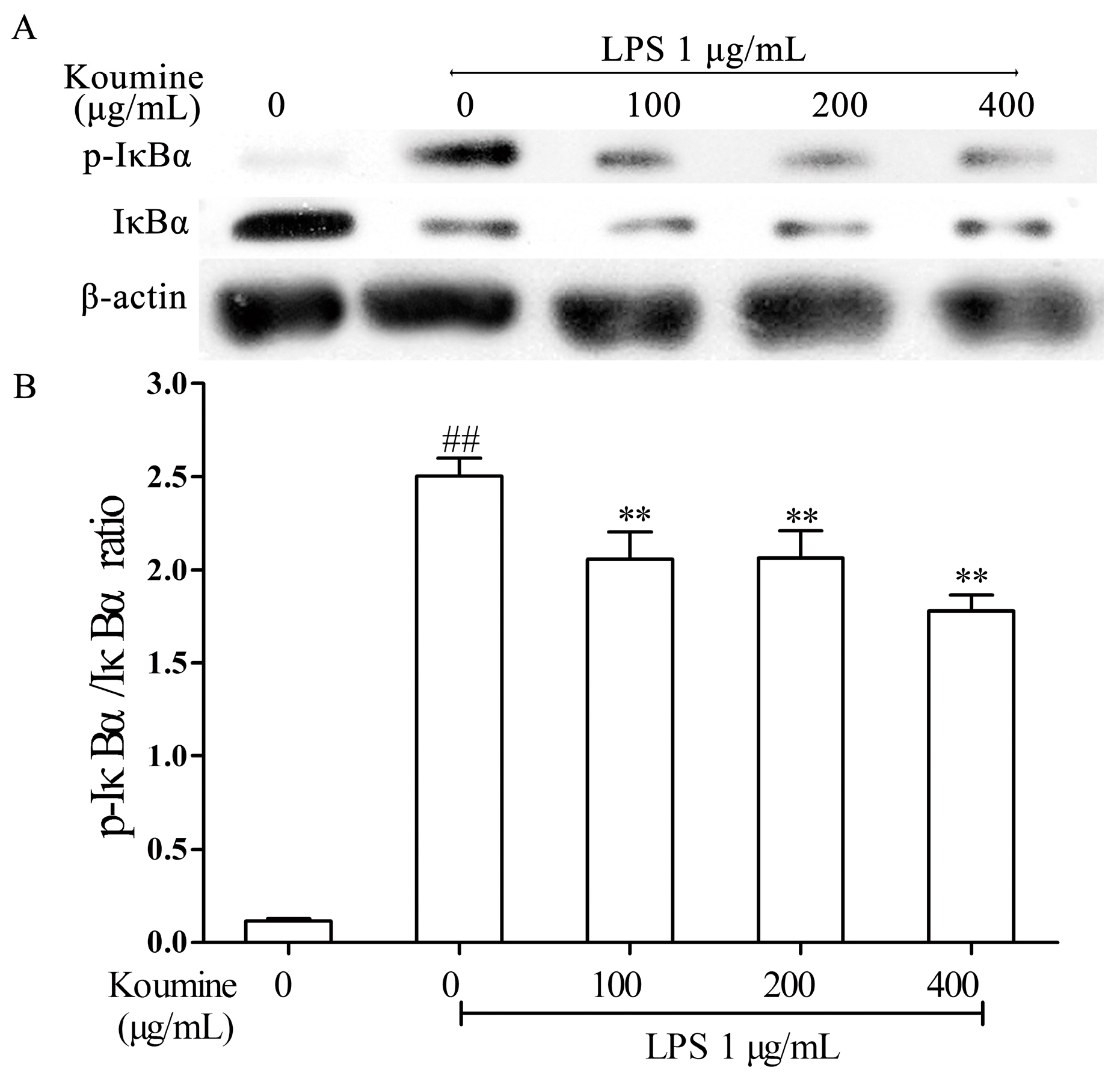
2.4. Koumine Hampered IκBα and p65 Phosphorylation in LPS-Stimulated RAW264.7 Macrophages
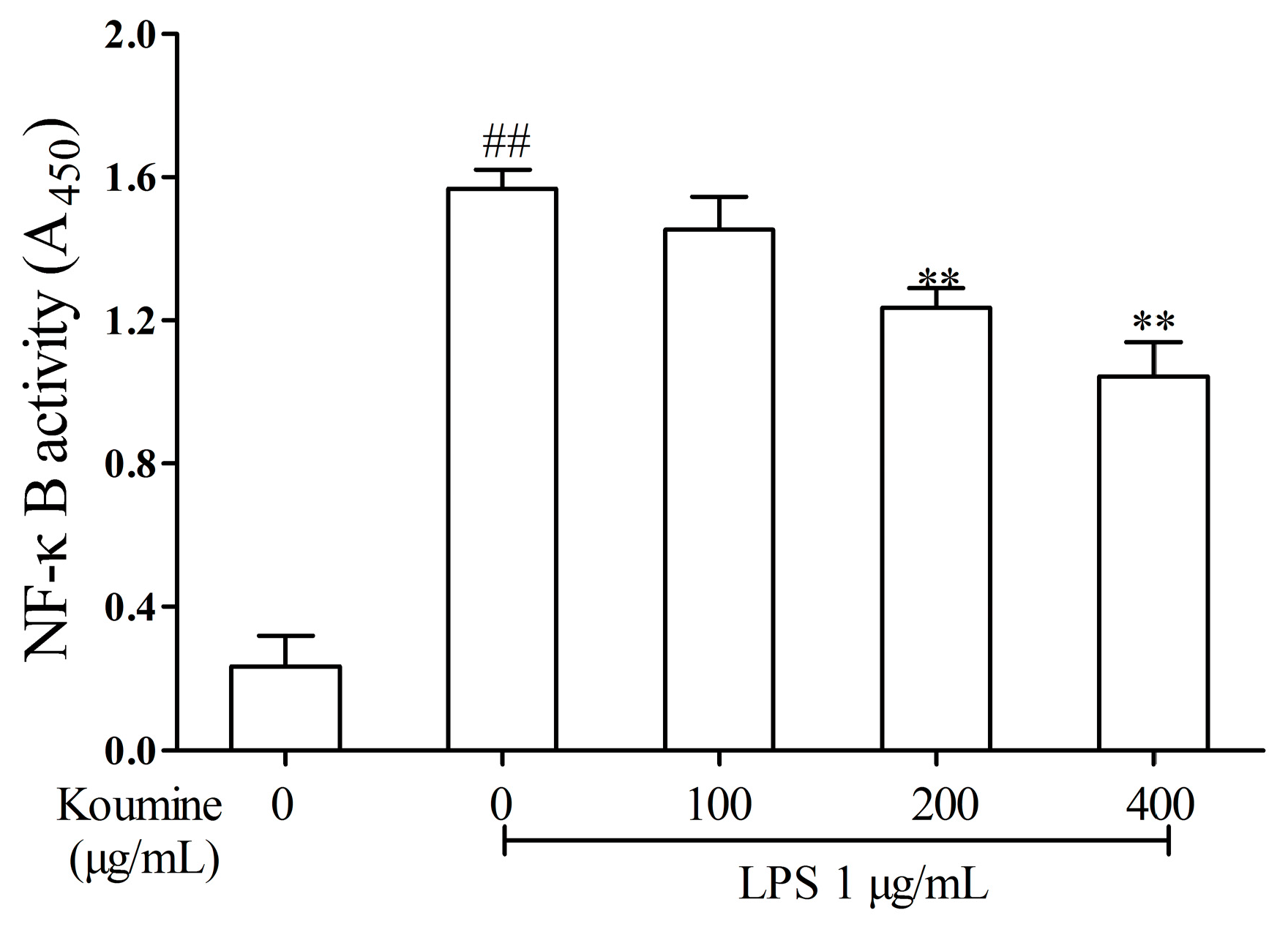
2.5. Effect of Koumine on DNA-Binding Activity of NF-κB in LPS-Treated RAW264.7 Cells
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Culture
4.3. Assessment of Cell Cytotoxicity
4.4. Measurement of Nitric Oxide and Pro-Inflammatory Cytokines
4.5. Western Blot Analysis
4.6. Nuclear Protein Extracts
4.7. NF-κB DNA-Binding Assay
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contribution
Conflicts of Interest
References
- Malyshev, I.Y.; Shnyra, A. Controlled modulation of inflammatory, stress and apoptotic responses in macrophages. Curr. Drug Targets Immune Endocr. Metab. Disord. 2003, 3, 1–22. [Google Scholar] [CrossRef]
- Bredt, D.S. Endogenous nitric oxide synthesis: Biological functions and pathophysiology. Free Radic. Res. 1999, 31, 577–596. [Google Scholar] [CrossRef] [PubMed]
- Tennyson, A.G.; Lippard, S.J. Generation, translocation, and action of nitric oxide in living systems. Chem. Biol. 2011, 18, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.; Clark, J.M.; Buzas, E.I.; Gorman, C.L.; Cope, A.P. Nitric oxide, chronic inflammation and autoimmunity. Immunol. Lett. 2007, 111, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Van Zonneveld, A.; de Boer, H.; van Der Veer, E.; Rabelink, T. Inflammation, vascular injury and repair in rheumatoid arthritis. Ann. Rheum. Dis. 2010, 69, i57–i60. [Google Scholar] [CrossRef] [PubMed]
- Oates, J.C.; Shaftman, S.R.; Self, S.E.; Gilkeson, G.S. Association of serum nitrate and nitrite levels with longitudinal assessments of disease activity and damage in systemic lupus erythematosus and lupus nephritis. Arthritis Rheum. 2008, 58, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Onur, O.; Akinci, A.S.; Akbiyik, F.; Unsal, I. Elevated levels of nitrate in rheumatoid arthritis. Rheumatol. Int. 2001, 20, 154–158. [Google Scholar] [PubMed]
- Ohashi, J.; Naka1, I.; Patarapotikul, J.; Hananantachai, H.; Looareesuwan, S.; Tokunaga, K. Significant association of longer forms of CCTTT Microsatellite repeat in the inducible nitric oxide synthase promoter with severe malaria in Thailand. J. Infect. Dis. 2002, 186, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Oates, J.C.; Levesque, M.C.; Hobbs, M.R.; Smith, E.G.; Molano, I.D.; Page, G.P.; Hill, B.S.; Weinberg, J.B.; Cooper, G.S.; Gilkeson, G.S. Nitric oxide synthase 2 promoter polymorphisms and systemic lupus erythematosus in african-americans. J. Rheumatol. 2003, 30, 60–67. [Google Scholar] [PubMed]
- Bouali, H.; Nietert, P.; Nowling, T.M.; Pandey, J.; Dooley, M.A.; Cooper, G.; Harley, J.; Kamen, D.L.; Oates, J.; Gilkeson, G. Association of G-463A myeloperoxidase gene polymorphism with renal disease in African Americans with systemic lupus erythematosus. J. Rheumatol. 2007, 34, 2028–2034. [Google Scholar] [PubMed]
- Connelly, L.; Jacobs, A.T.; Palacios-Callender, M.; Moncada, S.; Hobbs, A.J. Macrophage endothelial nitric-oxide synthase autoregulates cellular activation and pro-inflammatory protein expression. J. Biol. Chem. 2003, 278, 26480–26487. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.Y.; Huang, W.; Asagiri, M.; Spann, N.; Hoffmann, A.; Glass, C.; Ghosh, G. The transcriptional specificity of nf-κb dimers is coded within the κB DNA response elements. Cell Rep. 2012, 2, 824–839. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D. Introduction to NF-κB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed]
- Sizemore, N.; Lerner, N.; Dombrowski, N.; Sakurai, H.; Stark, G.R. Distinct roles of the iκ B kinase α and β subunits in liberating nuclear factor κB (NF-κB) from iκB and in phosphorylating the p65 subunit of NF-κB. J. Biol. Chem. 2002, 277, 3863–3869. [Google Scholar] [CrossRef] [PubMed]
- Nozell, S.; Laver, T.; Moseley, D.; Nowoslawski, L.; De Vos, M.; Atkinson, G.P.; Harrison, K.; Nabors, L.B.; Benveniste, E.N. The ING4 tumor suppressor attenuates NF-κB activity at the promoters of target genes. Mol. Cell. Biol. 2008, 28, 6632–6645. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Nowak, D.E.; Jamaluddin, M.; Wang, S.; Brasier, A.R. Identification of direct genomic targets downstream of the nuclear factor-κB transcription factor mediating tumor necrosis factor signaling. J. Biol. Chem. 2005, 280, 17435–17448. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Shen, J.; Liu, H.; Xu, Y.; Su, Y.-P.; Yang, J.; Yu, C.X. Gelsenicine from Gelsemium elegans attenuates neuropathic and inflammatory pain in mice. Biol. Pharm. Bull. 2011, 34, 1877–1880. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, H.H.; Yang, J.; Su, Y.P.; Lin, H.W.; Lin, L.Q.; Liao, W.J.; Yu, C.X. The active alkaloids of Gelsemium elegans benth. are potent anxiolytics. Psychopharmacology 2013, 225, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.R.; Qin, R.; Cai, J.; Chi, D.B. Antitumor activity of koumine in vitro and vivo. Pharmacol. Clin. Chin. Mater. Med. 2006, 22, 6–8. [Google Scholar]
- Tan, J.Q.; Qiu, C.Z.; Zheng, L.Z. Analgesic effect and no physical dependence of Gelsemium elegans Benth. Pharmacol. Clin. Chin. Mater. Med. 1988, 4, 24–28. [Google Scholar]
- Zhang, L.L.; Wang, Z.R.; Huang, C.Q.; Zhang, Z.Y.; Lin, J.M. Extraction and separation of koumine from Gelsemium alkaloids. J. First Mil. Med. Univ. 2004, 24, 1006–1008. [Google Scholar]
- Xu, Y.; Qiu, H.Q.; Liu, H.; Liu, M.; Huang, Z.Y.; Yang, J.; Su, Y.P.; Yu, C.X. Effects of koumine, an alkaloid of Gelsemium elegans Benth., on inflammatory and neuropathic pain models and possible mechanism with allopregnanolone. Pharm. Biochem. Bebahv. 2012, 101, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.C.R.; Kitagawa, R.R.; Raddi, M.S.G.; Carlos, I.Z.; Pombeiro-Sponchiado, S.R. Inhibition of nitric oxide and tumour necrosis factor-α production in peritoneal macrophages by Aspergillus nidulans Melanin. Biol. Pharm. Bull. 2013, 36, 1915–1920. [Google Scholar] [CrossRef]
- Regina, S.; Fausto, C.; Massimo, C.; Jamil, A.; Christoph, T. Neuronal nitric oxide synthase is involved in vascular hyporeactivity and multiple organ dysfunction associated with haemorrhagic Shock. Shock 2015. [Google Scholar] [CrossRef]
- Wong, S.S.; Zhou, H.R.; Marin-martinez, M.L.; Brooks, K.; Pestka, J.J. Modulation of IL-1b, IL-6 and TNF-a secretion and mRNA expression by the trichothecene vomitoxin in the RAW264.7 murine macrophage cell line. Food Chem. Toxicol. 1998, 36, 409–419. [Google Scholar] [CrossRef]
- Schrader, L.I.; Kinzenbaw, D.A.; Johnson, A.W.; Faraci, F.M.; Didion, S.P. IL-6 deficiency protects against angiotensin II induced endothelial dysfunction and hypertrophy. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2576–2581. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.R.; Wright, J.; Bauter, M.; Seweryniak, K.; Kode, A.; Rahman, I. Sirtuin regulates cigarette smoke-induced proinflammatory mediator release via RelA/p65 NF-κB in macrophages in vitro and in rat lungs in vivo: Implications for chronic inflammation and aging. Lung Cell. Mol. Physiol. 2007, 292, L567–L576. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Hong, S.G.; Lee, J.W.; Jeong, W.S. Red ginseng marc oil inhibits iNOS and COX-2 via NF-κB and p38 pathways in LPS-stimulated RAW264.7 macrophages. Molecules 2012, 17, 13769–13786. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Fujii, M.; Hou, D.X. 6-(Methylsulfinyl)hexyl isothiocyanate suppresses inducible nitric oxide synthase expression through the inhibition of Janus kinase 2-mediated JNK pathway in lipopolysaccharide-activated murine macrophages. Biochem. Pharmacol. 2005, 70, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, C.D.; Ceci, J.D.; Tsatsanis, C.; Kontoyiannis, D.; Stamatakis, K.; Lin, J.H.; Patriotis, C.; Jenkins, N.A.; Copeland, N.G.; Kollias, G.; et al. TNF-α induction by LPS is regulated posttranscriptionally via a Tpl2/ERK-dependent pathway. Cell 2000, 103, 1071–1083. [Google Scholar] [CrossRef]
- Hommes, D.W.; Peppelenbosch, M.P.; van Deventer, S.J. Mitogen activated protein (MAP) kinase signal transduction pathways and novel anti-inflammatory targets. Gut 2003, 52, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Robb, J.; Norval, M.; Neill, W. The use of tissue culture for the detection of mycotoxins. Lett. Appl. Microbiol. 1990, 10, 161–165. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Z.; Matias, F.B.; Wu, J.; Liang, Z.; Sun, Z. Koumine Attenuates Lipopolysaccaride-Stimulated Inflammation in RAW264.7 Macrophages, Coincidentally Associated with Inhibition of NF-κB, ERK and p38 Pathways. Int. J. Mol. Sci. 2016, 17, 430. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030430
Yuan Z, Matias FB, Wu J, Liang Z, Sun Z. Koumine Attenuates Lipopolysaccaride-Stimulated Inflammation in RAW264.7 Macrophages, Coincidentally Associated with Inhibition of NF-κB, ERK and p38 Pathways. International Journal of Molecular Sciences. 2016; 17(3):430. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030430
Chicago/Turabian StyleYuan, Zhihang, Froilan Bernard Matias, Jing Wu, Zengenni Liang, and Zhiliang Sun. 2016. "Koumine Attenuates Lipopolysaccaride-Stimulated Inflammation in RAW264.7 Macrophages, Coincidentally Associated with Inhibition of NF-κB, ERK and p38 Pathways" International Journal of Molecular Sciences 17, no. 3: 430. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030430