
Aripiprazole and Haloperidol Activate GSK3β-Dependent Signalling Pathway Differentially in Various Brain Regions of Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
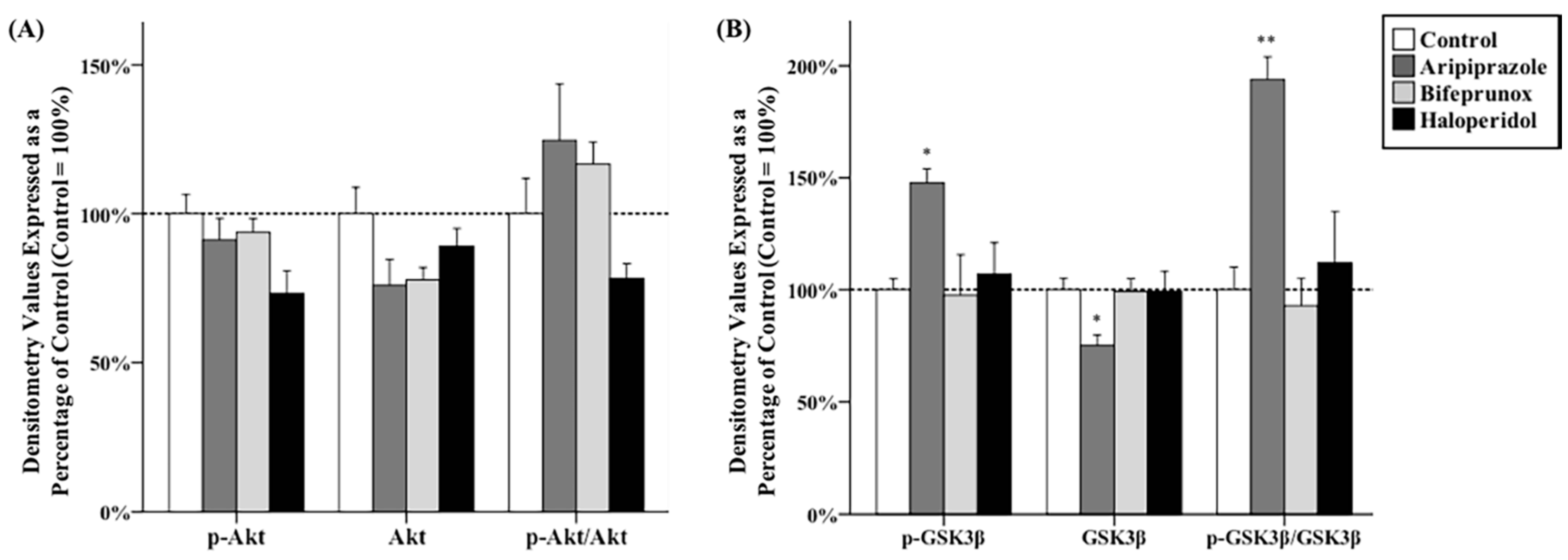
2.1. Effects of Antipsychotics in the Prefrontal Cortex
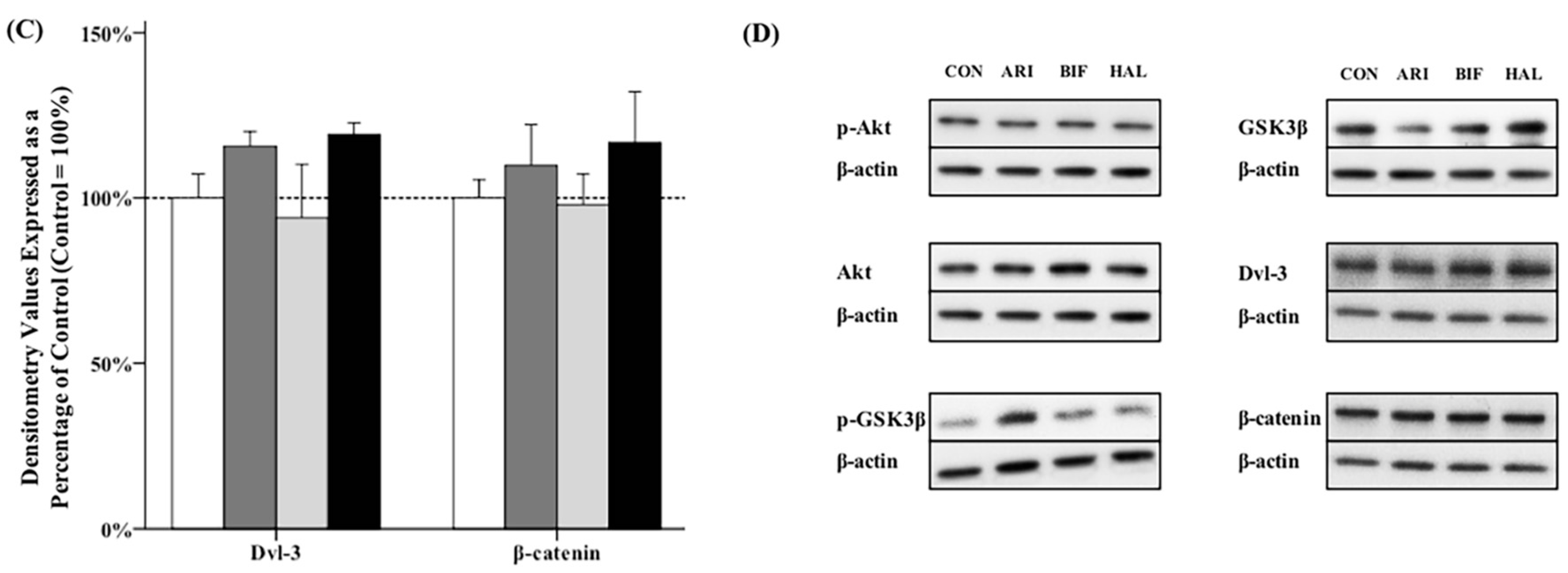
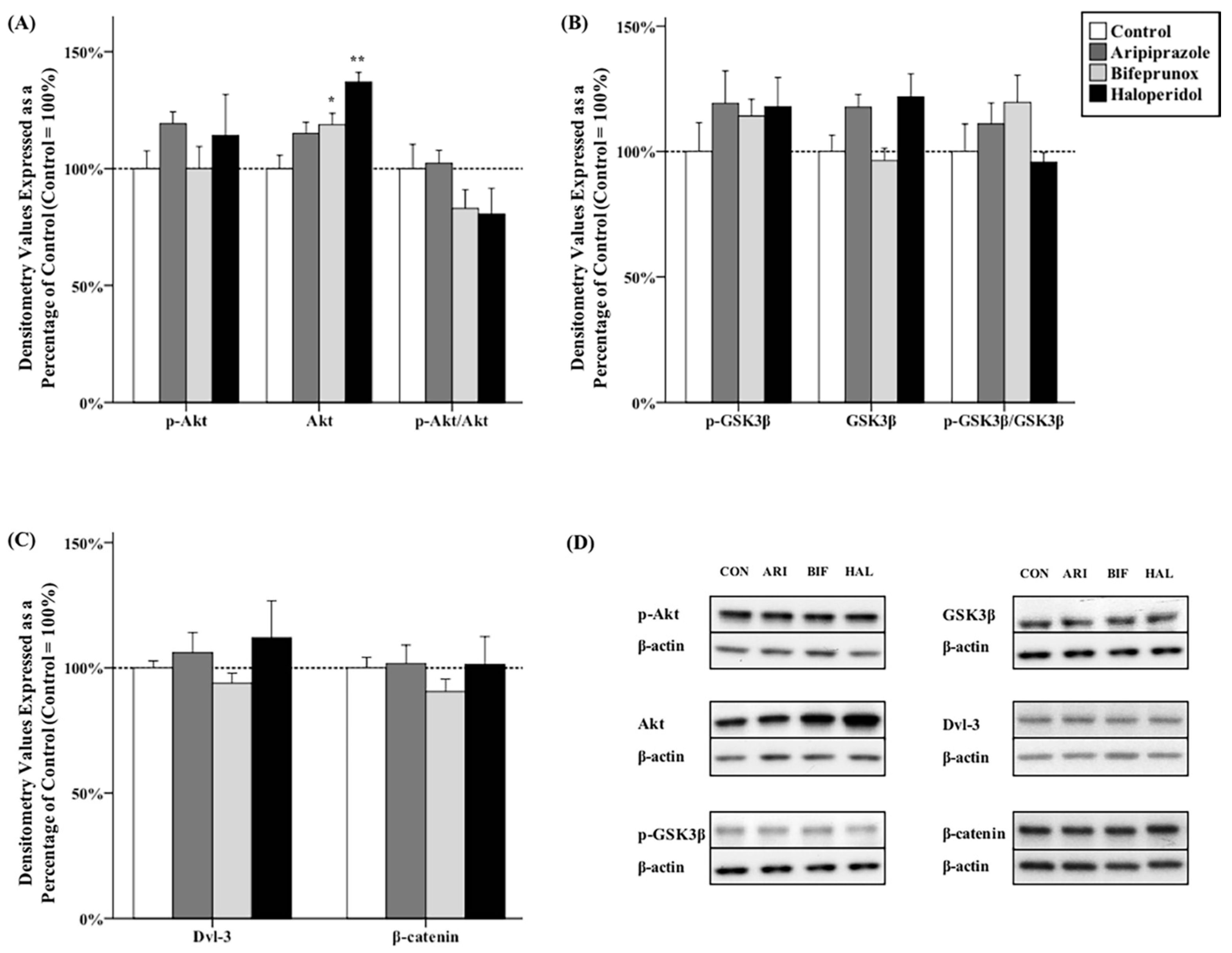
2.2. Effects of Antipsychotics in the Caudate Putamen
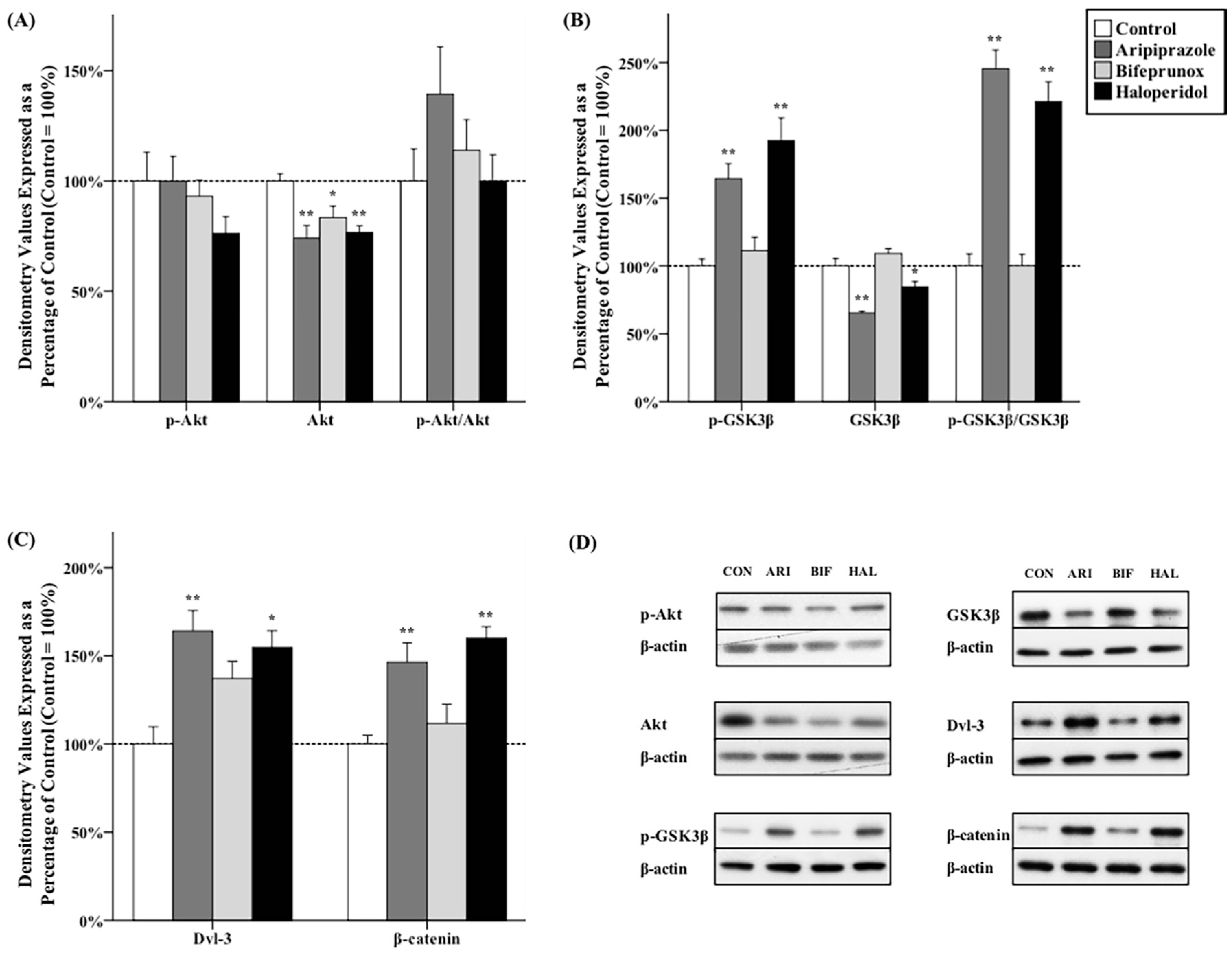
2.3. Effects of Antipsychotics in the Nucleus Accumbens
3. Discussion
4. Materials and Methods
4.1. Animals and Drug Administration
4.2. Micro-Dissection of Brain Samples
4.3. Western Blots
4.4. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Akt | protein kinase B |
| CPu | caudate putamen |
| D2R | dopamine D2 receptor |
| Dvl | dishevelled |
| EPS | extrapyramidal side-effects |
| GSK3β | glycogen synthase kinase 3β |
| NAc | nucleus accumbens |
| PFC | prefrontal cortex |
References
- Mailman, R.B.; Murthy, V. Third generation antipsychotic drugs: Partial agonism or receptor functional selectivity? Curr. Pharm. Des. 2010, 16, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Emamian, E.S. AKT/GSK3 signaling pathway and schizophrenia. Front. Mol. Neurosci. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.; McCutcheon, R.; Stone, J. Glutamate and dopamine in schizophrenia: An update for the 21st century. J. Psychopharmacol. 2015, 29, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Sotnikova, T.D.; Yao, W.D.; Kockeritz, L.; Woodgett, J.R.; Gainetdinov, R.R.; Caron, M.G. Lithium antagonizes dopamine-dependent behaviors mediated by an AKT/glycogen synthase kinase 3 signaling cascade. Proc. Natl. Acad. Sci. USA 2004, 101, 5099–5104. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Gainetdinov, R.R.; Caron, M.G. The Akt-GSK-3 signaling cascade in the actions of dopamine. Trends Pharmacol. Sci. 2007, 28, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Gainetdinov, R.R.; Caron, M.G. Akt/GSK3 signaling in the action of psychotropic drugs. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.K.; Lee, C.H.; Cho, H.Y.; You, Y.S.; Lee, B.J.; Lee, J.G.; Park, S.W.; Kim, Y.H. Effects of antipsychotic drugs on the expression of synapse-associated proteins in the frontal cortex of rats subjected to immobilization stress. Psychiatry Res. 2015, 229, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Chen, J.; Lian, J.; Huang, X.F.; Deng, C. Unique effects of acute aripiprazole treatment on the dopamine D2 receptor downstream cAMP-PKA and Akt-GSK3β signalling pathways in rats. PLoS ONE 2015, 10, e0132722. [Google Scholar] [CrossRef] [PubMed]
- Min, C.; Cho, D.I.; Kwon, K.J.; Kim, K.S.; Shin, C.Y.; Kim, K.M. Novel regulatory mechanism of canonical Wnt signaling by dopamine D2 receptor through direct interaction with β-catenin. Mol. Pharmacol. 2011, 80, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.P.; Honardoust, D.; Mouyal, J.; Rajakumar, N.; Rushlow, W.J. Activation of the canonical Wnt pathway by the antipsychotics haloperidol and clozapine involves dishevelled-3. J. Neurochem. 2007, 102, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Alimohamad, H.; Sutton, L.; Mouyal, J.; Rajakumar, N.; Rushlow, W.J. The effects of antipsychotics on β-catenin, glycogen synthase kinase-3 and dishevelled in the ventral midbrain of rats. J. Neurochem. 2005, 95, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Alimohamad, H.; Rajakumar, N.; Seah, Y.H.; Rushlow, W. Antipsychotics alter the protein expression levels of β-catenin and GSK-3 in the rat medial prefrontal cortex and striatum. Biol. Psychiatry 2005, 57, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Rosborough, K.M.; Friedman, A.B.; Zhu, W.; Roth, K.A. Regulation of mouse brain glycogen synthase kinase-3 by atypical antipsychotics. Int. J. Neuropsychopharmacol. 2007, 10, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.P.; Rushlow, W.J. The effects of neuropsychiatric drugs on glycogen synthase kinase-3 signaling. Neuroscience 2011, 199, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Phuong, V.T.; Lee, C.H.; Lee, J.G.; Seo, M.K.; Cho, H.Y.; Fang, Z.H.; Lee, B.J.; Kim, Y.H. Effects of antipsychotic drugs on BDNF, GSK-3β, and β-catenin expression in rats subjected to immobilization stress. Neurosci. Res. 2011, 71, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Hirose, T.; Kikuchi, T. Aripiprazole, a novel antipsychotic agent: Dopamine D2 receptor partial agonist. J. Med. Investig. 2005, 52, 284–290. [Google Scholar] [CrossRef]
- Burris, K.D. Aripiprazole, a novel antipsychotic, is a high-affinity partial agonist at human dopamine D2 receptors. J. Pharmacol. Exp. Ther. 2002, 302, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Wadenberg, M.-L.G. Bifeprunox: A novel antipsychotic agent with partial agonist properties at dopamine D2 and serotonin 5-HT1A receptors. Future Neurol. 2007, 2, 153–165. [Google Scholar] [CrossRef]
- Beasley, C.; Cotter, D.; Khan, N.; Pollard, C.; Sheppard, P.; Varndell, I.; Lovestone, S.; Anderton, B.; Everall, I. Glycogen synthase kinase-3β immunoreactivity is reduced in the prefrontal cortex in schizophrenia. Neurosci. Lett. 2001, 302, 117–120. [Google Scholar] [CrossRef]
- Koros, E.; Dorner-Ciossek, C. The role of glycogen synthase kinase-3β in schizophrenia. Drug News Perspect. 2007, 20, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Emamian, E.S.; Hall, D.; Birnbaum, M.J.; Karayiorgou, M.; Gogos, J.A. Convergent evidence for impaired AKT1-GSK3β signaling in schizophrenia. Nat. Genet. 2004, 36, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Tadori, Y.; Miwa, T.; Tottori, K.; Burris, K.D.; Stark, A.; Mori, T.; Kikuchi, T. Aripiprazole’s low intrinsic activities at human dopamine D2L and D2S receptors render it a unique antipsychotic. Eur. J. Pharmacol. 2005, 515, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Benkert, O.; Muller-Siecheneder, F.; Wetzel, H. Dopamine agonists in schizophrenia: A review. Eur. Neuropsychopharmacol. 1995, 5, 43–53. [Google Scholar] [CrossRef]
- Mikell, C.B.; McKhann, G.M.; Segal, S.; McGovern, R.A.; Wallenstein, M.B.; Moore, H. The hippocampus and nucleus accumbens as potential therapeutic targets for neurosurgical intervention in schizophrenia. Stereotact. Funct. Neurosurg. 2009, 87, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Roh, M.S.; Seo, M.S.; Kim, Y.; Kim, S.H.; Jeon, W.J.; Ahn, Y.M.; Kang, U.G.; Juhnn, Y.S.; Kim, Y.S. Haloperidol and clozapine differentially regulate signals upstream of glycogen synthase kinase 3 in the rat frontal cortex. Exp. Mol. Med. 2007, 39, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.C.; McEwen, H.; Steinberg, J.D.; Shepherd, P.R. The activation of the Akt/PKB signalling pathway in the brains of clozapine-exposed rats is linked to hyperinsulinemia and not a direct drug effect. Psychopharmacology 2014, 231, 4553–4560. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Sotnikova, T.D.; Marion, S.; Lefkowitz, R.J.; Gainetdinov, R.R.; Caron, M.G. An Akt/β-arrestin 2/PP2A signaling complex mediates dopaminergic neurotransmission and behavior. Cell 2005, 122, 261–273. [Google Scholar] [CrossRef] [PubMed]
- DeLeon, A.; Patel, N.C.; Crismon, M.L. Aripiprazole: A comprehensive review of its pharmacology, clinical efficacy, and tolerability. Clin. Ther. 2004, 26, 649–666. [Google Scholar] [CrossRef]
- Tadori, Y.; Kitagawa, H.; Forbes, R.A.; McQuade, R.D.; Stark, A.; Kikuchi, T. Differences in agonist/antagonist properties at human dopamine D(2) receptors between aripiprazole, bifeprunox and SDZ 208-912. Eur. J. Pharmacol. 2007, 574, 103–111. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services; Food and Drug Administration; Center for Drug Evaluation and Research. Food and Drug Administration. Estimating the maximum safe starting dose in initial clinical trials for therapeutics in adult healthy volunteers. In Guidance for Industry; Food and Drug Administration: Rockville, MD, USA, 2005. [Google Scholar]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.; Pan, B.; Lian, J.; Huang, X.F.; Deng, C. Different effects of bifeprunox, aripiprazole, and haloperidol on body weight gain, food and water intake, and locomotor activity in rats. Pharmacol. Biochem. Behav. 2014, 124, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Pan, B.; Hu, C.H.; Han, M.; Huang, X.F. Differential effects of short- and long-term antipsychotic treatment on the expression of neuregulin-1 and ErbB4 receptors in the rat brain. Psychiatry Res. 2015, 225, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Mace, S.; Taylor, D. Aripiprazole: Dose-response relationship in schizophrenia and schizoaffective disorder. CNS Drugs 2009, 23, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Casey, D.E.; Sands, E.E.; Heisterberg, J.; Yang, H.M. Efficacy and safety of bifeprunox in patients with an acute exacerbation of schizophrenia: Results from a randomized, double-blind, placebo-controlled, multicenter, dose-finding study. Psychopharmacology 2008, 200, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Emsley, R. Drugs in development for the treatment of schizophrenia. Expert Opin. Investig. Drugs 2009, 18, 1103–1118. [Google Scholar] [CrossRef] [PubMed]
- Kapur, S.; VanderSpek, S.C.; Brownlee, B.A.; Nobrega, J.N. Antipsychotic dosing in preclinical models is often unrepresentative of the clinical condition: A suggested solution based on in vivo occupancy. J. Pharmacol. Exp. Ther. 2003, 305, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Huang, X.F.; Deng, C. Aripiprazole differentially affects mesolimbic and nigrostriatal dopaminergic transmission: Implications for long-term drug efficacy and low extrapyramidal side-effects. Int. J. Neuropsychopharmacol. 2009, 12, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Assie, M.B.; Dominguez, H.; Consul-Denjean, N.; Newman-Tancredi, A. In vivo occupancy of dopamine D2 receptors by antipsychotic drugs and novel compounds in the mouse striatum and olfactory tubercles. Naunyn Schmiedebergs Arch. Pharmacol. 2006, 373, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Natesan, S.; Reckless, G.E.; Nobrega, J.N.; Fletcher, P.J.; Kapur, S. Dissociation between in vivo occupancy and functional antagonism of dopamine D2 receptors: Comparing aripiprazole to other antipsychotics in animal models. Neuropsychopharmacology 2006, 31, 1854–1863. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Elsevier Academic Press: San Diego, CA, USA, 2005. [Google Scholar]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, B.; Huang, X.-F.; Deng, C. Aripiprazole and Haloperidol Activate GSK3β-Dependent Signalling Pathway Differentially in Various Brain Regions of Rats. Int. J. Mol. Sci. 2016, 17, 459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17040459
Pan B, Huang X-F, Deng C. Aripiprazole and Haloperidol Activate GSK3β-Dependent Signalling Pathway Differentially in Various Brain Regions of Rats. International Journal of Molecular Sciences. 2016; 17(4):459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17040459
Chicago/Turabian StylePan, Bo, Xu-Feng Huang, and Chao Deng. 2016. "Aripiprazole and Haloperidol Activate GSK3β-Dependent Signalling Pathway Differentially in Various Brain Regions of Rats" International Journal of Molecular Sciences 17, no. 4: 459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17040459