
Differential MicroRNA Expression Profile in Myxomatous Mitral Valve Prolapse and Fibroelastic Deficiency Valves

 ,
,
Abstract
:1. Introduction
2. Results
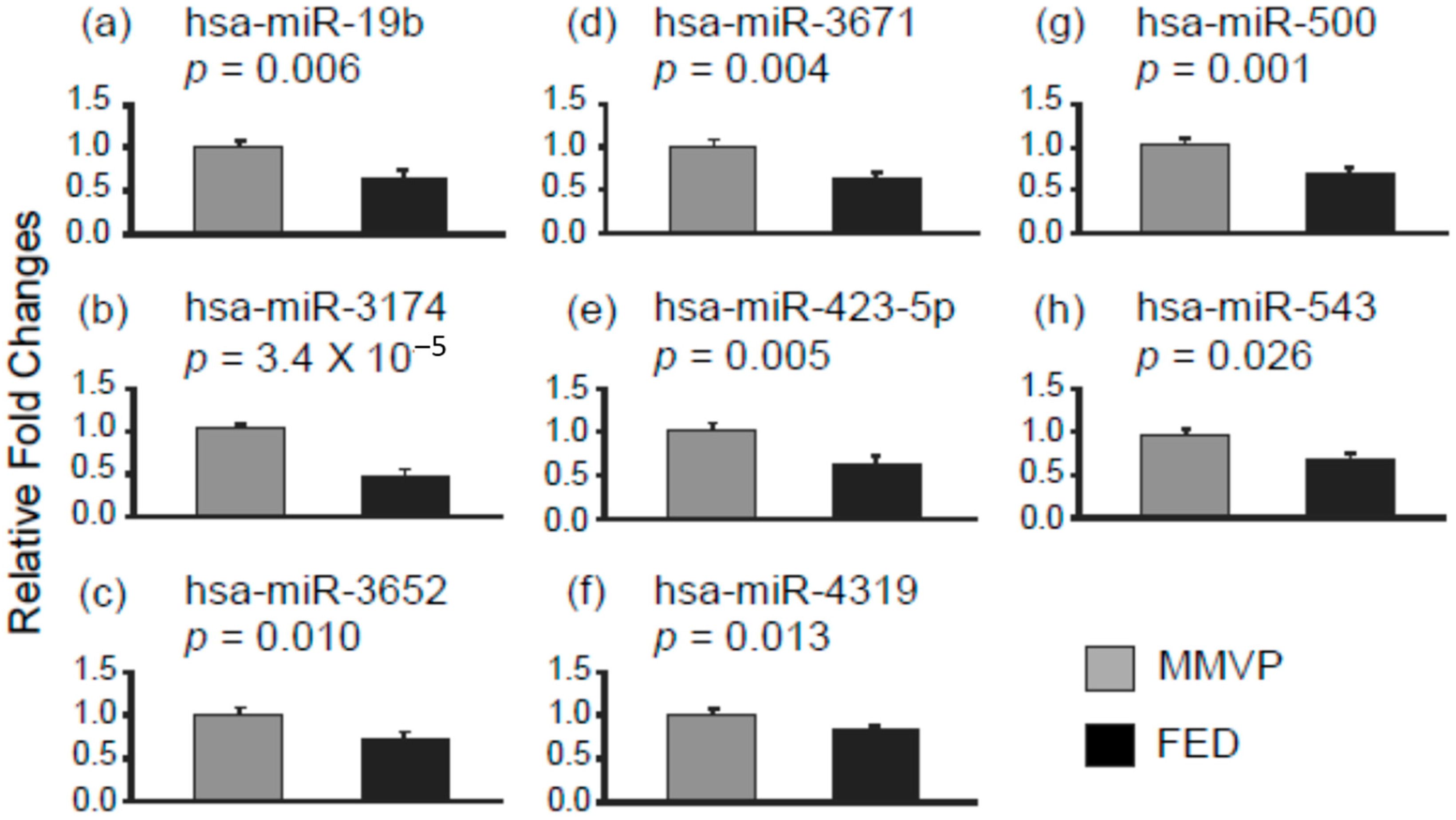
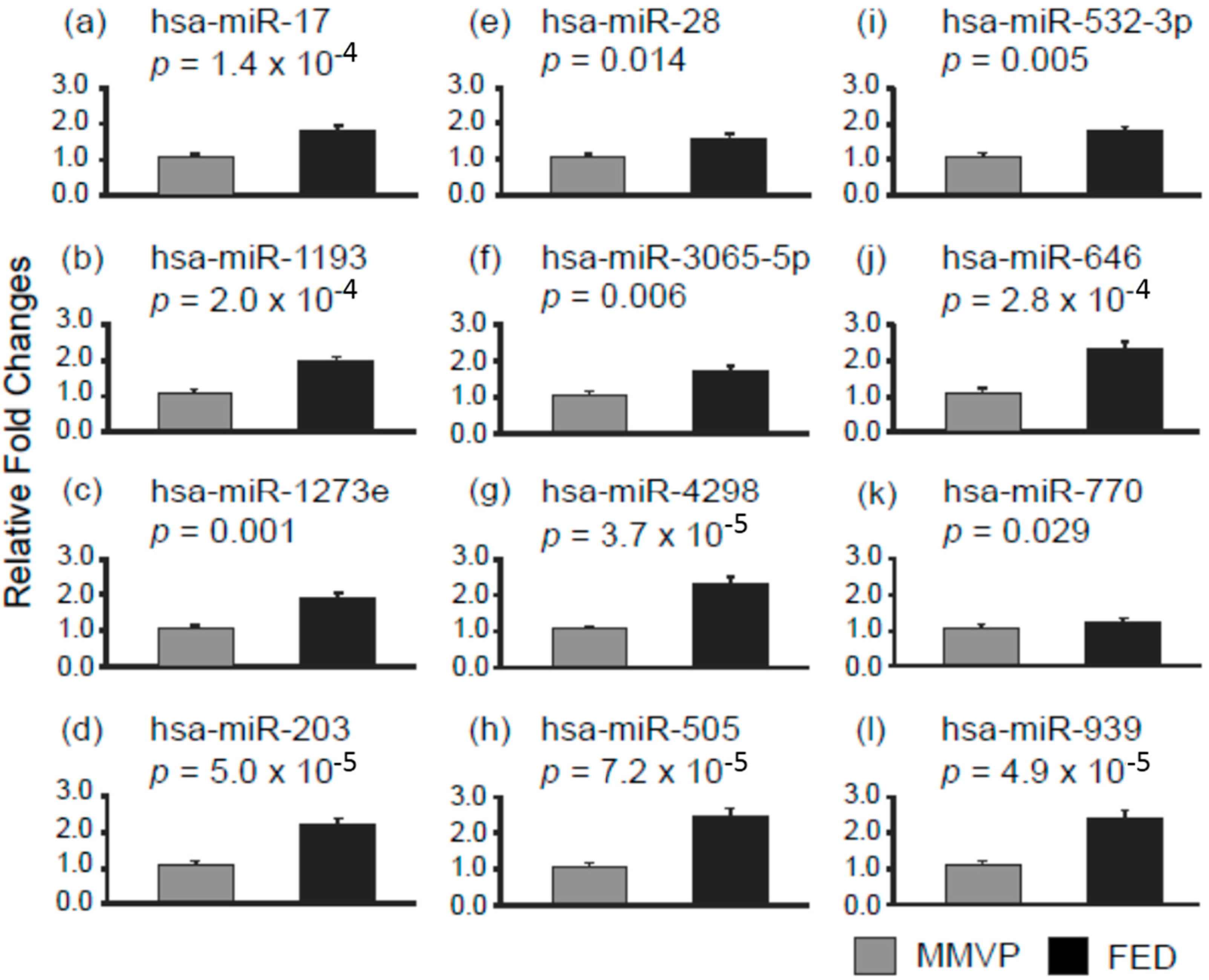
2.1. Differential MicroRNA Expression in Myxomatous Mitral Valve Prolapse (MMVP) and Fibroelastic Deficiency (FED) Mitral Valves
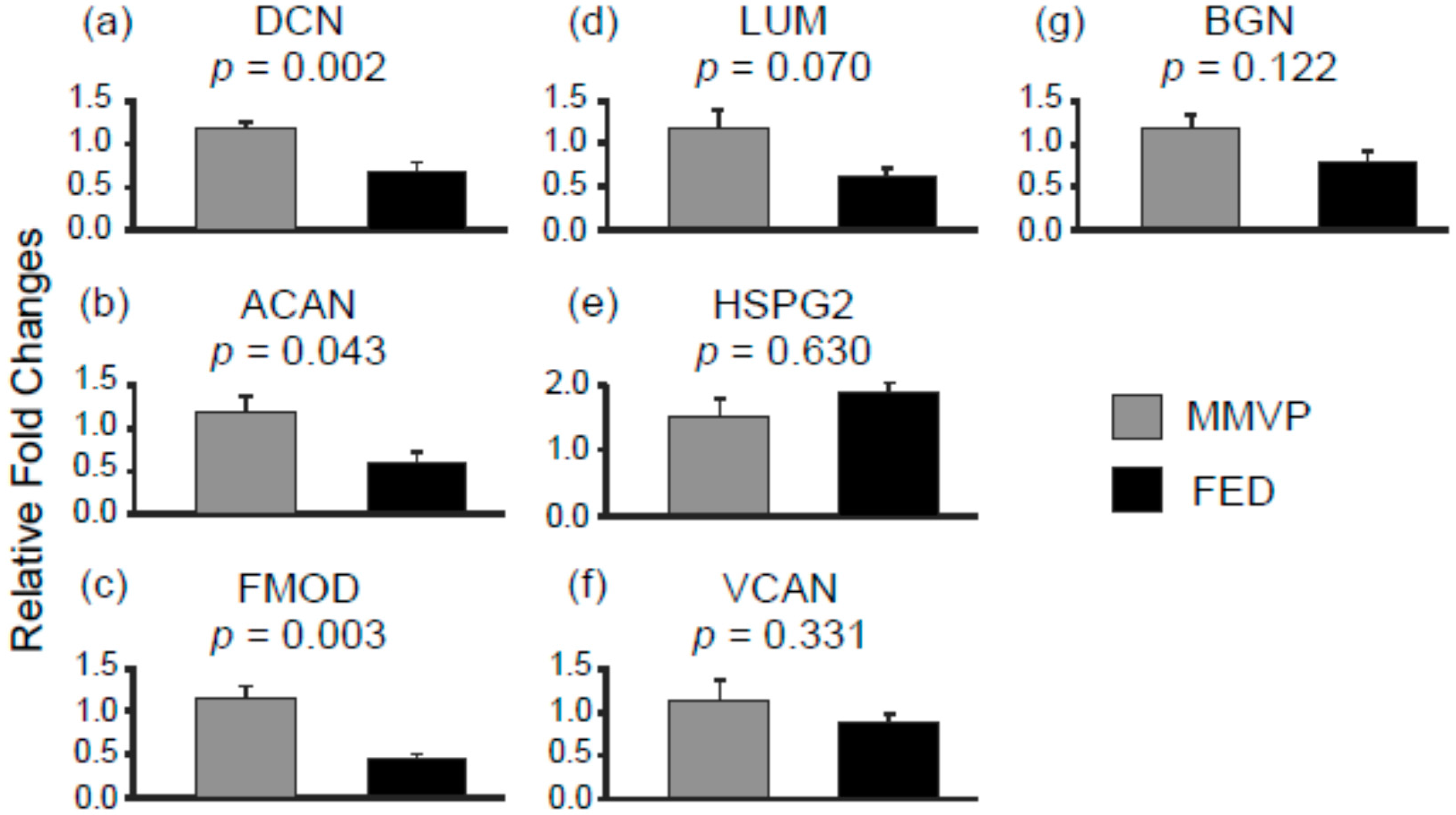
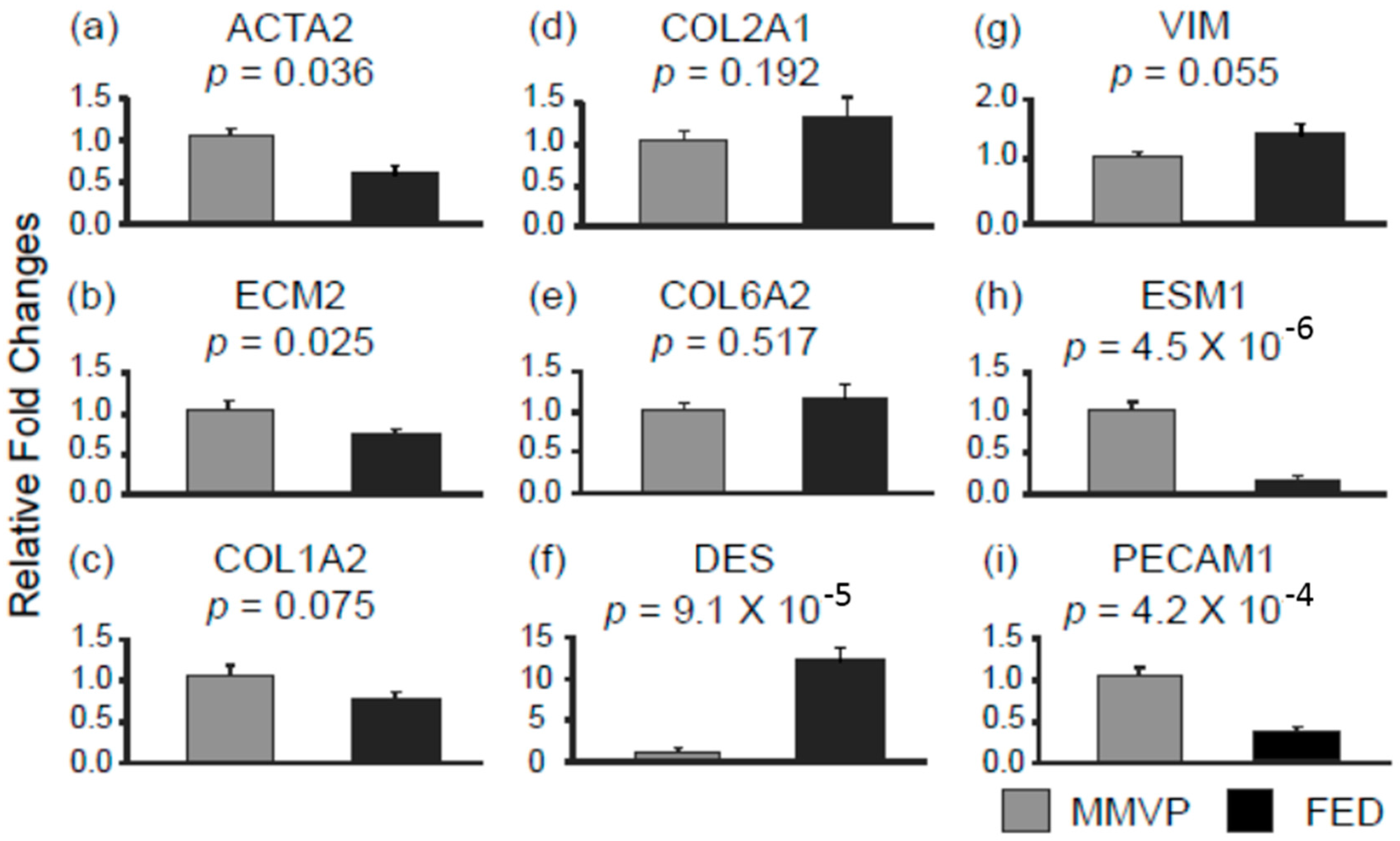
2.2. Differential mRNA Expression in MMVP and FED Mitral Valves
3. Discussion
4. Materials and Methods
4.1. Mitral Valve Tissues
4.2. Total RNA Extraction and Gene Expression Analysis
4.3. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MMVP | myxomatous mitral valve proplase |
| FED | fibroelastic deficiency disease |
| DMVD | degererative mitral valve disease |
| VHD | valvular heart disease |
| 3′UTR | 3′ end untranslated region |
| DIANA | DNA intelligent Analysis |
| KEGG pathways | Kyoto Encyclopedia of Genes and Genomes pathways |
| DCN | decorin |
| ACAN | aggrecan |
| FMOD | fibromodulin |
| LUM | lumican |
| HSPG2 | heparin sulfate proteoglycan 2 |
| VCAN | versican |
| BGN | biglycan |
| ACTA2 | α actin 2 |
| ECM2 | extracellular matrix protein 2 |
| COL1A2 | colleagen type 1 α 2 |
| COL2A1 | colleagen type 2 α 1 |
| COL6A2 | colleagen type 6 α 2 |
| DES | desmin |
| VIM | vimentin |
| ESM1 | endothelial cell specific molecule 1 |
| PECAM1 | platelet/ endothelial cell adhesion molecule 1 |
| VIC | valvular interstitial cell |
References
- Nkomo, V.T.; Gardin, J.M.; Skelton, T.N.; Gottdiener, J.S.; Scott, C.G.; Enriquez-Sarano, M. Burden of valvular heart diseases: A population-based study. Lancet 2006, 368, 1005–1011. [Google Scholar] [CrossRef]
- Iung, B.; Vahanian, A. Epidemiology of valvular heart disease in the adult. Nat. Rev. Cardiol. 2011, 8, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.B.; Bosman, C.K. Aneurysmal protrusion of the posterior leaflet of the mitral valve: An auscultatory-electrocardiographic syndrome. Am. Heart J. 1966, 71, 166–178. [Google Scholar] [CrossRef]
- Anyanwu, A.C.; Adams, D.H. Etiologic classification of degenerative mitral valve disease: Barlow’s disease and fibroelastic deficiency. Semin. Thorac. Cardiovasc. Surg. 2007, 19, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.H.; Rosenhek, R.; Falk, V. Degenerative mitral valve regurgitation: Best practice revolution. Eur. Heart J. 2010, 31, 1958–1966. [Google Scholar] [CrossRef] [PubMed]
- Fornes, P.; Heudes, D.; Fuzellier, J.F.; Tixier, D.; Bruneval, P.; Carpentier, A. Correlation between clinical and histologic patterns of degenerative mitral valve insufficiency: A histomorphometric study of 130 excised segments. Cardiovasc. Pathol. 1999, 8, 81–92. [Google Scholar] [CrossRef]
- Carpentier, A.; Chauvaud, S.; Fabiani, J.N.; Deloche, A.; Relland, J.; Lessana, A.; D’Allaines, C.; Blondeau, P.; Piwnica, A.; Dubost, C. Reconstructive surgery of mitral valve incompetence: Ten-year appraisal. J. Thorac. Cardiovasc. Surg. 1980, 79, 338–348. [Google Scholar] [PubMed]
- Eriksson, M.J.; Bitkover, C.Y.; Omran, A.S.; David, T.E.; Ivanov, J.; Ali, M.J.; Woo, A.; Siu, S.C.; Rakowski, H. Mitral annular disjunction in advanced myxomatous mitral valve disease: Echocardiographic detection and surgical correction. J. Am. Soc. Echocardiogr. 2005, 18, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Matsumaru, I.; Eishi, K.; Hashizume, K.; Kawano, H.; Tsuneto, A.; Hayashi, T. Clinical and pathological features of degenerative mitral valve disease: billowing mitral leaflet versus fibroelastic deficiency. Ann. Thorac. Cardiovasc. Surg. 2013. [Google Scholar] [CrossRef] [PubMed]
- Vaideeswara, P.; Butany, J. Valvular Heart Disease. In Cardiovascular Pathology, 4th ed.; Buja, L.M., Butany, J., Eds.; Elsevier: Oxford, UK, 2016; pp. 509–513. [Google Scholar]
- Disse, S.; Abergel, E.; Berrebi, A.; Houot, A.M.; Le Heuzey, J.Y.; Diebold, B.; Guize, L.; Carpentier, A.; Corvol, P.; Jeunemaitre, X. Mapping of a first locus for autosomal dominant myxomatous mitral-valve prolapse to chromosome 16p11.2-p12.1. Am. J. Hum. Genet. 1999, 65, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Freed, L.A.; Acierno, J.S.; Acierno, J.S., Jr.; Dai, D.; Leyne, M.; Marshall, J.E.; Nesta, F.; Levine, R.A.; Slaugenhaupt, S.A. A locus for autosomal dominant mitral valve prolapse on chromosome 11p15.4. Am. J. Hum. Genet. 2003, 72, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Nesta, F.; Leyne, M.; Yosefy, C.; Simpson, C.; Dai, D.; Marshall, J.E.; Hung, J.; Slaugenhaupt, S.A.; Levine, R.A. New locus for autosomal dominant mitral valve prolapse on chromosome 13: Clinical insights from genetic studies. Circulation 2005, 112, 2022–2030. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, F.; Schott, J.J.; Trochu, J.N.; Baranger, F.; Herbert, O.; Scott, V.; Fressinaud, E.; David, A.; Moisan, J.P.; Bouhour, J.B.; et al. Mapping of X-linked myxomatous valvular dystrophy to chromosome Xq28. Am. J. Hum. Genet. 1998, 62, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Durst, R.; Sauls, K.; Peal, D.S.; Toomer, K.; Leyne, M.; Salani, M.; Talkowski, M.E.; Brand, H.; Perrocheau, M.; Simpson, C.; et al. Mutations in DCHS1 cause mitral valve prolapse. Nature 2015, 525, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Dina, C.; Bouatia-Naji, N.; Tucker, N.; Delling, F.N.; Toomer, K.; Durst, R.; Perrocheau, M.; Fernandez-Friera, L.; Solis, J.; Le Tourneau, T.; et al. Genetic association analyses highlight biological pathways underlying mitral valve prolapse. Nat. Genet. 2015, 47, 1206–1211. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, R.A.; Otto, C.M.; Bonow, R.O.; Carabello, B.A.; Erwin, J.P., 3rd.; Guyton, R.A.; O’Gara, P.T.; Ruiz, C.E.; Skubas, N.J.; Sorajja, P.; et al. 2014 AHA/ACC guideline for the management of patients with valvular heart disease: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J. Am. Coll. Cardiol. 2014, 63, 2438–2488. [Google Scholar] [PubMed]
- Adams, D.H.; Anyanwu, A.C. Seeking a higher standard for degenerative mitral valve repair: Begin with etiology. J. Thorac. Cardiovasc. Surg. 2008, 136, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Flameng, W.; Meuris, B.; Herijgers, P.; Herregods, M.C. Durability of mitral valve repair in Barlow disease versus fibroelastic deficiency. J. Thorac. Cardiovasc. Surg. 2008, 135, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.L.; Armugam, A.; Sepramaniam, S.; Karolina, D.S.; Lim, K.Y.; Lim, J.Y.; Chong, J.P.; Ng, J.Y.; Chen, Y.T.; Chan, M.M.; et al. Circulating microRNAs in heart failure with reduced and preserved left ventricular ejection fraction. Eur. J. Heart Fail. 2015, 17, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.L.; Wee, A.S.; Lim, J.Y.; Ng, J.Y.; Chong, J.P.; Liew, O.W.; Lilyanna, S.; Martinez, E.C.; Ackers-Johnson, M.A.; Vardy, L.A.; et al. Natriuretic peptide receptor 3 (NPR3) is regulated by microRNA-100. J. Mol. Cell. Cardiol. 2015, 82, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- McNeill, E.; van Vactor, D. MicroRNAs shape the neuronal landscape. Neuron 2012, 75, 363–379. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.D.; Ambros, V. Circulating microRNAs in cardiovascular disease. Circulation 2011, 124, 1908–1910. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, G.; Latronico, M.V.; Cavarretta, E. MicroRNAs in cardiovascular diseases: Current knowledge and the road ahead. J. Am. Coll. Cardiol. 2014, 63, 2177–2187. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, B.; Lovren, F.; Pan, Y.; Garg, V.; Quan, A.; Tang, G.; Singh, K.K.; Shukla, P.C.; Kalra, N.P.; Peterson, M.D.; et al. miRNA-141 is a novel regulator of BMP-2-mediated calcification in aortic stenosis. J. Thorac. Cardiovasc. Surg. 2012, 144, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Kostoulas, N.; Vergoulis, T.; Georgakilas, G.; Reczko, M.; Maragkakis, M.; Paraskevopoulou, M.D.; Prionidis, K.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA miRPath v.2.0: Investigating the combinatorial effect of microRNAs in pathways. Nucleic Acids Res. 2012, 40, W498–W504. [Google Scholar] [CrossRef] [PubMed]
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. miRWalk–Database: Prediction of possible miRNA binding sites by “walking” the genes of three genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Prunotto, M.; Caimmi, P.P.; Bongiovanni, M. Cellular pathology of mitral valve prolapse. Cardiovasc. Pathol. 2010, 19, e113–e117. [Google Scholar] [CrossRef] [PubMed]
- Bauters, C.; Kumarswamy, R.; Holzmann, A.; Bretthauer, J.; Anker, S.D.; Pinet, F.; Thum, T. Circulating miR-133a and miR-423–5p fail as biomarkers for left ventricular remodeling after myocardial infarction. Int. J. Cardiol. 2013, 168, 1837–1840. [Google Scholar] [CrossRef] [PubMed]
- Goren, Y.; Kushnir, M.; Zafrir, B.; Tabak, S.; Lewis, B.S.; Amir, O. Serum levels of microRNAs in patients with heart failure. Eur. J. Heart Fail. 2012, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Tijsen, A.J.; Creemers, E.E.; Moerland, P.D.; de Windt, L.J.; van der Wal, A.C.; Kok, W.E.; Pinto, Y.M. miR423-5p as a circulating biomarker for heart failure. Circ. Res. 2010, 106, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, L.; Wang, H.; Xu, G.; Zhu, S.; Li, M.; Hu, X.; Zhu, J.; Zhu, C.; Xu, J.; et al. Effects of miR-19b knockdown on the cardiac differentiation of P19 mouse embryonic carcinoma cells. Mol. Med. Rep. 2015, 11, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Hurlstone, A.F.; Haramis, A.P.; Wienholds, E.; Begthel, H.; Korving, J.; van Eeden, F.; Cuppen, E.; Zivkovic, D.; Plasterk, R.H.; Clevers, H. The Wnt/β-catenin pathway regulates cardiac valve formation. Nature 2003, 425, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.D.; Tian, Y.; Morrisey, E.E. Wnt signaling: An essential regulator of cardiovascular differentiation, morphogenesis and progenitor self-renewal. Development 2008, 135, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.C.; Joag, V.R.; Gotlieb, A.I. The emerging role of valve interstitial cell phenotypes in regulating heart valve pathobiology. Am. J. Pathol. 2007, 171, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Mannion, S.; Mtintsilana, A.; Posthumus, M.; van der Merwe, W.; Hobbs, H.; Collins, M.; September, A.V. Genes encoding proteoglycans are associated with the risk of anterior cruciate ligament ruptures. Br. J. Sports Med. 2014, 48, 1640–1646. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Information | MMVP (n = 10) | FED (n = 10) | p Value |
|---|---|---|---|
| Age, years | 57.50 ± 3.76 | 59.10 ± 3.05 | 0.758 |
| Female gender | 3/10 (30%) | 1/10 (10%) | - |
| NYHA class | 2.40 ± 0.38 | 2.00 ± 0.32 | 0.453 |
| Atrial fibrillation rhythm | 3 (30%) | 2 (20%) | - |
| LV diastolic dimension, mm | 57.60 ± 2.22 | 59.80 ± 2.29 | 0.521 |
| LV systolic dimension, mm | 35.90 ± 2.05 | 36.20 ± 1.69 | 0.916 |
| LV ejection fraction, % | 61.80 ± 2.70 | 66.50 ± 2.10 | 0.210 |
| Left atrial volume index, mL/m2 | 93.78 ± 9.15 | 90.60 ± 7.28 | 0.807 |
| MA dimension, mm | 39.90 ± 0.87 | 34.20 ± 1.77 * | 0.016 |
| MA dimension/BSA, mm/m2 | 24.04 ± 1.36 | 19.81 ± 0.55 * | 0.019 |
| Severe mitral regurgitation | 9 (90%) | 10 (100%) | - |
| Mitral regurgitant volume, mL | 92.00 ± 12.36 | 100.44 ± 11.38 | 0.649 |
| Effective regurgitant orifice, cm2 | 0.67 ± 0.11 | 0.71 ± 0.09 | 0.825 |
| AML length, mm | 31.90 ± 1.65 | 23.44 ± 0.87 * | 0.0008 |
| AML thickness, mm | 4.68 ± 0.38 | 2.86 ± 0.11 * | 0.0013 |
| PML length, mm | 21.10 ± 1.67 | 18.22 ± 1.40 | 0.238 |
| PML thickness, mm | 4.49 ± 0.38 | 3.56 ± 0.39 | 0.133 |
| ID | Fold Change (FED/MMVP) | ID | Fold Change (FED/MMVP) |
|---|---|---|---|
| Decorin (DCN) | 0.6 | hsa-miR-203 | 2.1 |
| hsa-miR-505 | 2.4 | ||
| hsa-miR-532-3p | 1.7 | ||
| hsa-miR-770-5p | 1.2 | ||
| Aggrecan (ACAN) | 0.5 | hsa-miR-1273e | 1.8 |
| hsa-miR-203 | 2.1 | ||
| hsa-miR-532-3p | 1.7 | ||
| hsa-miR-646 | 2.2 | ||
| hsa-miR-770-5p | 1.2 | ||
| hsa-miR-939 | 2.3 | ||
| Fibromodulin (FMOD) | 0.4 | hsa-miR-1273e | 1.8 |
| hsa-miR-17 | 1.7 | ||
| hsa-miR-203 | 2.1 | ||
| hsa-miR-28 | 1.5 | ||
| hsa-miR-532-3p | 1.7 | ||
| hsa-miR-770-5p | 1.2 | ||
| hsa-miR-939 | 2.3 |
| ID | Fold Change (FED/MMVP) | ID | Fold Change (FED/MMVP) |
|---|---|---|---|
| ACTA2 | 0.6 | hsa-miR-17 | 1.7 |
| hsa-miR-28-5p | 1.5 | ||
| ECM2 | 0.7 | hsa-miR-203 | 2.1 |
| DES | 10.3 | hsa-miR-423-5p | 0.6 |
| hsa-miR-543 | 0.6 | ||
| ESM1 | 0.15 | hsa-miR-203 | 2.1 |
| hsa-miR-28-5p | 1.5 | ||
| hsa-miR-505 | 2.4 | ||
| hsa-miR-532-3p | 1.7 | ||
| hsa-miR-646 | 2.2 | ||
| VIM | 1.4 # | hsa-miR-19b | 0.6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-T.; Wang, J.; Wee, A.S.Y.; Yong, Q.-W.; Tay, E.L.-W.; Woo, C.C.; Sorokin, V.; Richards, A.M.; Ling, L.-H. Differential MicroRNA Expression Profile in Myxomatous Mitral Valve Prolapse and Fibroelastic Deficiency Valves. Int. J. Mol. Sci. 2016, 17, 753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050753
Chen Y-T, Wang J, Wee ASY, Yong Q-W, Tay EL-W, Woo CC, Sorokin V, Richards AM, Ling L-H. Differential MicroRNA Expression Profile in Myxomatous Mitral Valve Prolapse and Fibroelastic Deficiency Valves. International Journal of Molecular Sciences. 2016; 17(5):753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050753
Chicago/Turabian StyleChen, Yei-Tsung, Juan Wang, Abby S. Y. Wee, Quek-Wei Yong, Edgar Lik-Wui Tay, Chin Cheng Woo, Vitaly Sorokin, Arthur Mark Richards, and Lieng-Hsi Ling. 2016. "Differential MicroRNA Expression Profile in Myxomatous Mitral Valve Prolapse and Fibroelastic Deficiency Valves" International Journal of Molecular Sciences 17, no. 5: 753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050753