
Integrated Analysis of Expression Profile Based on Differentially Expressed Genes in Middle Cerebral Artery Occlusion Animal Models

Abstract
:
1. Introduction
2. Results
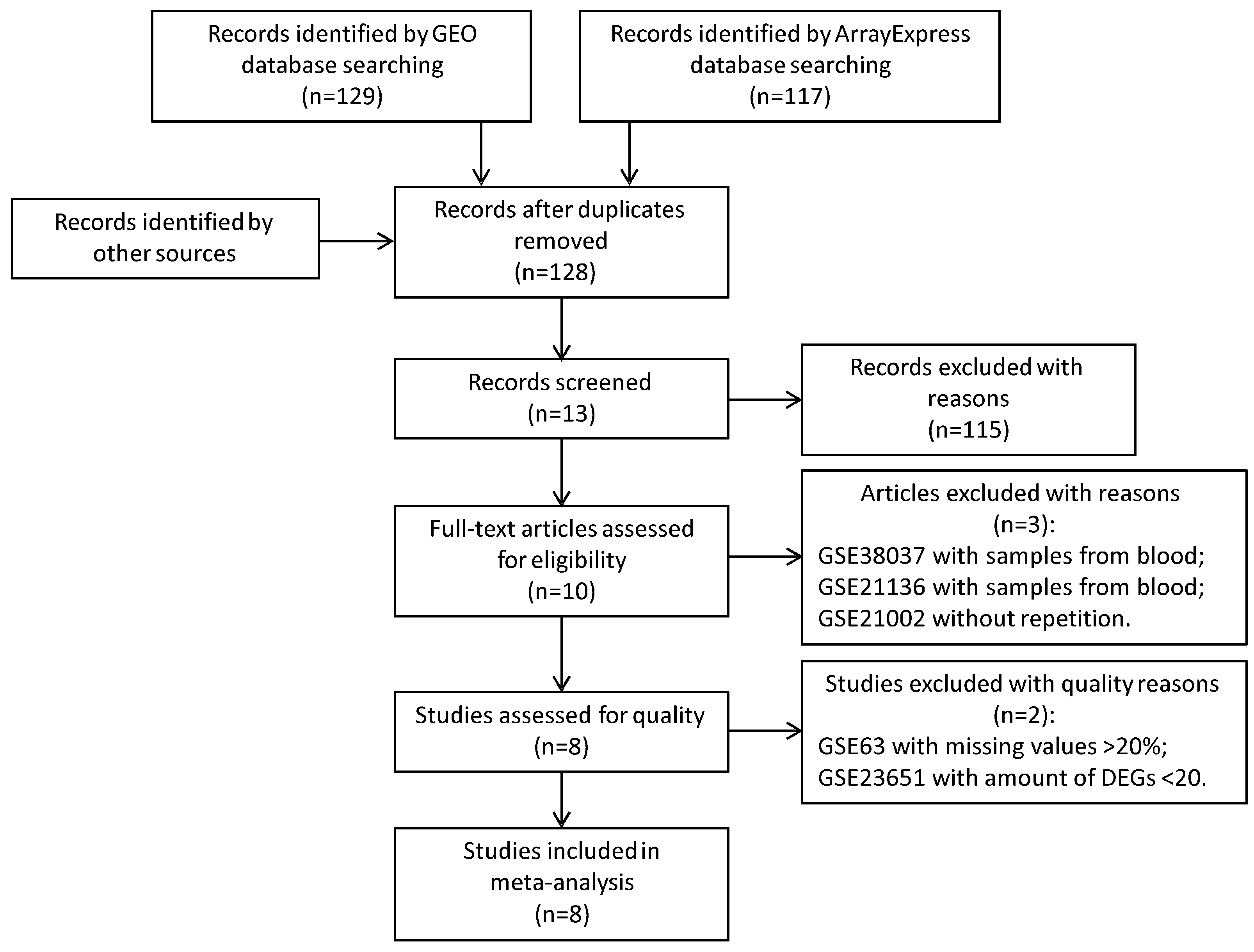
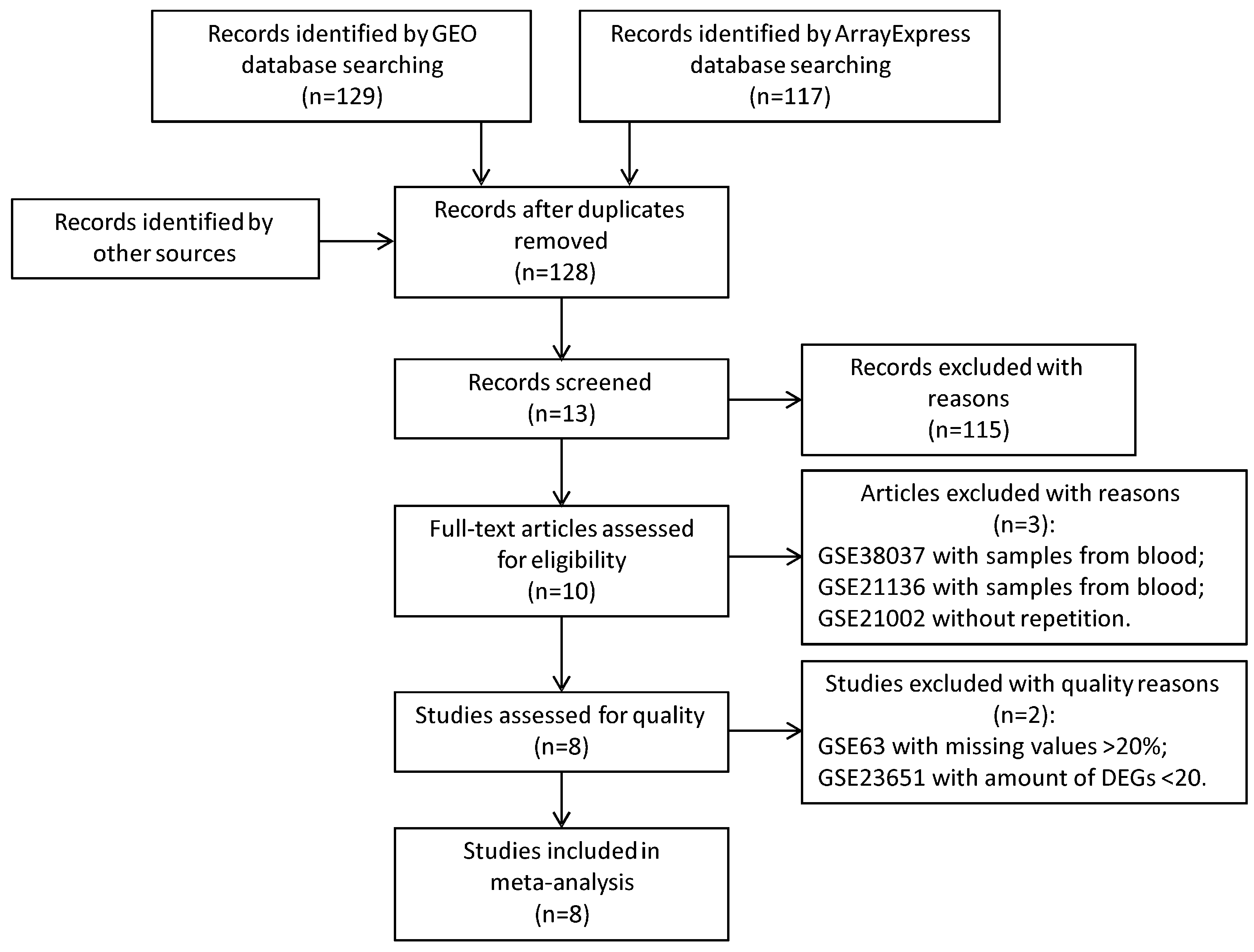
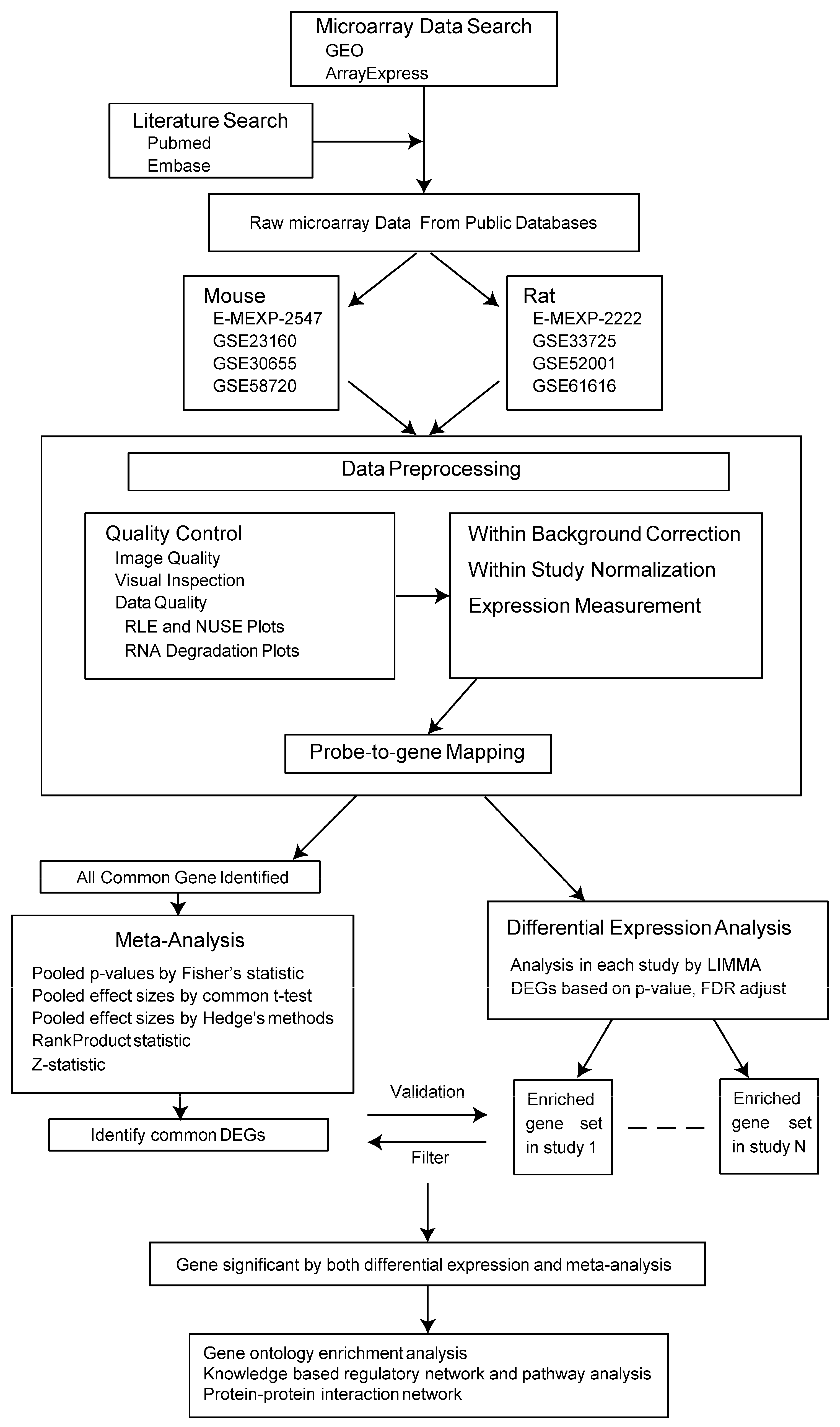
2.1. Microarray Data Search and Data Preprocessing
2.1.1. Microarray Data Search
2.1.2. Data Preprocessing
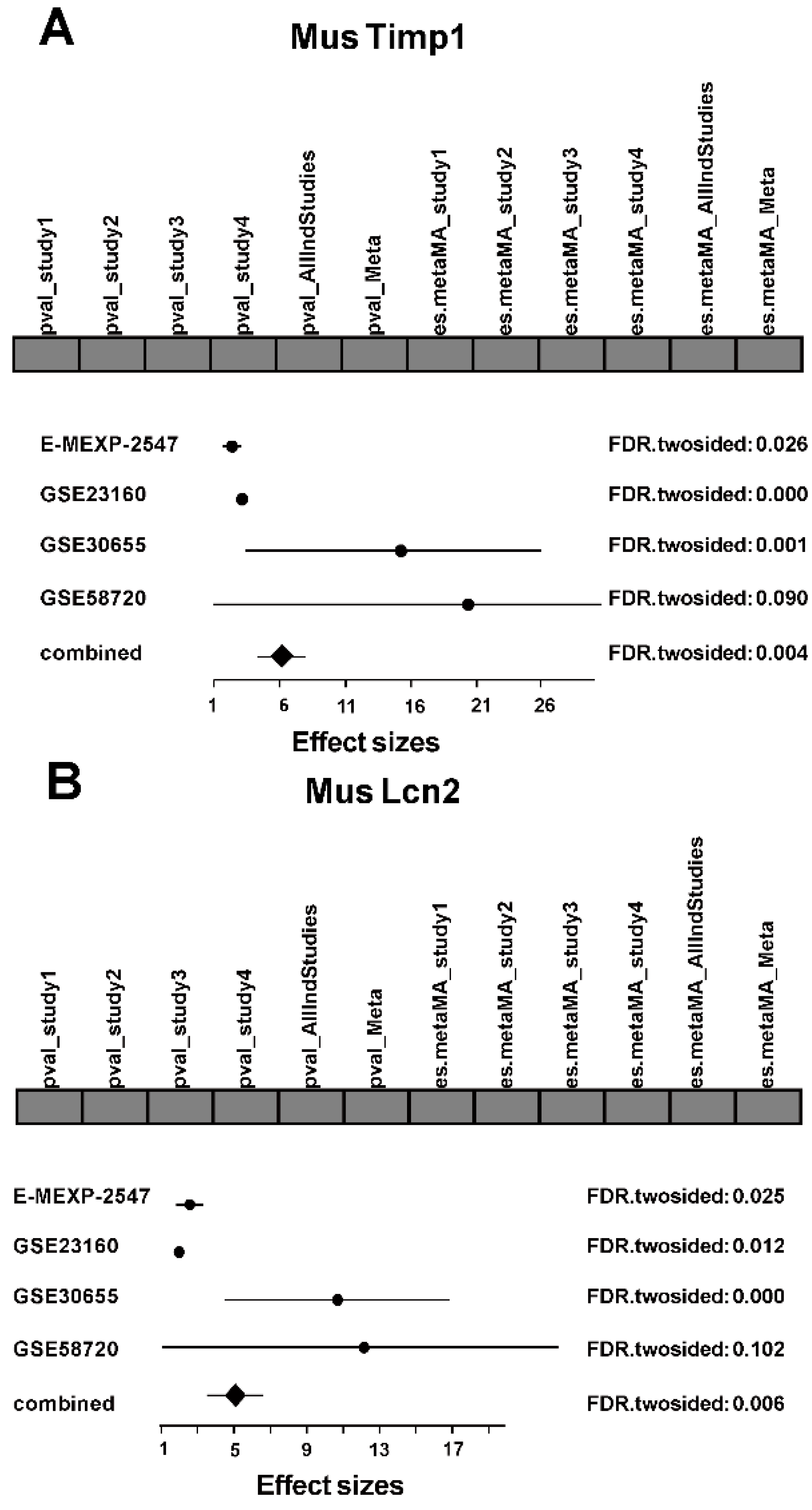
2.2. Meta-Analysis
2.2.1. Mus Function Annotation and Pathway Enrichment Analysis
2.2.2. Rat Function Annotation and Pathway Enrichment Analysis
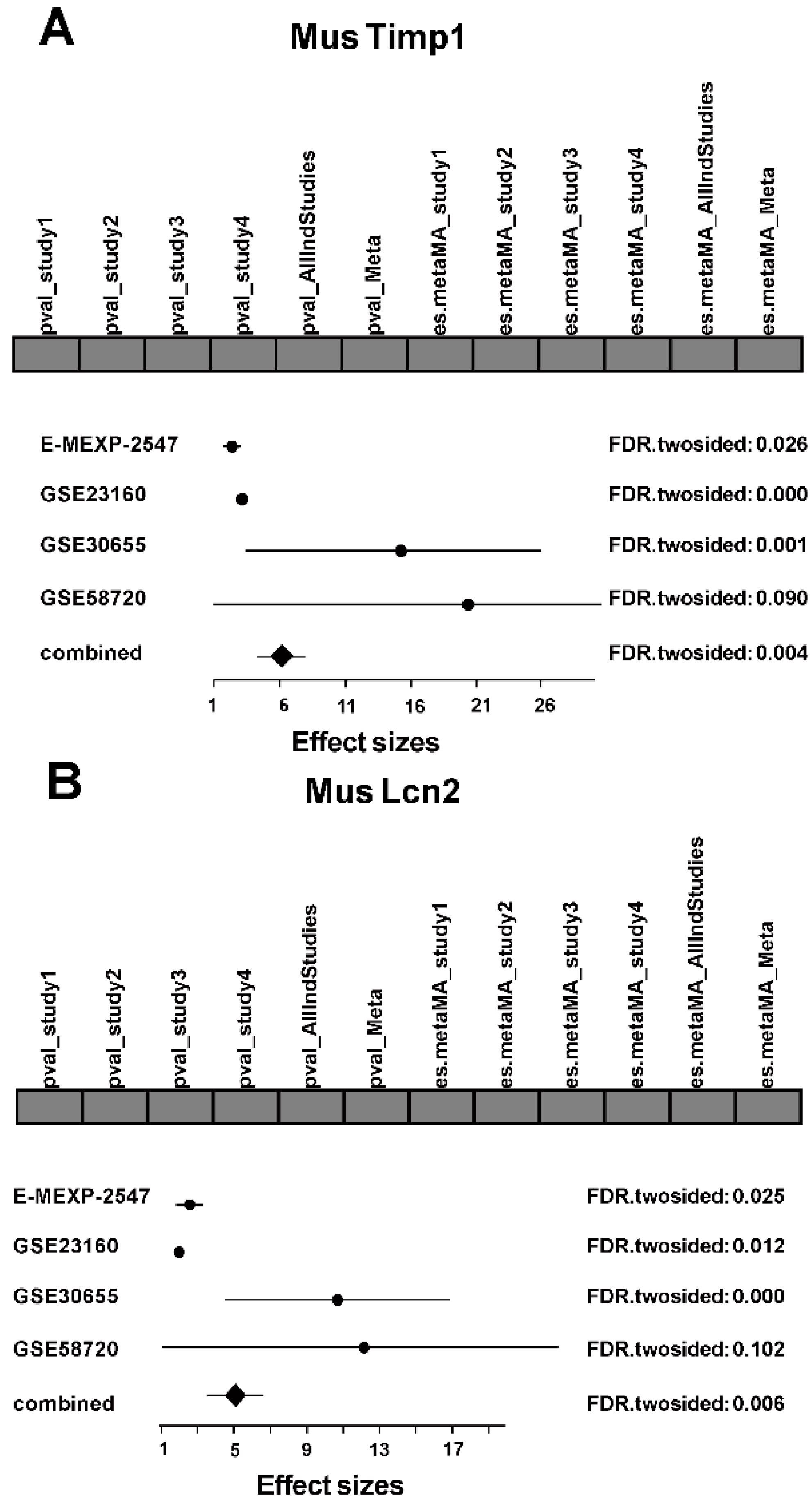
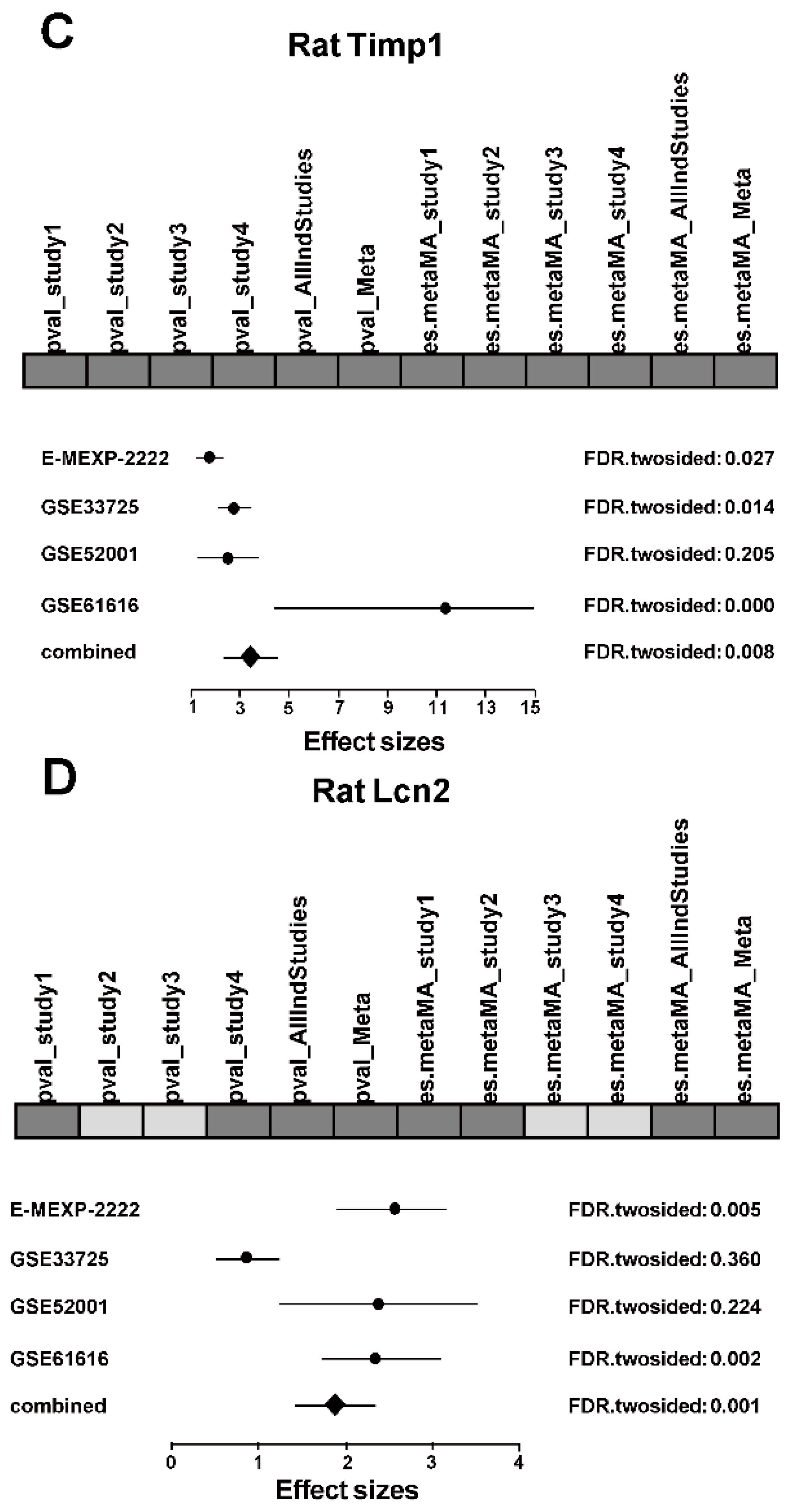
2.3. Differential Expression Analysis in Each Study
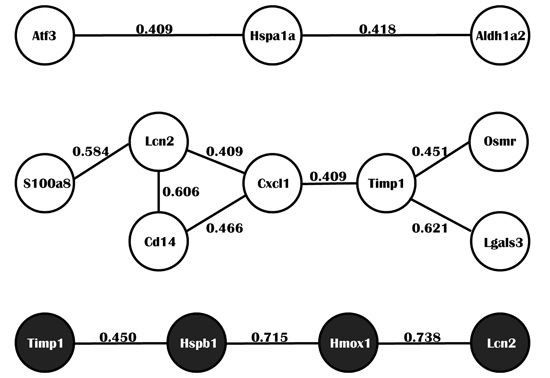
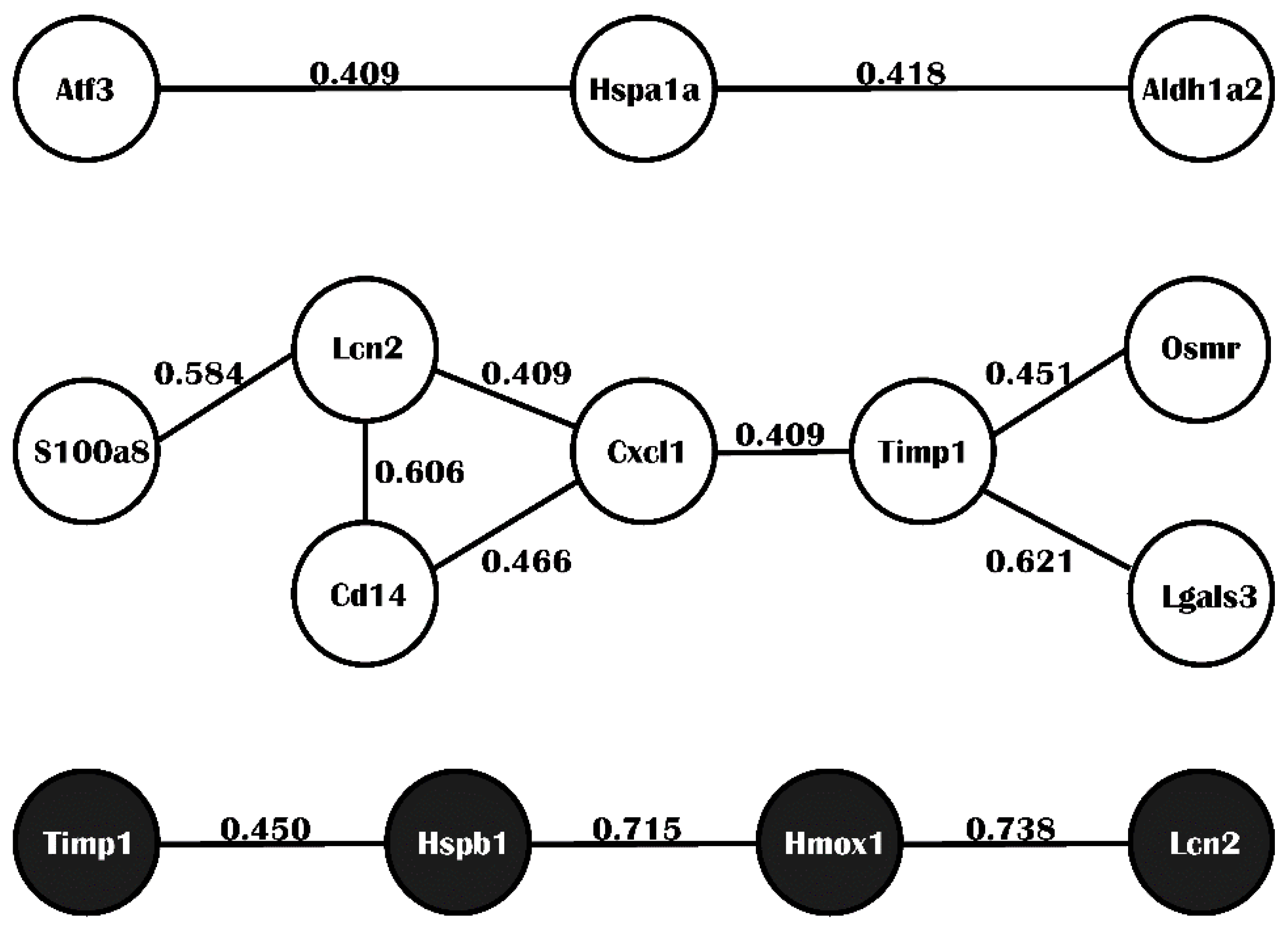
2.4. Integrated Analysis
3. Discussion
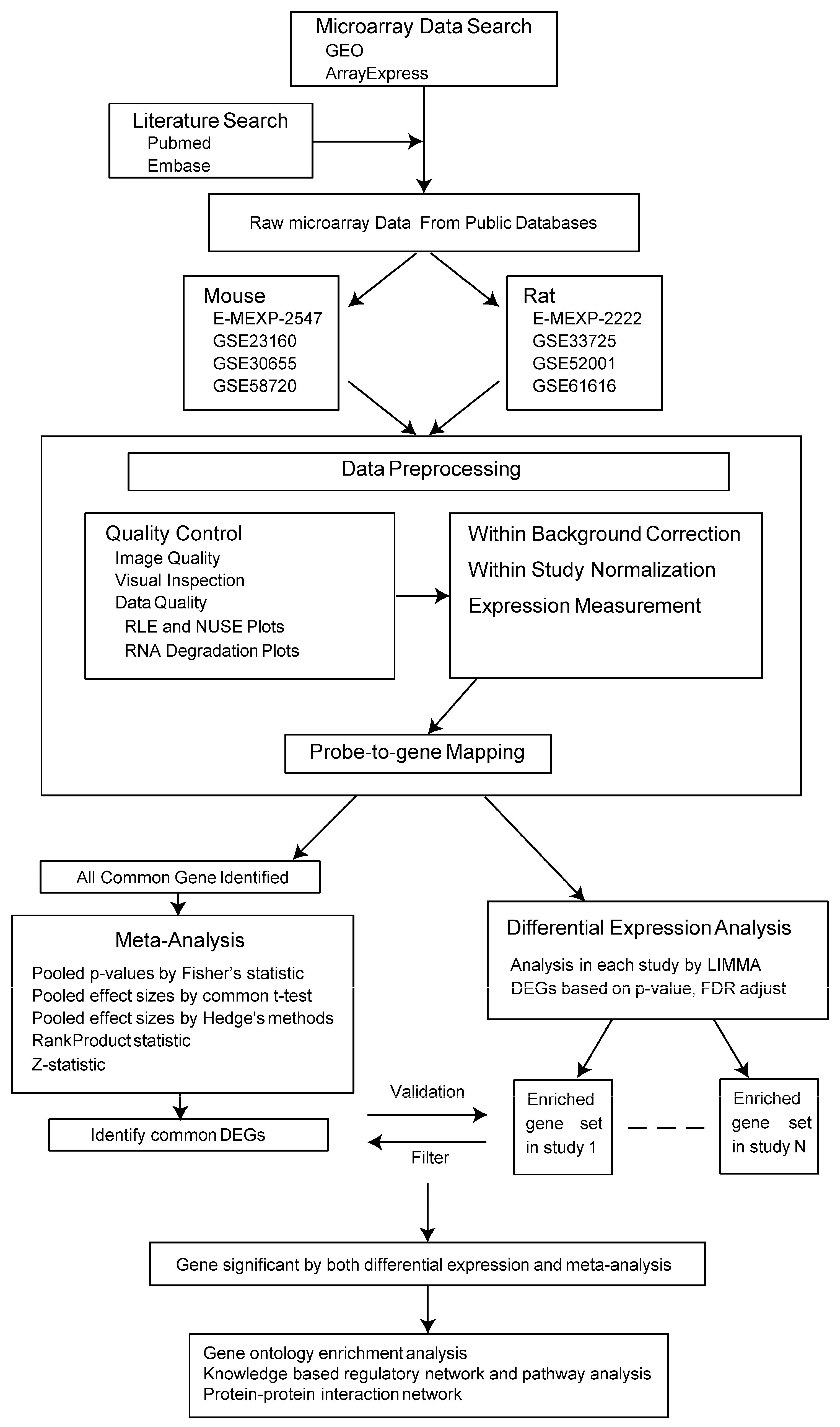
4. Materials and Methods
4.1. Microarray Data Search
4.2. Included and Excluded Criteria
4.3. Data Preprocessing
4.4. Meta-Analysis
4.5. Differential Expression Analysis in Each Study
4.6. Integrated Analysis
4.7. Functional Annotation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GO | Gene ontology |
| DEGs | Differentially expressed genes |
| ROS | Reactive oxygen species |
| CNS | Central neural system |
| RLE | Relative log expression |
| NUSE | Normalized unscaled standard errors |
| KNN | k-nearest neighbors |
| DAVID | Database for Annotation, Visualization and Integrated Discovery |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| PPI | Protein-protein interaction |
| NGAL | Neutrophil gelatinase-associated lipocalin |
| MCAO | Middle cerebral artery occlusion |
References
- Donnan, G.A.; Fisher, M.; Macleod, M.; Davis, S.M. Stroke. Lancet 2008, 371, 1612–1623. [Google Scholar] [CrossRef]
- Higgins, G.C.; Beart, P.M.; Shin, Y.S.; Chen, M.J.; Cheung, N.S.; Nagley, P. Oxidative stress: Emerging mitochondrial and cellular themes and variations in neuronal injury. J. Alzheimers Dis. 2010, 20, S453–S473. [Google Scholar] [PubMed]
- Ferrer, I.; Planas, A.M. Signaling of cell death and cell survival following focal cerebral ischemia: Life and death struggle in the penumbra. J. Neuropathol. Exp. Neurol. 2003, 62, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.Y.; Crack, P.J. Modulation of neuro-inflammation and vascular response by oxidative stress following cerebral ischemia-reperfusion injury. Curr. Med. Chem. 2008, 15, 1–14. [Google Scholar] [PubMed]
- Siddiqui, A.S.; Delaney, A.D.; Schnerch, A.; Griffith, O.L.; Jones, S.J.M.; Marra, M.A. Sequence biases in large scale gene expression profiling data. Nucleic Acids Res. 2006, 34. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, A.; Mondry, A.; Holmes, C.C.; Altman, D.G. Key issues in conducting a meta-analysis of gene expression microarray datasets. PLoS Med. 2008, 5, e184. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, J.; Julia, F.; Thallinger, G.G.; McFarlane, R.J.; Larcombe, L.D. Microarray meta-analysis: From data to expression to biological relationships. In Computational Medicine; Springer-Vienna: New York, NY, USA, 2012; pp. 59–77. [Google Scholar]
- Gertz, K.; Kronenberg, G.; Kälin, R.E.; Baldinger, T.; Werner, C.; Balkaya, M.; Eom, G.D.; Hellmann-Regen, J.; Kröber, J.; Miller, K.R. Essential role of interleukin-6 in post-stroke angiogenesis. Brain 2012, 135, 1964–1980. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Wong, C.H.; Peng, Z.F.; Manikandan, J.; Melendez, A.J.; Tan, T.M.; Crack, P.J.; Cheung, N.S. A global transcriptomic view of the multifaceted role of glutathione peroxidase-1 in cerebral ischemic—Reperfusion injury. Free Radic. Biol. Med. 2011, 50, 736–748. [Google Scholar] [CrossRef] [PubMed]
- White, R.E.; Palm, C.; Xu, L.; Ling, E.; Ginsburg, M.; Bernie, J.D.; Han, R.; Patterson, A.; Altman, R.B.; Giffard, R.G. Mice lacking the β2 adrenergic receptor have a unique genetic profile before and after focal brain ischaemia. ASN Neuro 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Jiang, Y.; Zhu, J.; Wen, Z.; Xu, X.; Xu, X.; Xie, Y.; Yang, L.; Xu, L.; Lan, W.; et al. Orosomucoid1: Involved in vascular endothelial growth factor-induced blood-brain barrier leakage after ischemic stroke in mouse. Brain Res. Bull. 2014, 109, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Szydlowska, K.; Gozdz, A.; Dabrowski, M.; Zawadzka, M.; Kaminska, B. Prolonged activation of ERK triggers glutamate-induced apoptosis of astrocytes: Neuroprotective effect of FK506. J. Neurochem. 2010, 113, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Mengozzi, M.; Cervellini, I.; Villa, P.; Erbayraktar, Z.; Gökmen, N.; Yilmaz, O.; Erbayraktar, S.; Manohasandra, M.; van Hummelen, P.; Vandenabeele, P. Erythropoietin-induced changes in brain gene expression reveal induction of synaptic plasticity genes in experimental stroke. Proc. Natl. Acad. Sci. USA 2012, 109, 9617–9622. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.; Zheng, Z.; Zhang, X.; Wei, Y.; Chu, K.; Brown, J.; Hong, G.; Chen, L. Salidroside-mediated neuroprotection is associated with induction of early growth response genes (EGRS) across a wide therapeutic window. Neurotox. Res. 2015, 28, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, Y.; Yang, J.; Zhao, X.; Li, Z. Dissecting Xuesaitong’s mechanisms on preventing stroke based on the microarray and connectivity map. Mol. BioSyst. 2015, 11, 3033–3039. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2012, 41, D808–D815. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Shuaib, A. Involvement of inflammatory cytokines in central nervous system injury. Prog. Neurobiol. 2002, 67, 161–172. [Google Scholar] [CrossRef]
- Roy Choudhury, G.; Ryou, M.-G.; Poteet, E.; Wen, Y.; He, R.; Sun, F.; Yuan, F.; Jin, K.; Yang, S.-H. Involvement of p38 MAPK in reactive astrogliosis induced by ischemic stroke. Brain Res. 2014, 1551, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Wang, Y.-C.; Bai, Y.-Y.; Lu, C.-Q.; Xu, T.-T.; Zhu, L.; Ju, S. Role of P38 MAPK on MMP activity in photothrombotic stroke mice as measured using an ultrafast MMP activatable probe. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J.; Brew, B.J. Microglia, macrophages, perivascular macrophages, and pericytes: A review of function and identification. J. Leukoc. Biol. 2004, 75, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Waje-Andreassen, U.; Krakenes, J.; Ulvestad, E.; Thomassen, L.; K.-M, M.; Aarseth, J.; Vedeler, C.A. IL-6: An early marker for outcome in acute ischemic stroke. Acta Neurol. Scand. 2005, 111, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Graeber, M.B.; Streit, W.J. Microglia: Biology and pathology. Acta Neuropathol. 2009, 119, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Arac, A.; Grimbaldeston, M.A.; Nepomuceno, A.R.B.; Olayiwola, O.; Pereira, M.P.; Nishiyama, Y.; Tsykin, A.; Goodall, G.J.; Schlecht, U.; Vogel, H.; et al. Evidence that meningeal mast cells can worsen stroke pathology in mice. Am. J. Pathol. 2014, 184, 2493–2504. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Lian, Y.-J.; Ma, Y.-Q.; Xie, N.-C.; Wu, C.-J. Elevated serum levels of CXC chemokine ligand-12 are associated with unfavorable functional outcome and mortality at 6-month follow-up in Chinese patients with acute ischemic stroke. Mol. Neurobiol. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.F.d.C.; Coelho, F.M.; Rodrigues, D.H.; Campos, A.C.; Barcelos, L.d.S.; Teixeira, M.M.; Rachid, M.A.; Teixeira, A.L. Blockade of CXCR1/2 chemokine receptors protects against brain damage in ischemic stroke in mice. Clinics 2013, 68, 391–394. [Google Scholar] [CrossRef]
- Connell, B.J.; Gordon, J.R.; Saleh, T.M. ELR-CXC chemokine antagonism is neuroprotective in a rat model of ischemic stroke. Neurosci. Lett. 2015, 606, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.; Ghorpade, A. Tissue inhibitor of metalloproteinase (TIMP)-1: The TIMPed balance of matrix metalloproteinases in the central nervous system. J. Neurosci. Res. 2003, 74, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Barone, F.C.; White, R.F.; Feuerstein, G.Z. Subtractive cloning identifies tissue inhibitor of matrix metalloproteinase-1 (TIMP-1) increased gene expression following focal stroke. Stroke 1998, 29, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Vukasovic, I.; Tesija-Kuna, A.; Topic, E.; Supanc, V.; Demarin, V.; Petrovcic, M. Matrix metalloproteinases and their inhibitors in different acute stroke subtypes. Clin. Chem. Lab. Med. 2006, 44, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Kurzepa, J.; Szczepanska-Szerej, A.; Stryjecka-Zimmer, M.; Malecka-Massalska, T.; Stelmasiak, Z. Simvastatin could prevent increase of the serum MMP-9/TIMP-1 ratio in acute ischaemic stroke. Folia Biol. 2006, 52, 181–183. [Google Scholar]
- Thang, N.M.; Kumasawa, K.; Tsutsui, T.; Nakamura, H.; Masaki, H.; Ono, T.; Kimura, T. Overexpression of endogenous TIMP-2 increases the proliferation of BeWo choriocarcinoma cells through the MAPK-signaling pathway. Reprod. Sci. 2013, 20, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Polimeni, M.; Valente, E.; Ulliers, D.; Opdenakker, G.; van den Steen, P.E.; Giribaldi, G.; Prato, M. Natural haemozoin induces expression and release of human monocyte tissue inhibitor of metalloproteinase-1. PLoS ONE 2013, 8, e71468. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, L.; Johnsen, A.H.; Sengeløv, H.; Borregaard, N. Isolation and primary structure of NGAL, a novel protein associated with human neutrophil gelatinase. J. Biol. Chem. 1993, 268, 10425–10432. [Google Scholar] [PubMed]
- Yang, J.; Goetz, D.; Li, J.Y.; Wang, W.; Mori, K.; Setlik, D.; Du, T.; Erdjument-Bromage, H.; Tempst, P.; Strong, R.; et al. An iron delivery pathway mediated by a lipocalin. Mol. Cell 2002, 10, 1045–1056. [Google Scholar] [CrossRef]
- Hasegawa, M.; Ishii, J.; Kitagawa, F.; Takahashi, K.; Hayashi, H.; Koide, S.; Tomita, M.; Takahashi, H.; Ozaki, Y.; Yuzawa, Y. Urinary neutrophil gelatinase-associated lipocalin as a predictor of cardiovascular events in patients with chronic kidney disease. Heart Vessels 2015, 30, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Weng, Y.-C.; Han, X.; Whaley, J.D.; McCrae, K.R.; Chou, W.-H. Lipocalin-2 released in response to cerebral ischaemia mediates reperfusion injury in mice. J. Cell. Mol. Med. 2015, 19, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Kim, J.-H.; Jang, E.; Lee, Y.M.; Soo Han, H.; Woo, D.K.; Park, D.H.; Kook, H.; Suk, K. Lipocalin-2 deficiency attenuates neuroinflammation and brain injury after transient middle cerebral artery occlusion in mice. J. Cereb. Blood Flow Metab. 2014, 34, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Haskill, S.; Peace, A.; Morris, J.; Sporn, S.A.; Anisowicz, A.; Lee, S.W.; Smith, T.; Martin, G.; Ralph, P.; Sager, R. Identification of three related human GRO genes encoding cytokine functions. Proc. Natl. Acad. Sci. USA 1990, 87, 7732–7736. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Grotendorst, G.R. Cloning and sequencing of a new gro transcript from activated human monocytes: Expression in leukocytes and wound tissue. Mol. Cell. Biol. 1990, 10, 5596–5599. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, C.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. High- and low-affinity binding of GROα and neutrophil-activating peptide 2 to interleukin 8 receptors on human neutrophils. Proc. Natl. Acad. Sci. USA 1992, 89, 10542–10546. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-H.; Frost, E.; To, V.; Robinson, S.; Ffrench-Constant, C.; Geertman, R.; Ransohoff, R.M.; Miller, R.H. The chemokine receptor CXCR2 controls positioning of oligodendrocyte precursors in developing spinal cord by arresting their migration. Cell 2002, 110, 373–383. [Google Scholar] [CrossRef]
- Losy, J.; Zaremba, J.; Skrobański, P. CXCL1 (GRO-α) chemokine in acute ischaemic stroke patients. Folia Neuropathol. 2005, 43, 97–102. [Google Scholar] [PubMed]
- Yang, Z.; Chen, Y.; Fu, Y.; Yang, Y.; Zhang, Y.; Chen, Y.; Li, D. Meta-analysis of differentially expressed genes in osteosarcoma based on gene expression data. BMC Med. Genet. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Gentleman, R.; Carey, V.; Huber, W.; Irizarry, R.; Dudoit, S. Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer Science & Business Media: New York, NY, USA, 2006; p. 494. [Google Scholar]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affy—Analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43. [Google Scholar] [CrossRef] [PubMed]
- Troyanskaya, O.; Cantor, M.; Sherlock, G.; Brown, P.; Hastie, T.; Tibshirani, R.; Botstein, D.; Altman, R.B. Missing value estimation methods for DNA microarrays. Bioinformatics 2001, 17, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Marot, G.; Foulley, J.-L.; Mayer, C.-D.; Jaffrézic, F. Moderated effect size and P-value combinations for microarray meta-analyses. Bioinformatics 2009, 25, 2692–2699. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.K.; Yu, U.; Kim, S.; Yoo, O.J. Combining multiple microarray studies and modeling interstudy variation. Bioinformatics 2003, 19, i84–i90. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Michael, A.; Ball, C.A.; Blake, J.A.; David, B.; Heather, B.; Michael Cherry, J.; Davis, A.P.; Kara, D.; Dwight, S.S.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Series Dataset | Ischemic Period (min) | Reperfusion Period (h) | No. of Samples | Platform | |
|---|---|---|---|---|---|
| MCAO | Sham | ||||
| Mus musculus | |||||
| E-MEXP-2547 [8] | 30 | 24, 240 | 5 | 5 | Affymetrix GeneChip Mouse Gene 1.0 ST Array |
| GSE23160 [9] | 120 | 2, 8, 24 | 24 | 8 | Illumina MouseRef-8 v2.0 expression beadchip |
| GSE30655 [10] | 60 | 24 | 7 | 3 | Affymetrix Mouse Genome 430 2.0 Array |
| GSE58720 [11] | 90 | 24 | 3 | 3 | Agilent-028005 SurePrint G3 Mouse GE 8 × 60 K Microarray |
| Rattus norvegicus | |||||
| E-MEXP-2222 [12] | 90 | 6, 24 | 12 | 3 | Affymetrix Rat Genome 230 2.0 Array |
| GSE33725 [13] | NA | 2, 6 | 6 | 6 | Agilent-014879 Whole Rat Genome Microarray 4 × 44 K G4131F |
| GSE52001 [14] | 120 | 144 | 3 | 3 | Agilent-028282 Whole Rat Genome Microarray 4 × 44 K v3 |
| GSE61616 [15] | 120 | 168 | 5 | 5 | Affymetrix Rat Genome 230 2.0 Array |
| GO ID | GO Term | % | p-Value |
|---|---|---|---|
| Biological Process | |||
| GO:0002237 | response to molecule of bacterial origin | 1.67 | 2.10 × 10−3 |
| GO:0032655 | regulation of interleukin-12 production | 1.11 | 2.46 × 10−3 |
| GO:0043086 | negative regulation of catalytic activity | 2.22 | 2.78 × 10−3 |
| GO:0010033 | response to organic substance | 5.56 | 2.92 × 10−3 |
| GO:0044092 | negative regulation of molecular | 2.50 | 3.28 × 10−3 |
| Cellular Component | |||
| GO:0044445 | cytosolic part | 1.67 | 5.75 × 10−3 |
| GO:0031974 | membrane-enclosed lumen | 8.89 | 2.27 × 10−2 |
| GO:0005829 | cytosol | 5.00 | 2.33 × 10−2 |
| GO:0005886 | plasma membrane | 18.61 | 2.59 × 10−2 |
| GO:0005938 | cell cortex | 1.94 | 2.78 × 10−2 |
| Molecular Function | |||
| GO:0005509 | calcium ion binding | 7.50 | 1.03 × 10−2 |
| GO:0019838 | growth factor binding | 1.67 | 1.21 × 10−2 |
| GO:0043167 | ion binding | 25.28 | 2.49 × 10−2 |
| GO:0046872 | metal ion binding | 24.72 | 2.75 × 10−2 |
| GO:0003677 | DNA binding | 12.78 | 2.92 × 10−2 |
| ID | Term | % | p-Value |
|---|---|---|---|
| Mus musculus | |||
| mmu04010 | MAPK signaling pathway | 4.72 | 8.14 × 10−5 |
| mmu04060 | Cytokine-cytokine receptor interaction | 3.06 | 2.65 × 10−2 |
| mmu04062 | Chemokine signaling pathway | 2.50 | 3.18 × 10−2 |
| mmu04722 | Neurotrophin signaling pathway | 1.94 | 4.90 × 10−2 |
| mmu05410 | Hypertrophic cardiomyopathy (HCM) | 1.39 | 9.16 × 10−2 |
| Rattus norvegicus | |||
| rno04730 | Long-term depression | 2.77 | 1.44 × 10−6 |
| rno04010 | MAPK signaling pathway | 5.14 | 1.52 × 10−6 |
| rno04070 | Phosphatidylinositol signaling system | 2.37 | 1.82 × 10−6 |
| rno04540 | Gap junction | 2.37 | 2.29 × 10−6 |
| rno04670 | Leukocyte transendothelial migration | 2.77 | 3.29 × 10−6 |
| GO ID | GO Term | % | p-Value |
|---|---|---|---|
| Biological Process | |||
| GO:0009611 | response to wounding | 9.49 | 1.91 × 10−7 |
| GO:0022604 | regulation of cell morphogenesis | 4.74 | 5.44 × 10−6 |
| GO:0006954 | inflammatory response | 5.53 | 3.57 × 10−5 |
| GO:0006813 | potassium ion transport | 4.35 | 7.65 × 10−5 |
| GO:0006811 | ion transport | 9.88 | 1.16 × 10−4 |
| Cellular Component | |||
| GO:0044456 | synapse part | 6.72 | 1.44 × 10−6 |
| GO:0005886 | plasma membrane | 24.90 | 1.52 × 10−6 |
| GO:0043005 | neuron projection | 8.70 | 1.82 × 10−6 |
| GO:0005856 | cytoskeleton | 13.44 | 2.29 × 10−6 |
| GO:0045202 | synapse | 7.91 | 3.29 × 10−6 |
| Molecular Function | |||
| GO:0005509 | calcium ion binding | 9.49 | 5.33 × 10−5 |
| GO:0005216 | ion channel activity | 6.32 | 6.73 × 10−5 |
| GO:0043167 | ion binding | 25.30 | 8.37 × 10−5 |
| GO:0022838 | substrate specific channel activity | 6.32 | 9.35 × 10−5 |
| GO:0022836 | gated channel activity | 5.53 | 9.48 × 10−5 |
| Study | Count | Up-Regulated | Down-Regulated |
|---|---|---|---|
| Mus musculus | |||
| E-MEXP-2547 | 179 | 179 | 0 |
| GSE23160 | 22 | 22 | 0 |
| GSE30655 | 341 | 128 | 213 |
| GSE58720 | 1162 | 841 | 321 |
| Rattus norvegicus | |||
| E-MEXP-2222 | 83 | 81 | 2 |
| GSE33725 | 38 | 38 | 0 |
| GSE52001 | 94 | 66 | 28 |
| GSE61616 | 827 | 695 | 132 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Qiu, Z.; Gao, S.; Chen, Q.; Li, S.; Tan, W.; Liu, X.; Wang, Z. Integrated Analysis of Expression Profile Based on Differentially Expressed Genes in Middle Cerebral Artery Occlusion Animal Models. Int. J. Mol. Sci. 2016, 17, 776. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050776
Zhou H, Qiu Z, Gao S, Chen Q, Li S, Tan W, Liu X, Wang Z. Integrated Analysis of Expression Profile Based on Differentially Expressed Genes in Middle Cerebral Artery Occlusion Animal Models. International Journal of Molecular Sciences. 2016; 17(5):776. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050776
Chicago/Turabian StyleZhou, Huaqiang, Zeting Qiu, Shaowei Gao, Qinchang Chen, Si Li, Wulin Tan, Xiaochen Liu, and Zhongxing Wang. 2016. "Integrated Analysis of Expression Profile Based on Differentially Expressed Genes in Middle Cerebral Artery Occlusion Animal Models" International Journal of Molecular Sciences 17, no. 5: 776. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050776