
MicroRNA-24 Attenuates Neointimal Hyperplasia in the Diabetic Rat Carotid Artery Injury Model by Inhibiting Wnt4 Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
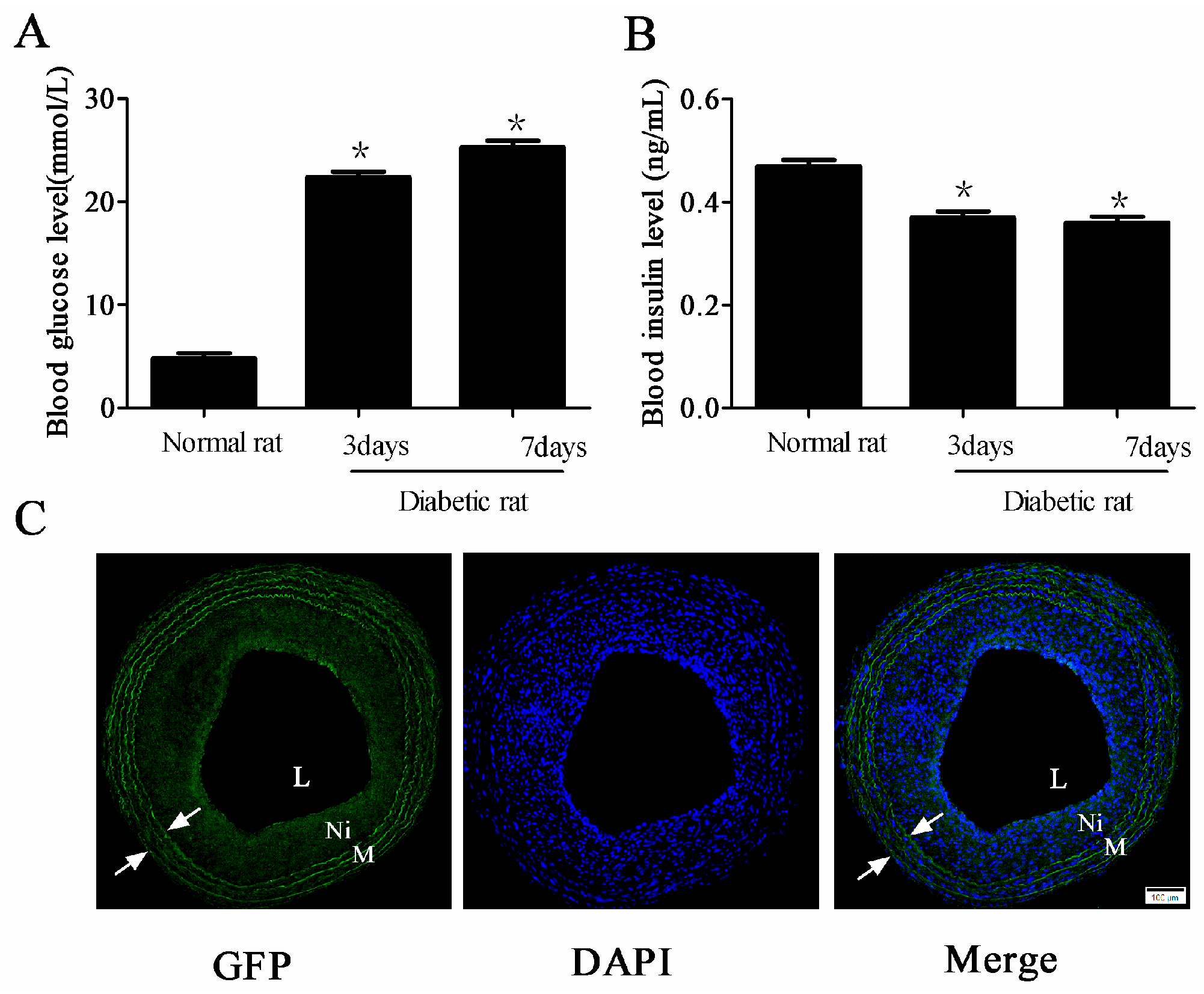
2.1. Adenovirus Was Transfected into the Carotid Artery in Diabetic Rats Successfully
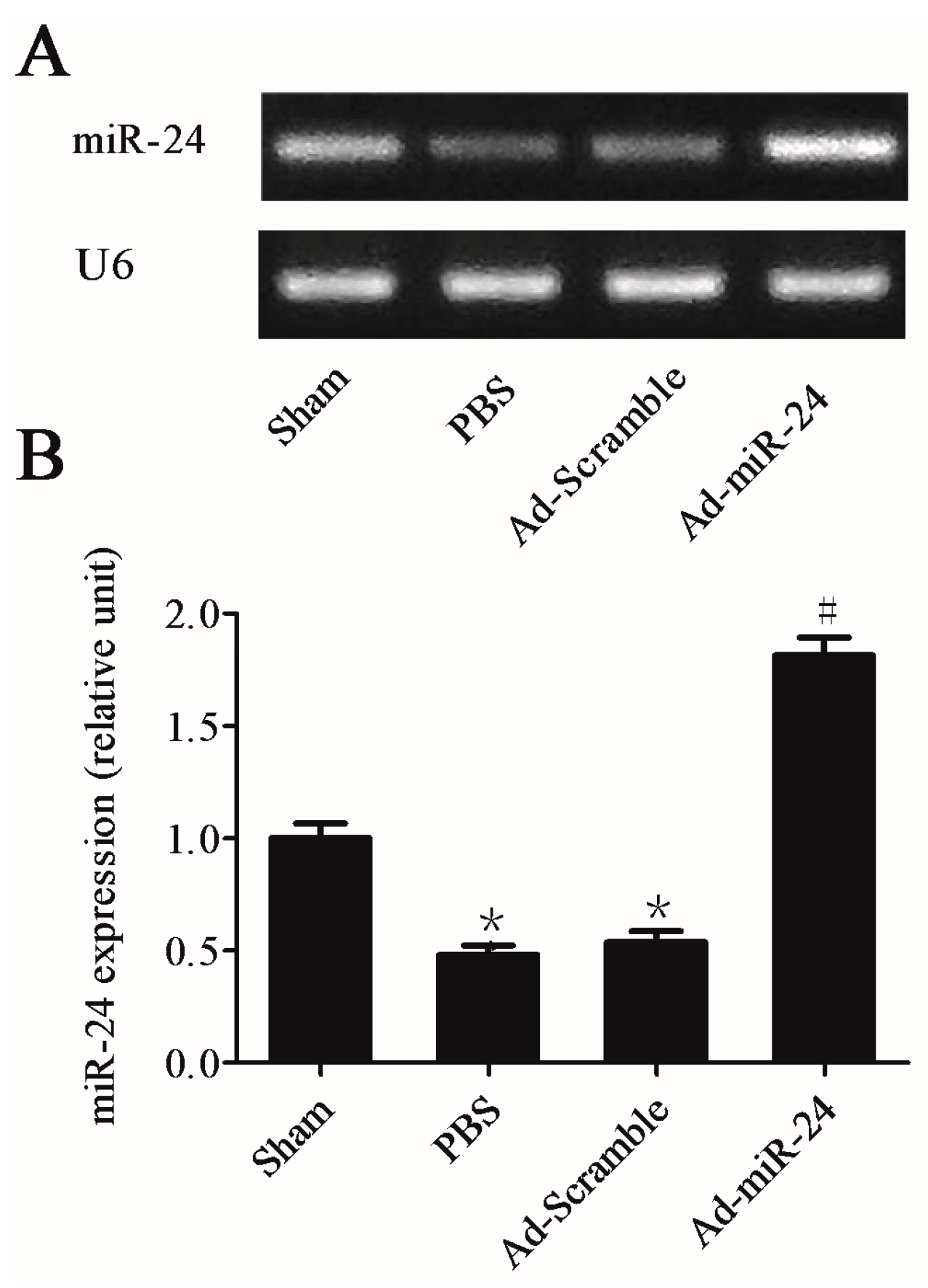
2.2. miR-24 Expression Was Decreased in Carotid Artery after Balloon Injury in Diabetic Rats
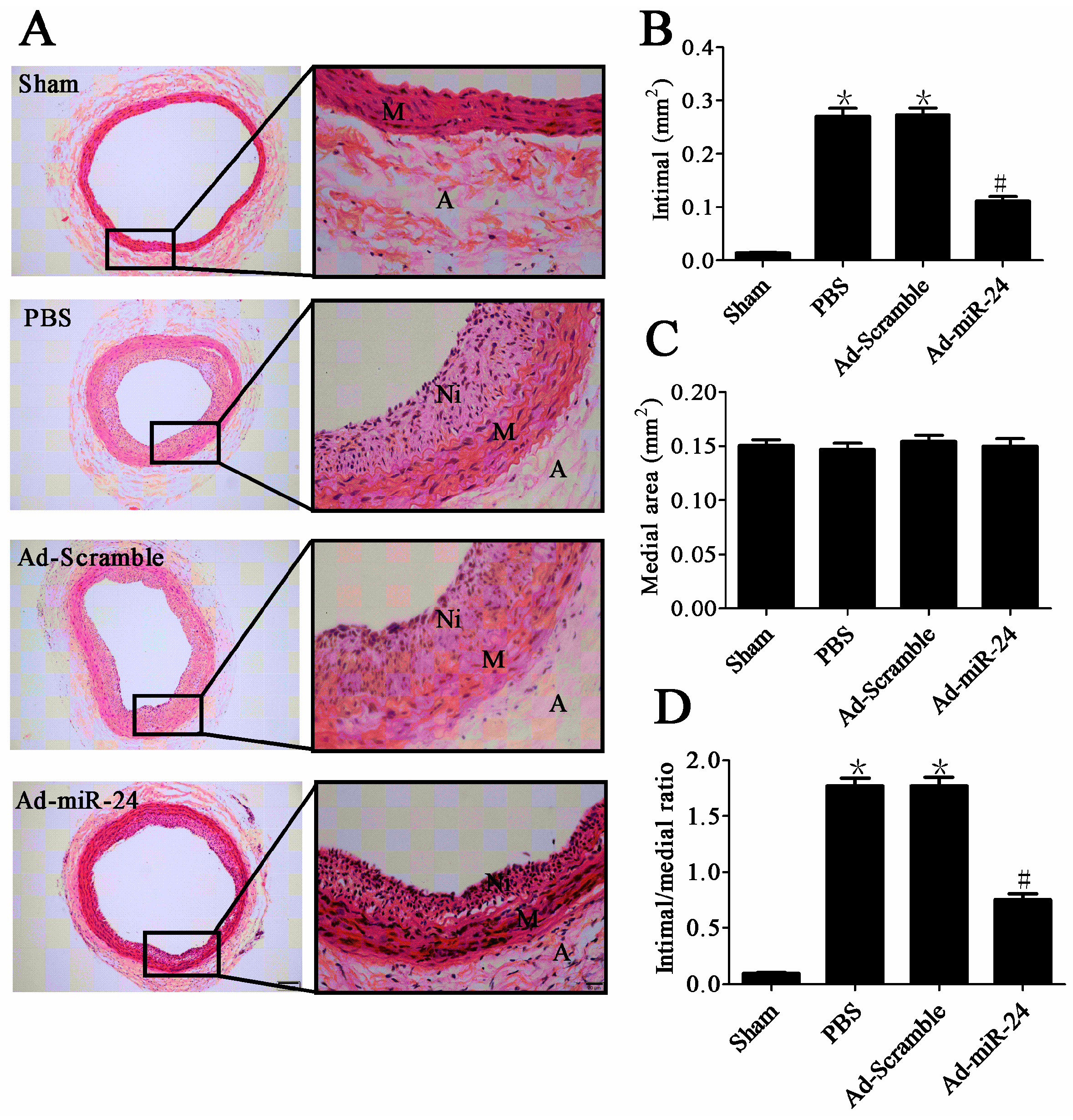
2.3. Over-Expression of miR-24 Suppressed Neointimal Hyperplasia in Diabetic Rats
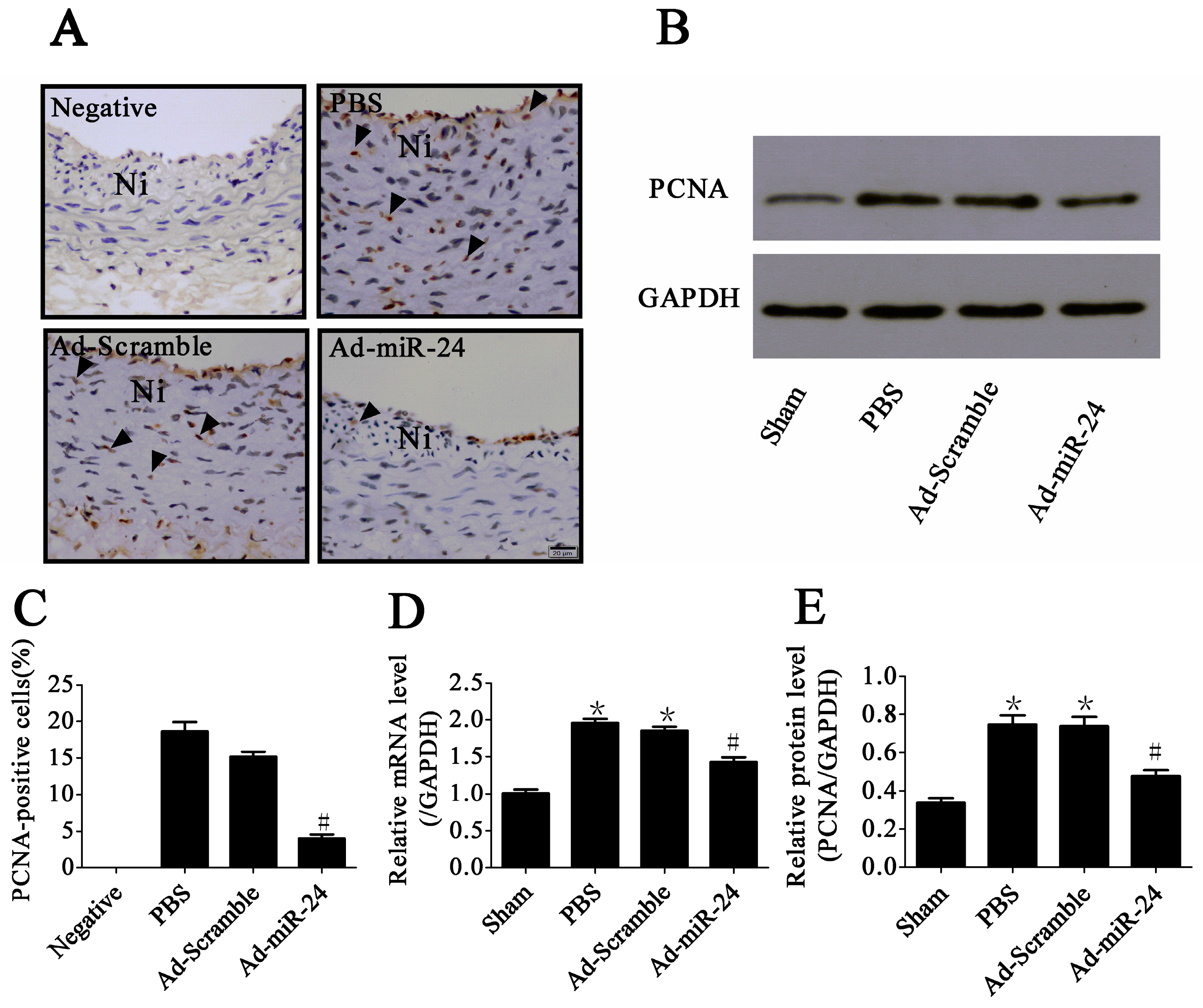
2.4. Over-Expression of miR-24 Inhibited VSMC Proliferation in Diabetic Rats
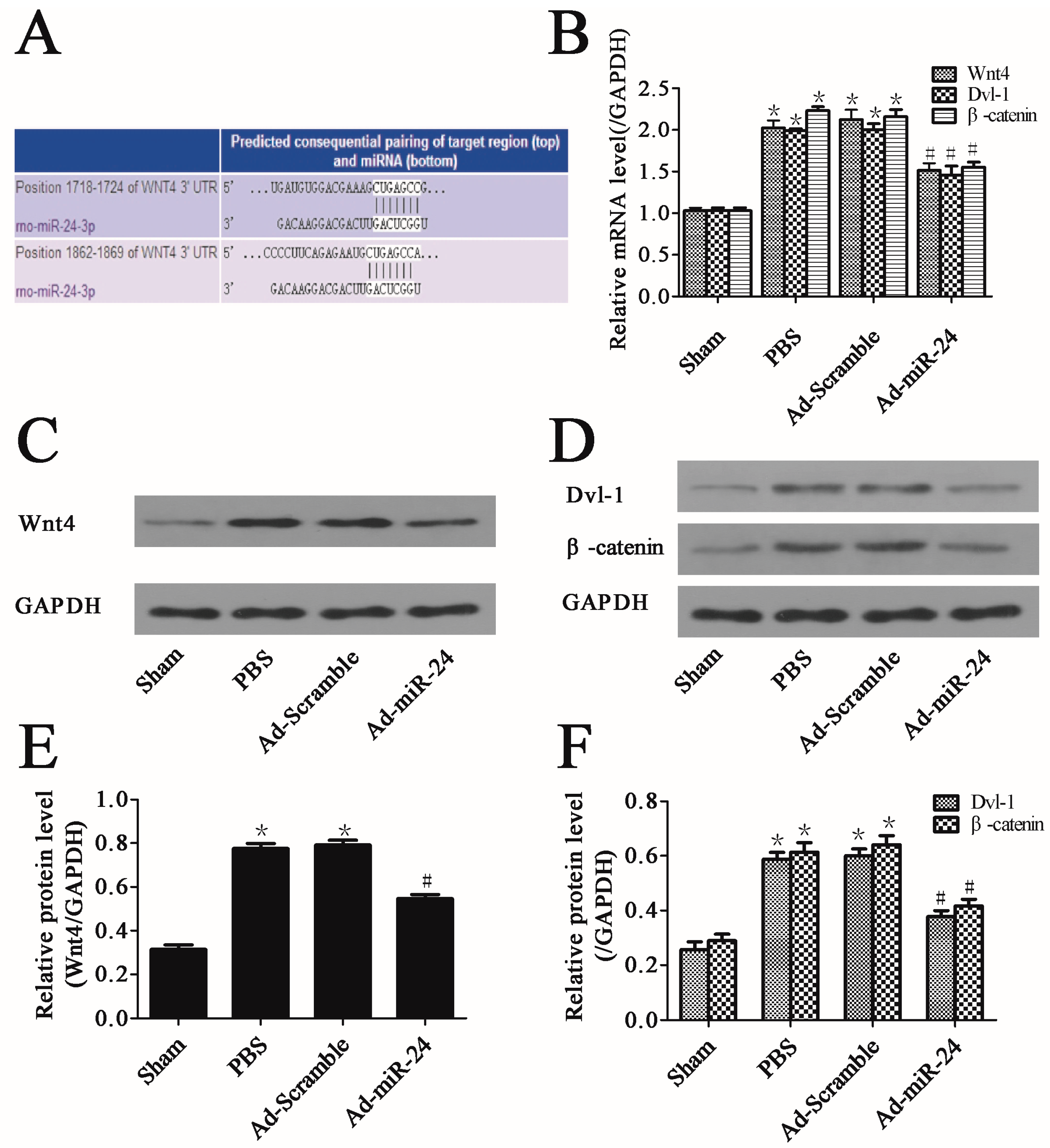
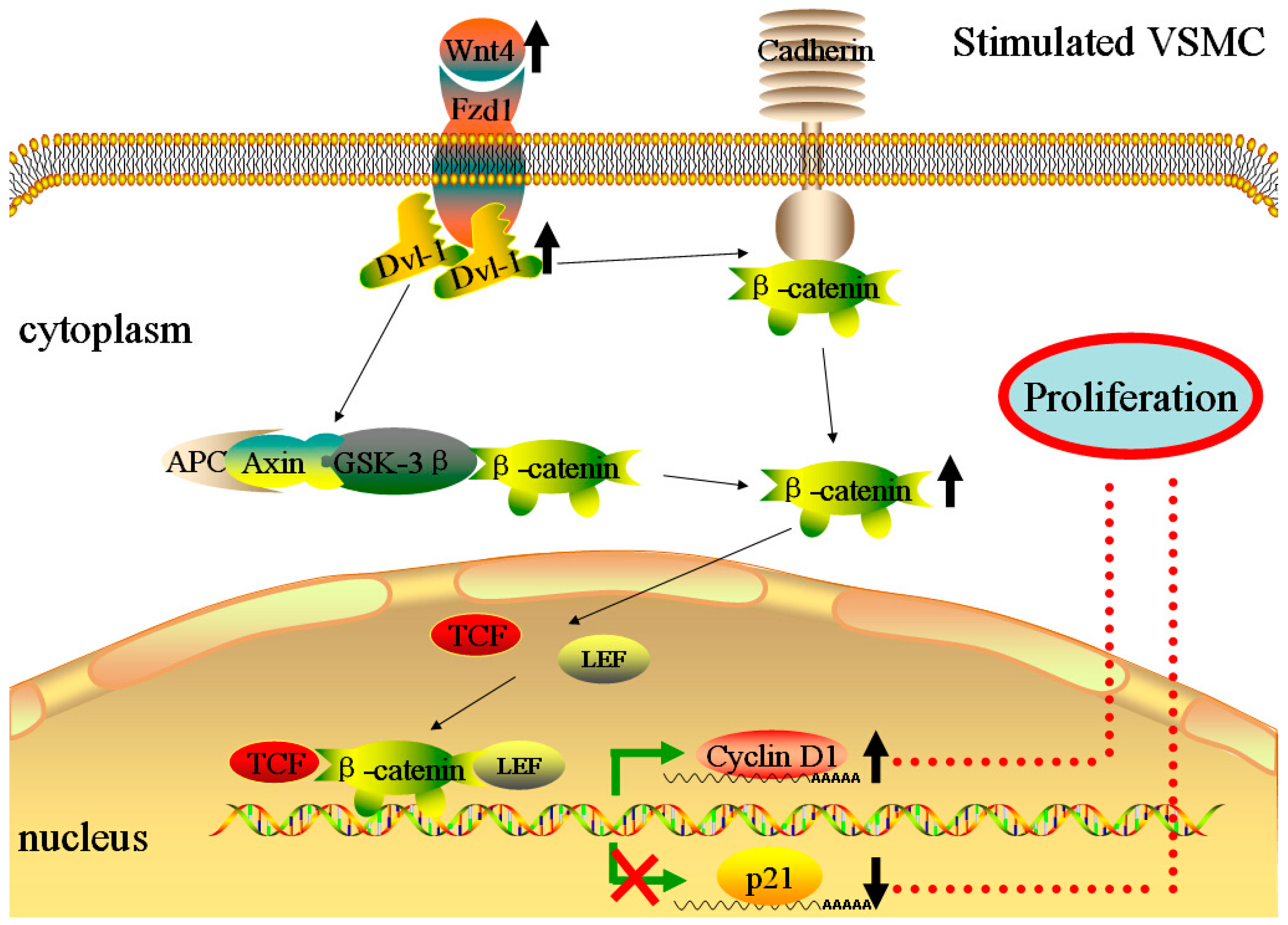
2.5. Over-Expression of miR-24 Suppressed the Expression of Wnt4 Signaling Pathway
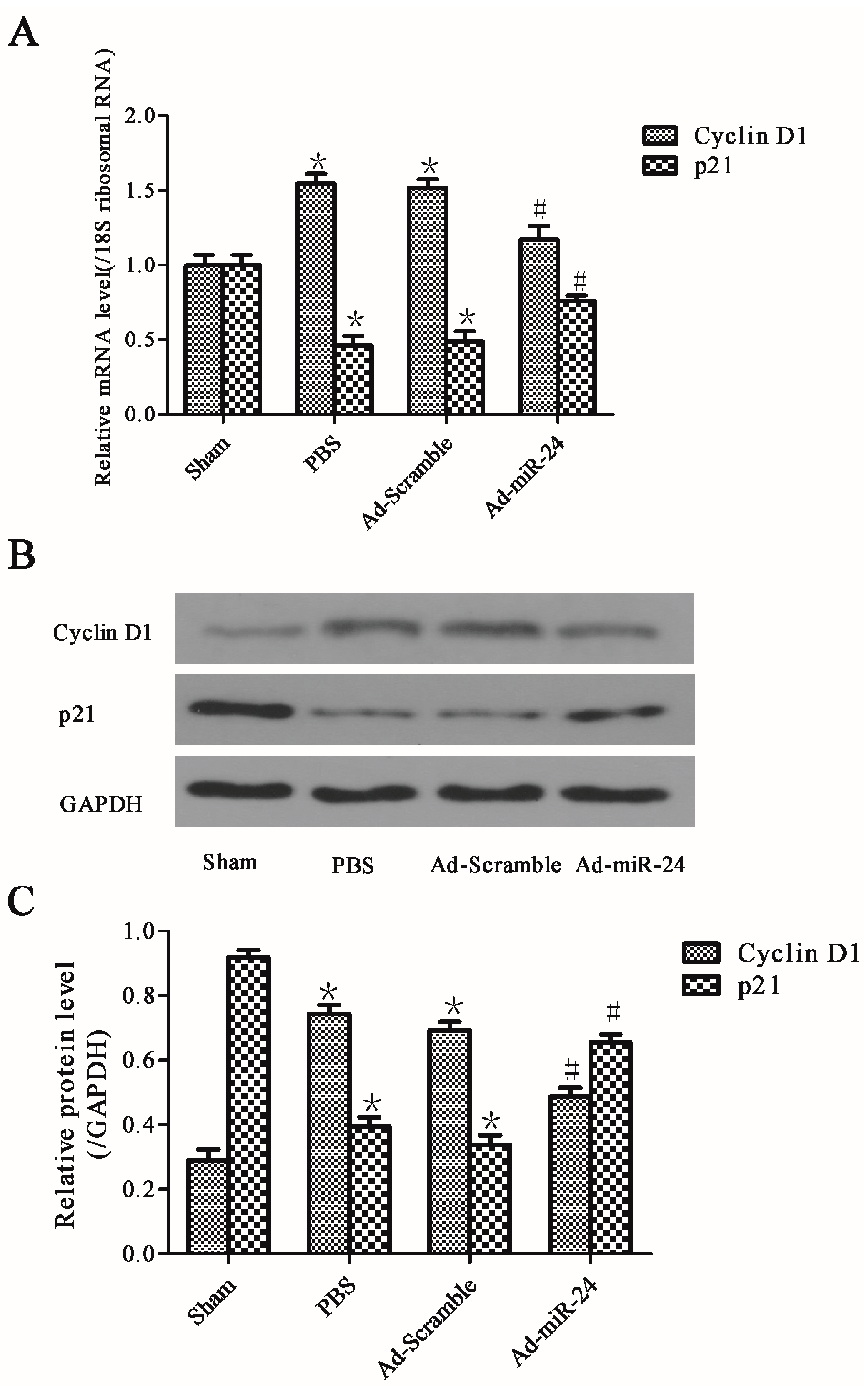
2.6. Over-Expression of miR-24 Altered the Expression of Cell Cycle-Associated Molecules
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Streptozotocin (STZ)-Induced Diabetic Rats
4.3. Construction of miR-24 Expression Adenoviral Vector
4.4. Diabetic Rat Carotid Artery Balloon Injury Model and Adenovirus Transduction
4.5. Assessment of Adenovirus Transfection Efficiency
4.6. Histologic Examination
4.7. Assessment of VSMC Proliferation
4.8. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.9. Western Blotting Analysis
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gomes, M.B. Glitazones and the metabolic syndrome: Mechanism of action, pathophysiology and therapeutic indications. Arq. Bras. Endocrinol. Metabol. 2006, 50, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Yang, C.; Yang, J. A hypothesis: Glucagon-like peptide-1 may play a protective role in diabetic cardiomyopathy. Int. J. Cardiol. 2015, 197, 294–295. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, L.; He, J.; Bi, Y.; Li, M.; Wang, T. Prevalence and control of diabetes in Chinese adults. JAMA 2013, 310, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Lastra, G.; Manrique, C. Perivascular adipose tissue, inflammation and insulin resistance: Link to vascular dysfunction and cardiovascular disease. Horm. Mol. Biol. Clin. Investig. 2015, 22, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jiang, H.; Chen, S.S.; Chen, J.; Li, W.Q.; Xu, S.K. Lentivirus-mediated RNAi targeting CREB binding protein attenuatesneointimal formation and promotes re-endothelialization in balloon injured rat carotid artery. Cell. Physiol. Biochem. 2010, 26, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Tezuka, D.; Morishita, R.; Isobe, M. An initial case of suppressed restenosis with nuclear factor-κB decoy transfection after percutaneous coronary intervention. J. Genet. Med. 2009, 11, 89–91. [Google Scholar]
- Hao, H.; Gabbiani, G.; Bochaton-Piallat, M.L. Arterial smooth muscle cell heterogeneity: Implications for atherosclerosis and restenosis development. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, J.; Xu, C.; Yang, J.; Guo, Q.; Hu, Q. Resveratrol inhibits phenotypic switching of neointimal vascular smooth muscle cells after balloon injury through blockade of Notch pathway. J. Cardiovasc. Pharmacol. 2014, 63, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Benter, I.F.; Sarkhou, F.; Al-Khaldi, A.T.; Chandrasekhar, B.; Attur, S.; Dhaunsi, G.S. The dual targeting of EGFR and ErbB2 with the inhibitor Lapatinib corrects high glucose-induced apoptosis and vascular dysfunction by opposing multiple diabetes-induced signaling changes. J. Drug Target. 2015, 23, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Rawal, S.; Manning, P.; Katare, R. Cardiovascular microRNAs: As modulators and diagnostic biomarkers of diabetic heart disease. Cardiovasc. Diabetol. 2014, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.X.; Yang, J. Microribonucleic acids and vascular restenosis. Saudi. Med. J. 2014, 35, 796–801. [Google Scholar] [PubMed]
- Ji, R.; Cheng, Y.; Yue, J.; Yang, J.; Liu, X.; Chen, H. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of microRNA in vascular neointimal lesion formation. Circ. Res. 2007, 100, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Marchand, A.; Atassi, F.; Gaaya, A.; Leprince, P.; Le Feuvre, C.; Soubrier, F. The Wnt/β-catenin pathway is activated during advanced arterial aging in humans. Aging Cell. 2011, 10, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Tsaousi, A.; Williams, H.; Lyon, C.A.; Taylor, V.; Swain, A.; George, S.J. Wnt4/β-catenin signaling induces VSMC proliferation and is associated with intimal thickening. Circ. Res. 2011, 108, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Quasnichka, H.; Slater, S.C.; Beeching, C.A.; Boehm, M.; Sala-Newby, G.B.; George, S.J. Regulation of smooth muscle cell proliferation by β-catenin/T-cell factor signaling involves modulation of Cyclin D1 and p21 expression. Circ. Res. 2006, 99, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.Y.; He, Y.Z.; Xu, Y.; Jiang, X.H.; Ye, W.; Pan, Z.M. Emodin prevents intima thickness via Wnt4/Dvl-1/β-catenin signalingpathway mediated by miR-126 in balloon-injured carotid artery rats. Exp. Mol. Med. 2015, 47, e170. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Chen, Y.G. Dishevelled: The hub of wnt signaling. Cell Signal. 2010, 22, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Xu, Y.; He, Y.; Jiang, X.; Ye, W.; Pan, Z. Wnt4/β-catenin signaling pathway modulates balloon-injured carotid arteryrestenosis via disheveled-1. Int. J. Clin. Exp. Pathol. 2014, 7, 8421–8431. [Google Scholar] [PubMed]
- Lewis, B.P.; Shih, I.H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of Mammalian MicroRNA Targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Patti, G.; Nusca, A.; di Sciascio, G. Meta-analysis comparison (nine trials) of outcomes with drug-eluting stents versus bare metal stents in patients with diabetes mellitus. Am. J. Cardiol. 2008, 102, 1328–1334. [Google Scholar] [CrossRef] [PubMed]
- Salvi, A.; Abeni, E.; Portolani, N.; Barlati, S.; de Petro, G. Human hepatocellular carcinoma cell-specific miRNAs reveal the differential expression of miR-24 and miR-27a in cirrhotic/non-cirrhotic HCC. Int. J. Oncol. 2013, 42, 391–402. [Google Scholar] [PubMed]
- Chan, M.C.; Hilyard, A.C.; Wu, C.; Davis, B.N.; Hill, N.S.; Lal, A. Molecular basis for antagonism between PDGF and the TGFβ family of signalling pathways by control of miR-24 expression. EMBO J. 2010, 29, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xiao, Y.; Mou, Y.; Zhao, Y.; Blankesteijn, W.M.; Hall, J.L. A role for the β-catenin/T-cell factor signaling cascade in vascular remodeling. Circ. Res. 2002, 90, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Grandy, D.; Han, J.; Zhang, X.; Rao, S.; Akunuru, S.; Li, H. Discovery and characterization of a small molecule inhibitor of the PDZ domain of dishevelled. J. Biol. Chem. 2009, 284, 16256–16263. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, J.B.; Habas, R. The developmental biology of dishevelled: An enigmatic protein governing cell fate and cell polarity. Development 2005, 132, 4421–4436. [Google Scholar] [CrossRef] [PubMed]
- Vlad-Fiegen, A.; Langerak, A.; Eberth, S.; Müller, O. The Wnt pathway destabilizes adherens junctions and promotes cell migration via β-catenin and its target gene cyclin D1. FEBS Open Bio 2012, 2, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Kamei, J.; Toyofuku, T.; Hori, M. Negative regulation of p21 by β-catenin/TCF signaling: A novel mechanism by which cell adhesion molecules regulate cell proliferation. Biochem. Biophys. Res. Commun. 2003, 312, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Bond, M.; Sala-Newby, G.B.; Wu, Y.J.; Newby, A.C. Biphasic effect of p21Cip1 on smooth muscle cell proliferation: Role of PI 3-kinase and Skp2-mediated degradation. Cardiovasc. Res. 2006, 69, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Fan, Z.; Yang, C.; Yang, J. MicroRNA-facilitated gene delivery:A potential therapeutic approach for vascularrestenosis without influencing endothelial function. Int. J. Cardiol. 2015, 201, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Wronska, A.; Uryu, K.; Diacovo, T.G.; Gao, M.; Marx, S.O. A selective microRNA-based strategy inhibits restenosis while preserving endothelial function. J. Clin. Investig. 2014, 124, 4102–4114. [Google Scholar] [CrossRef] [PubMed]
- Lyon, A.R.; Sato, M.; Hajjar, R.J.; Samulski, R.J.; Harding, S.E. Gene therapy: Targeting the myocardium. Heart 2008, 94, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Mason, D.; Chen, Y.Z.; Krishnan, H.V.; Sant, S. Cardiac gene therapy: Recent advances and future directions. J. Control. Release 2015, 215, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.T.; Zhen, J.; Pang, B.; Gu, J.N.; Wu, S.S. Ginsenoside Rg1 ameliorates oxidative stress and myocardial apoptosis in streptozotocin-induced diabetic rats. J. Zhejiang Univ. Sci. B 2015, 16, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yang, J.; Bai, J.; Pu, P.; Liu, J.; Wang, F. Suv39h1 protects from myocardial ischemia-reperfusion injury in diabetic rats. Cell. Physiol. Biochem. 2014, 33, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, L.; Yang, J.; Ding, J.; Li, S.; Wu, H. MicroRNA-22 targeting CBP protects against myocardial ischemia-reperfusion injury through anti-apoptosis in rats. Mol. Biol. Rep. 2014, 41, 555–561. [Google Scholar] [CrossRef] [PubMed]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Fan, Z.; Yang, J.; Ding, J.; Yang, C.; Chen, L. MicroRNA-24 Attenuates Neointimal Hyperplasia in the Diabetic Rat Carotid Artery Injury Model by Inhibiting Wnt4 Signaling Pathway. Int. J. Mol. Sci. 2016, 17, 765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060765
Yang J, Fan Z, Yang J, Ding J, Yang C, Chen L. MicroRNA-24 Attenuates Neointimal Hyperplasia in the Diabetic Rat Carotid Artery Injury Model by Inhibiting Wnt4 Signaling Pathway. International Journal of Molecular Sciences. 2016; 17(6):765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060765
Chicago/Turabian StyleYang, Jian, Zhixing Fan, Jun Yang, Jiawang Ding, Chaojun Yang, and Lihua Chen. 2016. "MicroRNA-24 Attenuates Neointimal Hyperplasia in the Diabetic Rat Carotid Artery Injury Model by Inhibiting Wnt4 Signaling Pathway" International Journal of Molecular Sciences 17, no. 6: 765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060765