1. Introduction
Humanity living in developed countries is experiencing an increase in life expectancy; however, this positive outcome seems to be at the cost of a greater incidence of lifestyle- and age-associated diseases. These include cardiovascular diseases (CVDs), cancer and amyloid pathologies, both systemic (e.g., type 2 diabetes, T2DM) and neurodegenerative (e.g., Alzheimer’s (AD) and Parkinson (PD) diseases). Most of these pathologies are particularly hard to treat due to their slow progression, possibly spanning several decades, and the appearance of their clinical signs at mid- or old age, when cell loss is already conspicuous and irreversible. It is then evident that, in the absence of early reliable diagnostic tools and effective therapies, prevention is still the best strategy to combat these pathological conditions.
It is not surprising that the shift of researchers’ attention from “cure” to “prevention” has gradually led to an extension of the focus of their search, by adding “food”, and hence “diet”, to “drugs”. Several epidemiological and observational studies support the belief that traditional alimentary regimens such as the Mediterranean (MD) and Asian diets are associated with improved ageing and a reduced incidence of age-associated diseases, including CVDs, cancer and cognitive decline [
1]. The new design of the MD pyramid, proposed by the Mediterranean Diet Foundation Expert Group [
2], emphasizes the importance, in addition to caloric restriction (CR), of frugality, conviviality, physical activity and adequate rest; it also confirms the importance of the plant-based core (vegetables, fruits, legumes, grains, nuts and seeds) and, in particular, of extra virgin olive oil (EVOO) as the main lipid source. Moreover, a key feature of the MD is the high intake of phytonutrients (notably vitamins and natural phenols) that, by themselves, can induce multiple signalling pathways involved in protein homeostasis, DNA repair, metabolism regulation and antioxidant defences that recall a caloric restriction regime [
3,
4].
A key feature of natural phenolics is their remarkable antioxidant power. The latter has been associated with many beneficial properties of plant polyphenols via modulation of oxidative pathways [
2], through direct action on enzymes, proteins, receptors and several types of signalling pathways [
3,
4] as well as by interfering with epigenetic modifications of chromatin [
5]. In particular, the beneficial effects of olive oil and olive leaf extracts were already known in the ancient world, and scientifically investigated since the last couple of centuries, leading to a focus on their biological properties, including the antioxidant, antimicrobial, hypoglycemic, vasodilator and antihypertensive effects, whose clinical significance was first reported in 1950 [
6]. Some of these properties have led to the inclusion in the European Pharmacopoeia (Ph. Eur.) of the 80% alcoholic extract of olive leaves [
7], containing oleuropein (OLE), hydroxytyrosol (HT), caffeic acid, tyrosol, apigenin and verbascoside [
8].
The increasing interest in natural polyphenols has produced a plethora of studies that have investigated their medical efficacy
in vitro, in cell cultures, in model organisms and, to a lesser extent, in humans, together with the biochemical and biological modifications underlying their effects. Plant polyphenols, or their molecular scaffolds, can also be the starting point in developing new drugs especially designed to combat chronic inflammatory states, atherosclerosis and the risk of thrombosis related to CVDs [
9], cancer [
10], amyloid deposition associated with AD and T2DM, and age-associated neurodegeneration [
1,
11].
Here we review the results of the studies on the polyphenols found in the olive tree and in the EVOO and the most recent advances towards their possible clinical use, mainly concerning neurodegenerative diseases, atherosclerosis, cancer, T2DM and the metabolic syndrome.
2. Olive Tree Polyphenols
Natural phenolic substances are secondary plant metabolites, a major group of plant compounds (over 8000) chemically characterized by the presence of one or more aromatic rings with one or more hydroxyl substituents [
1]. Plant polyphenols are elaborated as phytoalexins used to combat pests and bacterial infections. The olive tree (
Olea europaea) produces its own battery of polyphenols that includes flavonols, lignans and glycosides. The latter belong to the class of iridoids, a type of monoterpenes composed of a cyclopentane ring fused to a six-atom oxygen heterocycle; the molecules containing a broken cyclopentane ring are known as secoiridoids.
Olive tree polyphenols are found in the lipid and water (as minute droplets), fractions of olive oil, and include the phenolic alcohols, HT (3,4-dihydroxyphenylethanol, 3,4-DHPEA) and tyrosol (
p-hydroxyphenylethanol,
p-HPEA) and their secoiridoid precursors. These include the HT ester of elenolic acid (known as oleuropein, OLE), the main responsible for the bitter taste of olive leaves and drupes; the dialdehydic derivative of decarboxymethyl elenolic acid bound to either HT (3,4-dihydroxyphenylethanol-elenolic acid dialdehyde, 3,4-DHPEA-EDA, also known as oleacein) or to tyrosol (
p-hydroxyphenylethanol-elenolic acid dialdehyde,
p-HPEA-EDA, also known as oleocanthal). The latter is the main responsible for the burning sensation that occurs in the back of the throat when consuming EVOO [
12,
13]. Olive tree polyphenols also include verbascoside, the caffeoylrhamnosylglucoside of HT, a phenolic acid derivative, the lignans 1-acetoxypinoresinol and pinoresinol, and other secoiridoids.
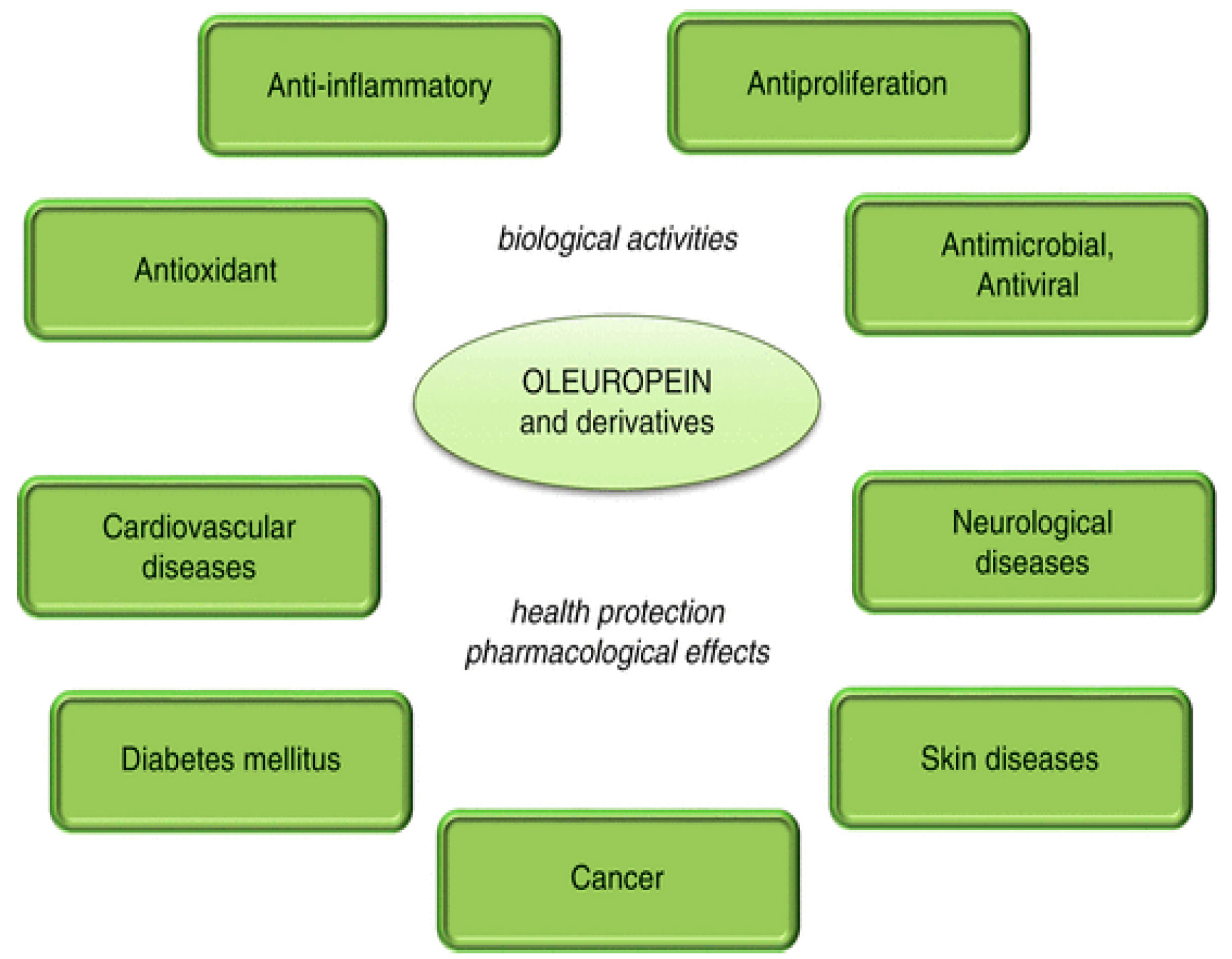
Olive tree polyphenols may be responsible for some of the properties of medical interest in this plant; these include anti-atherogenic, antihepatotoxic, hypoglycemic, anti-inflammatory, antitumor, antiviral and immunomodulator activities [
14,
15] that appear only in part related to the antioxidant power of these molecules. OLE, demethyloleuropein and ligstroside, together with their metabolic derivatives (elenolic acid, HT), are the most abundant phenolics in the EVOO [
16].
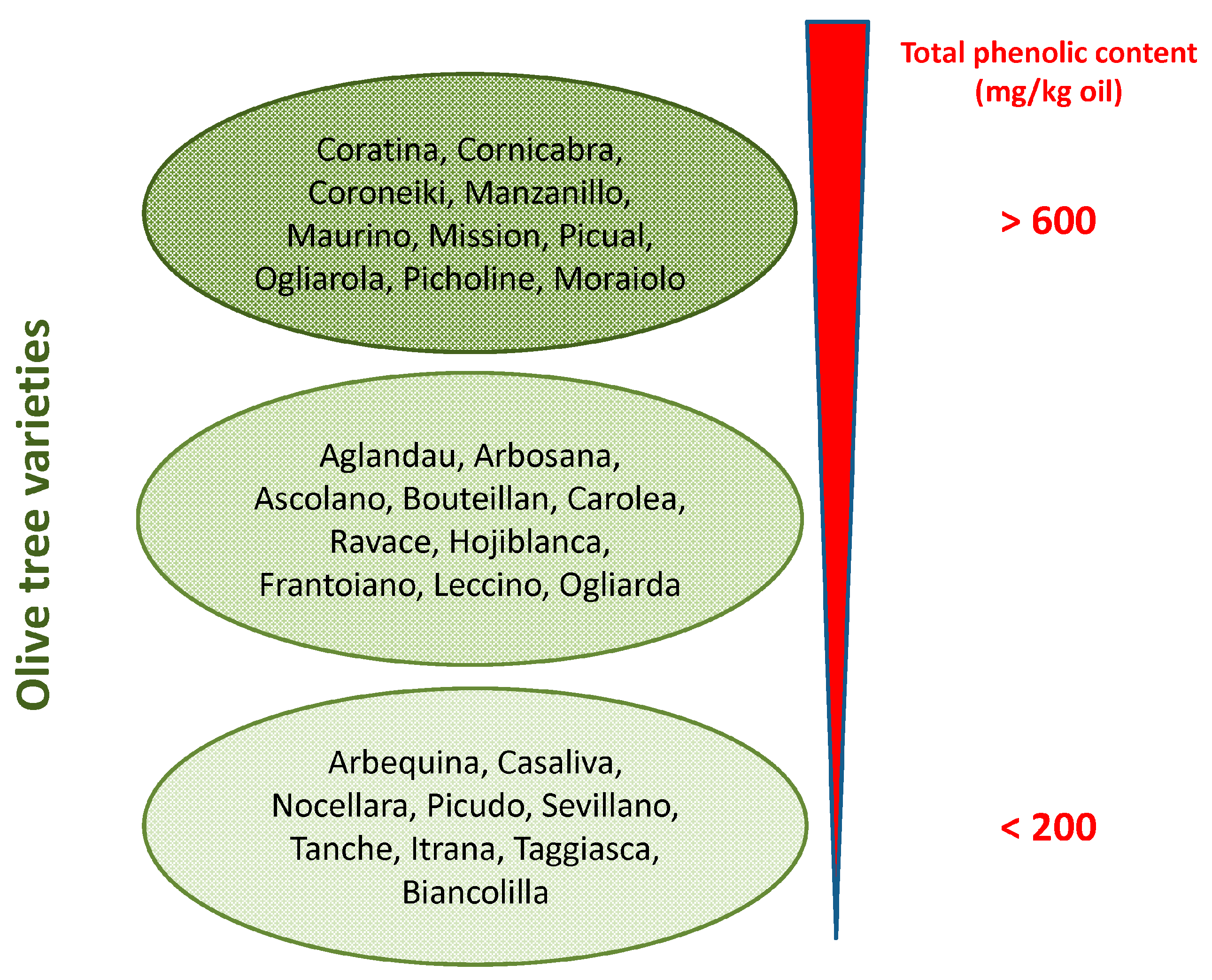
Phenolic concentration in EVOO depends on several variables such as (i) the olive cultivar (
Figure 1) and the ripening stage of fruit [
17]; (ii) environmental factors (altitude, cultivation practices, and amount of irrigation); (iii) extraction conditions (heating, added water and malaxation); (iv) extraction systems used to separate oil from olive pastes (pressure, centrifugation systems); and (v) storage conditions and time, due to spontaneous oxidation, and suspended particle deposition [
18]. At best, the content of OLE in EVOO can reach levels exceeding 60 mg/100 g (
Figure 1).
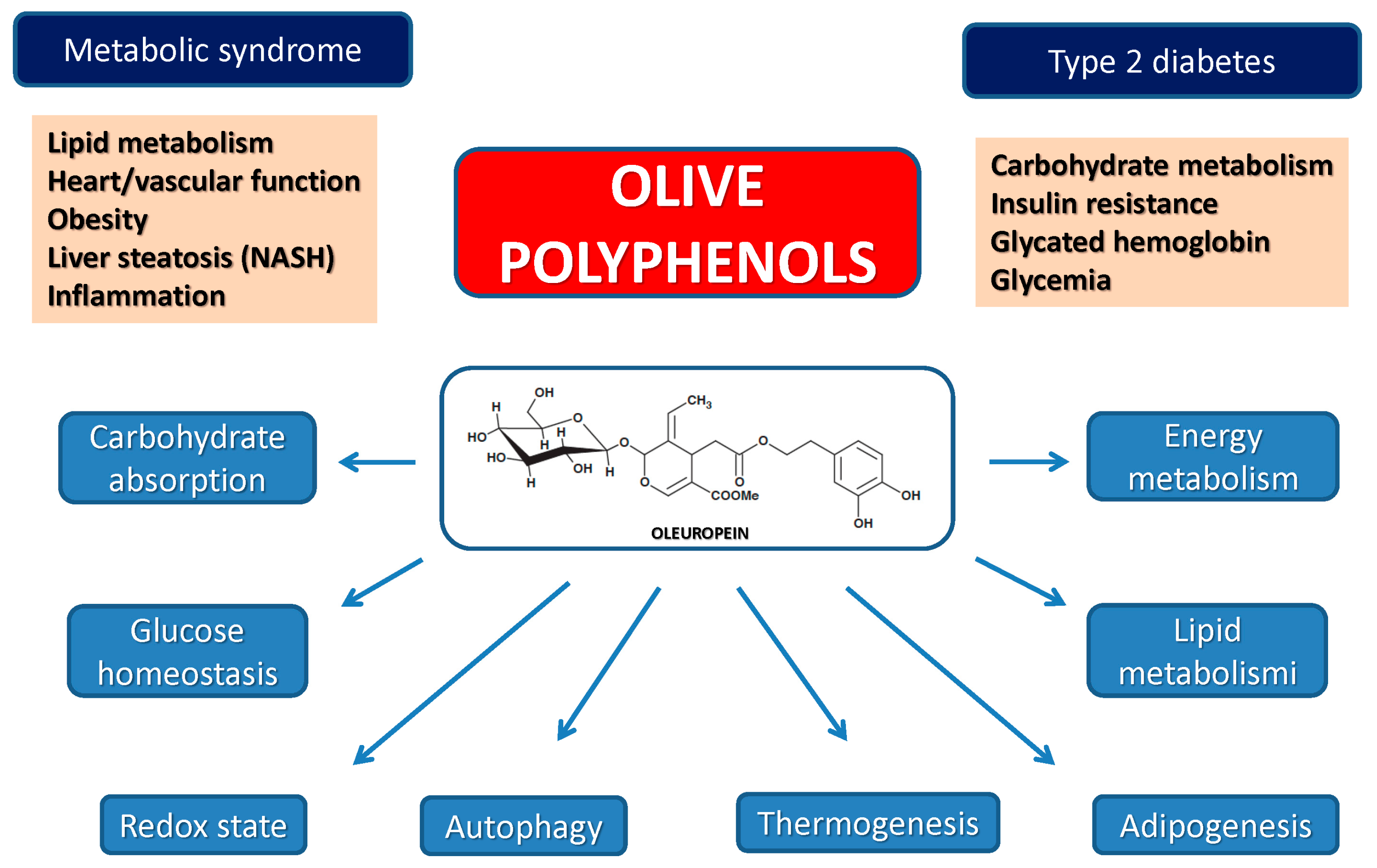
3. Biochemical Effects of Olive Polyphenols Considered as Caloric Restriction Mimickers
The high content in plant polyphenols, one of the main features underlying the beneficial effects of the MD, is provided mainly by the use of EVOO, the main source of alimentary lipids. The wide and increasingly recognized beneficial properties of plant polyphenols have led to proposing them as nutraceuticals and the aliments containing them as functional foods. The latter are defined as “Natural or processed foods that contain known or unknown biologically-active compounds; these foods, in defined, effective, and non-toxic amounts, provide a clinically proven and documented health benefit for the prevention, management, or treatment of chronic diseases” [
19]. Extensive research, clinical trials, epidemiological and observational studies have for a long time described the close association between the MD and the Asian diet regarding their polyphenols and a number of physiological and metabolic effects [
1,
20,
21,
22,
23]; the latter, most often, are similar to those associated with caloric restriction (CR) in humans [
24,
25], indicating that these substances are CR mimickers [
26] (
Table 1).
The effectiveness of CR to prolong lifespan and to reduce the risk of age-associated diseases is widely recognized [
25]. However, a CR regime can hardly be sustained for long periods of time; this is why diet integration with factors able to mimic the beneficial effects of a reduction of caloric intake can be highly appreciated. Plant polyphenols, including olive ones, induce CR-like effects in muscle, brain, fat tissue and kidney in several ways, particularly through the activation and increased levels of sirtuins (Sirt) [
26,
27].
Sirt are NAD-dependent type-3 deacetylases [
28] whose activity is modulated by the metabolic state of the cells and induced by CR. More contradictory data on CR-induced Sirt1 changes have been reported in liver, where decreased Sirt1 levels were found with an ensuing decrease of hepatic fat synthesis and accumulation [
29]. Sirt are involved in lifespan and metabolism regulation in varying organisms [
26,
29]. Among the Sirt family, Sirt1, the most investigated, protects the cells against oxidative stress and DNA damage. Many of the cellular effects of Sirt1 are mediated by gene regulation following its ability to control the acetylation/deacetylation state, and hence the activity, of several transcription factors, including p53, FOXOs, NFκB, Nrf2, PPARα/γ, PGC1α and LXR [
30,
31,
32,
33,
34,
35]. These factors are known to be involved in the control of apoptosis, autophagy, cell proliferation, oxidative stress, inflammation, protein synthesis, carbohydrate and lipid metabolism. Sirt induction by CR, resveratrol or other plant polyphenols [
36] results in many cellular outcomes and is considered responsible for the epigenetic effects of these molecules [
5] (see below). In particular, it has been reported that Sirt induction (i) counteracts elevated inflammation and lowers cholesterol and triglyceride synthesis [
37]; (ii) reduces oxidative damage markers and increases expression of Nrf2-dependent genes that modulates antioxidant factors in mice fed with a diet rich in olive oil phenolics [
38]; (iii) activates Nrf2 thus attenuating oxidative stress with endothelium protection in mice fed with resveratrol [
39]; (iv) downregulates the pro-inflammatory agent NFκB thus inhibiting the inflammatory response in rat heart subjected to myocardial ischemia and reperfusion [
40]; and (v) inhibits directly the transcriptional activity of PPARγ, with ensuing anti-adipogenic effects [
41]. In addition, a cross-talk does exist between Sirt1 and AMP-activated protein kinase (AMPK), a sensor of the energy state of the cell. The AMPK–Sirt1 relation results in mutual activation [
42] with modulation of the response of the organism to limited nutrients or increased energy demand and autophagy activation. Actually, the latter, in addition to CR [
43], has been reported to be induced by Sirt1 and AMPK [
44] that, in turn, can be activated by resveratrol [
45], or by olive polyphenols [
46,
47,
48].
The data reviewed above indicate that most of the biochemical and physiological effects of plant polyphenols go well beyond their known antioxidant power and support mechanistically their apparent protection against a number of diseases (see below). In this respect, as pointed out above, EVOO polyphenols, notably OLE, have been shown to directly modulate the insulin/IGF1/AKT and the mTOR pathways, whose downregulation results in FOXO3 activation with ensuing transcription of homeostatic genes favoring longevity and reducing inflammatory states. mTOR, a master regulator of cell life, is one of the most potent upstream regulators of autophagy; activation of the latter appears as one of the ways olive polyphenols can induce most of their beneficial effects against neurodegeneration [
46,
47]. Genetic inhibition of autophagy results in degenerative modifications in mammalian cells that compromise the longevity-promoting effects of CR and recall the aging-associated ones; conversely, normal or pathological aging is often associated with impaired autophagy [
49]. A plethora of studies has clearly shown that plant polyphenols, including those in the olive tree, control the phosphorylation state of signaling molecules such as PI3K, Akt, eNOS, AMPK and STAT3; these are involved in the mechanism of ischemic preconditioning [
50] and in autophagy promotion via Sirt1 activation and/or via Ca
2+ increase with ensuing stimulation of the calcium/calmodulin-dependent protein kinase kinase β (CAMKKβ)-AMPK-mTOR pathway [
48]. In cancer cells, the same polyphenols appear to promote cell death by stimulation of apoptosis with features that appear to depend on the cell type [
51,
52,
53], as better specified below.
Overall, the data currently available support the idea that different plant polyphenols, including those from the olive tree, are able to mimic CR effects by affecting the same, or very similar, cellular targets and can therefore be taken into consideration for prevention and/or long-term treatment of aging-associated diseases resulting from chronic inflammation or transcriptional, redox or metabolic derangement.
5. Epigenetic Effects
Epigenetics is defined as the complex of heritable changes to the transcriptome that are distinct from those resulting from the base sequence in the genome but are associated with post-transcriptional gene regulation by non-coding RNAs and with histone and DNA chemical modifications; the latter include DNA methylation, histone methylation, acetylation and phosphorylation [
138]. Non-coding RNAs include microRNAs (miRNAs), small non-coding RNAs that post-transcriptionally modulate gene expression. miRNAs contribute to the control of the expression of both DNA methyltransferases (DNMTs) and histone-modifying enzymes and influence many cellular processes including survival of neuronal cells (reviewed in [
139]). Epigenetic modifications regulate gene expression in a synergistic and cooperative way by changing chromatin arrangement and DNA openness, thus switching on/off a number of genes associated with important physiological and pathological processes (aging, and age-related pathologies, including cancer and neurodegeneration, reviewed in [
140]). These changes are acquired throughout life, including embryonic and fetal development, and depend on environmental clues such as diet, lifestyle and exposure to toxins. When epigenetic modifications are inherited following cell division, they result in the enduring maintenance of the acquired phenotype; however, they can also occur at some time in the course of life and, as such, may remarkably influence phenotypic outcomes in terms of health, disease or risk of disease [
5].
Although numerous compounds have been developed to specifically alter the function of chromatin-modifying enzymes, for example, histone deacetylase (HDAC) inhibitors, we are only beginning to understand the epigenetic effects of dietary compounds. Well-known examples of dietary chromatin-modifying compounds include curcumin, the active constituent of turmeric, which has been shown to be an HDAC inhibitor, as well as the red wine phenol, resveratrol, which activates Sirt1 [
141,
142]. Actually, the investigation of epigenome modifications following administration of plant polyphenols dates back less than one decade. Since then, an increasing number of studies has clearly shown that plant polyphenols, as other nutrients, directly regulate both transcriptional and translational processes by modulating the activity and expression levels of enzymes involved in chemical modifications of histones and DNA (reviewed in [
5]). A growing body of evidence suggests that epigenetic changes triggered by diet nutrients, including plant polyphenols, contribute to preventing some diseases, notably cancer. In particular, plant polyphenols can counteract aging as well as many of its pathological consequences resulting from aberrant epigenetic mechanisms [
52,
139,
143,
144,
145]. In the context of neurodegenerative diseases, the epigenetic modifications have been shown to induce effects similar to those provided by CR [
146]. Accordingly, epigenetics issues targeted by diet polyphenols have become an attractive approach for disease prevention and intervention and provide a rationale for most of the anti-cancer and anti-neurodegeneration power shown by these substances. Epigenetic therapy is an expanding field and is providing clues useful for discovering new drugs, some of which are undergoing clinical investigations, mostly as anti-cancer drugs [
147,
148]. The latter include plant polyphenols such as genistein and quercetin, exploited for their activity as HDAC and DNMT modulators.
The studies reported in the last decade have been carried out mostly on cancer cells and the modifications of gene and protein expression profiles underlying many of the effects listed above can be ascribed to recently shown epigenetic modifications elicited by many plant polyphenols; these include resveratrol, curcumin, epigallocathechins, genistein, quercetin and others in the anti-diabetic, anti-ageing and anti-neurodegeneration fields [
140]. Information about the role of olive polyphenols as epigenome modulators is quite scarce as opposed to other polyphenols. Recent data show that OLE aglycone given orally for eight weeks to TgCRND8 mice, a model of Aβ deposition, downregulates HDAC2 [
48], an enzyme known to be upregulated in AD [
149]. In these mice, the downregulation of HDAC2 resulted in a significant increase in the level of histone acetylation, in particular of H3 at K9 and of H4 at K5 [
48]. Histone acetylation has been reported to improve cognitive deficits in animal models of AD and its indication is considered a promising novel therapeutic strategy against AD [
150]. A recent
in silico molecular modeling study combined with known experimental affinities for controls has identified potential chromatin-modifying compounds from
Olea europaea; in particular, HT was highlighted as a potential inhibitor of HDAC6 and lysine-specific histone demethylase 1 (LSD1) following its high affinity for binding to the active site of various chromatin-modifying enzymes [
151]. Other recent studies have shown that an olive oil-enriched diet increases global DNA methylation in the mammary gland and in a murine model of breast cancer induced by dimethylbenz(a)anthracene (DMBA) [
152]. Finally, EVOO or its phenolic compounds have been reported to modulate the expression of the CNR1 gene encoding for the type 1 cannabinoid receptor via epigenetic mechanisms, both in rats and in human Caco-2 colon cancer cells [
153].
In spite of the limited information on the epigenetic effects of olive polyphenols presently available, the similarities between many effects of different plant polyphenols at the molecular level hold promise that the well documented epigenetic effects reported for many other plant polyphenols could be largely retrieved also in olive polyphenols. In conclusion, modulation of epigenetic flaws by natural polyphenols appears as a promising subject for the discovery of new compounds effective against chronic diseases, even though the present clinical studies have been dedicated prevalently to deciphering polyphenol effects in cancer treatment. HDACs and DNMTs are promising objectives for the control of human pathologies; the modulation of their activity is affected by plant polyphenols, some of which are found in significant amounts in widely used foods that characterize the MD and the Asian diet. However, the data on the epigenetic effects of olive polyphenols are still scarce and further research is needed to increase the information necessary to propose the possible use of these substances as epigenome modulators in humans.
6. Epidemiological Studies and Clinical Trials with Olive Oil and Its Polyphenols
During the last decade, the beneficial properties of EVOO and EVOO polyphenols for human health have been assessed in epidemiological studies and clinical trials. A number of these studies, often carried out on a limited number of patients, have been cited in the preceding sections [
83,
84,
87,
103,
104,
107]. In this section, the studies carried out on large cohorts of participants will be reviewed (
Table 6).
One of the most cited surveys on protection against neurodegeneration and cognitive decline by olive oil is the “Three-City Study” that enrolled around 7000 elderly subjects. The results showed lower odds of cognitive deficit for those subjects who used olive oil moderately (just for cooking or dressing) or intensively (for cooking and dressing) as opposed to those who never used it [
154]. In the same cohort, a lower incidence of stroke in people using higher amounts of olive oil was also observed [
155]. The results of this study were confirmed by two multicenter, randomized, controlled trials, the so-called PREvención con DIeta MEDiterránea (PREDIMED) and PREDIMED-NAVARRA studies carried out in Spain on people at high cardiovascular risk. The PREDIMED study was a primary prevention trial originally designed to test the long-term effects of the MD on the incidence of CVD in people with high cardiovascular risk. The cohort was also evaluated for cognitive performance, after adjustment for several potentially interfering factors. It emerged that an intervention with a MD enriched in EVOO (better than with wine or nuts) significantly improved cognition, and that EVOO phenolic content was the main factor responsible for this result [
21,
156]. The main goal of the study was assessing CVD prevention; in this sense, the trial suggested that EVOO consumption is associated with a reduced risk of CVD and mortality [
157,
158].
The role played by polyphenols in cardiovascular protection was further confirmed by two sub-studies on patients subjected to MD high in polyphenols (from nuts or EVOO). Total polyphenol excretion by people adopting a MD rich either in nuts or EVOO was positively correlated with changes in plasma levels of triglycerides, glucose and nitric oxide (NO); moreover, the statistically significant increase in plasma NO levels was associated with a reduction in systolic and diastolic blood pressure [
159]. Accordingly, data from a subsample (
n = 990) of the PREDIMED study have shown that the MD significantly decreases LDL oxidation only when it is enriched in EVOO with medium-high phenolic content [
160].
Overall, as outlined by a recent survey, the PREDIMED study in patients at the MD rich in phenols (from nuts or EVOO) showed a significant improvement of classical and emerging CVD risk factors, including inflammation, oxidative stress blood pressure, carotid atherosclerosis, insulin resistance, lipoproteins and lipid profiles [
161]. Breast cancer incidence was also investigated in the PREDIMED cohort. Thirty-five out of the 4282 women enrolled in the survey displayed confirmed cases of invasive breast cancer, whose risk was reduced by 68% in the EVOO group as compared with the low-fat group even after accounting for factors such as age, body mass index, exercise and drinking habits. The risk of being affected by invasive breast cancer was highest for women who were instructed to eat less fat (2.9 cases for 1000 person-years). This value was compared to a diagnosis rate of 1.8 cases per 1000 person-years for women on the MD supplemented with nuts and a rate of 1.1 cases per 1000 person-years for women on the MD with increased EVOO, suggesting a better protection against breast cancer of EVOO over nuts [
162].
More recently, a cross-sectional sampling population study was carried out among the whole Spanish people. The study selected, in 100 health centers, 4572 individuals aged >18 years, representative of the Spanish population. Clinical, demographic and lifestyle parameters were considered, together with physiological parameters (body weight and height, body mass index, waist and hip measurement, blood pressure and oral glucose tolerance). The participants were analyzed considering whether they were consumers of olive oil or sunflower oil. The main outcome of the study showed that the consumption of olive oil was associated with significant beneficial effects on several cardiovascular risk factors, particularly in the presence of obesity and a sedentary lifestyle, and with a significant improvement of impaired glucose tolerance and insulin resistance [
163]. Another study aimed at examining the association between olive oil intake and the incidence of T2DM was carried out in the USA by following 59,930 women aged 35–65 years from the Nurses’ Health Study (NHS) and 85,157 women aged 26–45 years from the NHS II with no diabetes, CVD and cancer at baseline. The diet was controlled and validated by food-frequency questionnaires. Hidden T2DM cases were identified and confirmed by questionnaires. At the end of the 22 years follow-up, 5738 and 3914 cases of T2DM were documented in the NHS and NHSII, respectively. The outcome compared people taking at least one tablespoon of olive oil daily with those who never assumed olive oil. The results suggested that higher olive oil intake was associated with a modestly lower risk of T2DM; conversely the risk was raised in people who substituted olive oil with other lipids [
164]. Finally, a very recent study has been conducted with 25 healthy subjects randomly allocated in a cross-over design to a Mediterranean-type meal supplemented with or without 10 g EVOO/day or the same amount of corn oil. The lipid profile and glycemic parameters related to glucose tolerance, determined two hours after the meal, showed that EVOO improves post-prandial glucose and LDL-cholesterol, confirming the anti-atherosclerotic power of the MD [
165].
Overall, these surveys support the notion that natural EVOO phenols might counteract age-associated cognitive decline, CVD and cancer, particularly breast cancer. Accordingly, many clinical trials have been conducted employing olive oil or polyphenol-enriched olive extracts, but the results are still scarce even from those studies that were completed, except for trials aimed at investigating the effect of EVOO and its polyphenols against several conditions associated with CVD (oxidative stress, inflammation, haemostasis, endothelial function and blood pressure). In a small crossover trial, carried out in the context of the EUROLIVE (Effect of Olive Oil Consumption on Oxidative Damage in European Populations) study (Trial number: ISRCTN09220811), 200 participants were subjected to three rounds of daily administration of 25 mL of three olive oils with a different phenolic content (low, 2.7 mg/kg of olive oil; medium, 164 mg/kg; medium-high, 366 mg/kg) for three weeks preceded by a two-week washout period. The results showed a linear decrease of the total cholesterol/HDL-cholesterol ratio and of oxidative stress markers with the increase of the phenolic content of the olive oil [
166]. Protection against atherosclerosis was confirmed by a sub-set of the same trial with 25 healthy volunteers fed for three weeks with 25 mL/d uncooked olive oil with a medium-high polyphenol-content (366 mg/kg) or a low-polyphenol-content (2.7 mg/kg) [
167]. The results provided a first-level evidence that the phenolic content is the main factor responsible for the health benefits of EVOO. This correlates also with the results coming from the randomised, controlled clinical trial involving pre-hypertensive patients fed with 30 mL of two similar olive oils, a functional EVOO enriched with its phenolic compounds (961 mg/kg,) or a medium polyphenol content EVOO (289 mg/kg). Data from this study support a significant upregulation of genes regulating the cell-HDL cholesterol efflux [
81]. That olive polyphenols increase human HDL functionality has recently been confirmed by a subsample of the EUROLIVE study, in which it was shown that the consumption of olive oil with high phenolic content increased the cholesterol efflux from macrophages mediated by HDL [
82].
Most of the above studies were aimed at assessing the effects of EVOO and its polyphenols after a relatively long-term ingestion; studies were also carried out to determine the effects of an acute administration of olive oil, particularly in the case of protection against postprandial hyperlipidaemia and the associated inflammation. A study conducted on 20 obese subjects that were given muffins made with different oils previously subjected to 20 heating cycles, showed that oils rich in phenols, whether natural (EVOO) or artificially added, reduced postprandial inflammation; this outcome was determined by the activation of nuclear NFκB, the cytosolic levels of IκB-α, an NFκB inhibitor, the mRNA levels of p65, IKKβ, and IKKα (NFκB subunits and activators) and by the levels of lipopolysaccharide (LPS) and other pro-inflammatory molecules (TNF-α, IL-1β, IL-6, migration inhibiting factor (MIF), JNK); seed oil (sunflower) failed to produce similar results [
168]. The protection against post-prandial oxidative stress was confirmed by other randomised, cross-over, controlled human studies showing that the serum antioxidant capacity was increased after EVOO ingestion at the same single doses (40 or 50 mL) at which oxidative stress normally occurs if the ingested oil is not EVOO [
169]; the study also reported a lower lipid oxidative damage in subjects fed with an olive oil with high, rather than low, phenolic content [
170,
171]. Finally, a comprehensive review synthetically reports the results of randomized, controlled trials showing the efficacy of EVOO in lowering numerous inflammation markers such as thromboxan2 (TBX
2), leukotriene B4 (LTB
4), intercellular adhesion molecule 1 (ICAM-1) and vascular cell adhesion molecule 1 (VCAM-1), TNF-α, IL-1β, IL-6, MIF, JNK and LPS, NFκB and its activators, high-sensitivity C-reactive protein (hs-CRP), asymmetric dimethylarginine (ADMA) relevant in the context of several pathologies including CVD [
172].
7. Bioavailability of Olive Polyphenols
A problem associated with the use of olive and other plant polyphenols is their reduced bioavailability due both to incomplete intestinal absorption and to rapid biotransformation favoring urinary excretion. Moreover, in the case of the brain, orally ingested polyphenols must cross an additional barrier, the blood–brain barrier, in addition to that represented by the enterocytes. With few exceptions, only polyphenol aglycones can be absorbed in the small intestine [
173] and deglycosylation by β-glucosidase in small intestinal epithelial cells has been focused on as a crucial step in absorption and ensuing metabolism of dietary polyphenols, notably the glycated forms [
174]. Once released from the enterocyte into the lymph and therefrom into the blood stream, most polyphenols undergo substantial biotransformations including methylation, glucuronidation, sulphation and thiol conjugation [
175] that alter their chemical properties, favor their excretion and, possibly, provide them new biological activities [
176]. Moreover, recent research has highlighted the importance for polyphenol bioavailability of the colonic microflora that can extensively metabolize and chemically modify polyphenols [
177]. However, recent studies on this theme carried out both on rats and humans have shown that these compounds are indeed absorbed in discrete amounts from the intestine and rapidly distributed through the blood flow to the whole organism, including the brain. In particular, recent data clearly indicate that, similarly to other polyphenols, the glycated and (preferentially) the aglycone forms of OLE are indeed absorbed and found in the plasma after ingestion both in rats [
178,
179] and in humans [
179,
180,
181,
182,
183]. From the plasma they, at least in part, are distributed to different organs and tissues, including the brain, where they, or some derivatives, have been found [
179]. Finally, a recent metabolite-profiling study with cultured breast cancer cells treated with an olive leaf extract has shown that OLE is the main polyphenol found inside the cells, suggesting its ability to cross the plasma membrane in this cell line [
184]. As a confirmation, a very recent study has shown that OLE interacts with synthetic phospholipid membranes, but the extent of the interaction depends on membrane lipid composition and is favored by the presence of anionic lipids, suggesting specific interactions [
185]. These data agree with those reported in a recent study showing that a number of polyphenols (those from the olive tree were not included) are able to protect the mitochondrial membrane from permeabilization by amyloid oligomers, suggesting some interference with the formation of the oligomer–membrane complex [
186]. Finally, in a recent study we have shown that the OLE metabolite, HT, arising mainly from acid hydrolysis in the stomach, is found in the brain of TgCRND8 mice fed with OLE for eight weeks [
48]; this finding agrees with previous data reported in rats [
178] supporting the ability of some OLE derivatives, including HT, to cross the blood–brain barrier, even though its generation from OLE once it has crossed the blood-brain-barrier cannot be excluded.
Accurate studies on the effective daily dose of olive polyphenols to be administered to humans to get significant protection are still lacking. What must be taken into account is that, apparently, the amount of OLE and other plant polyphenols present in foods is not adequate to ensure daily doses suitable to get short-term acute effects. However, clinical and experimental evidence suggests that the continuous assumption of foods containing moderate amounts of these molecules can be effective in the long term, also due to their possible accumulation as lipophilic molecules, producing a low-intensity continuative stimulus of cell defenses against oxidative stress, amyloid deposition and other alterations underlying age-associated pathologies. Nevertheless, the low daily consumption of olive oil polyphenols with a typical MD suggests the value of the integration of polyphenol-enriched olive leaf extracts that can intensify, in the short-term, the beneficial effects of these molecules.
8. Conclusions
At the end of our itinerary through the nutraceutical properties of EVOO and its polyphenols we should recognize that the research in this field is actively proliferating, hence it is becoming mandatory to accommodate the plethora of biochemical, cellular and physiological effects of EVOO polyphenols in a coherent picture. In fact, looking at the multitude of cellular effects elicited by these compounds, that often look similar to those elicited by other plant polyphenols, one could have the uncomfortable sensation that their action is rather non-specific. Moreover, in some cases, contradictory results have been reported; these can possibly result, among others, from differences in the study design and administered doses of olive polyphenols in the case of clinical trials, the type of animal (race, model, wild-type or transgenic) in the case of animal studies, or the type of investigated cells and cell culture conditions. However, we do believe that some common traits are emerging that will pave the way to mechanistically defining the nutraceutical activity of EVOO and olive polyphenols. These traits can be summarized as follows.
First, plant, notably olive, polyphenols act mainly as signaling molecules. Although the direct target of interaction must still be identified, a calcium-mediated activation of AMPK via CaMKKβ has been reported for OLE aglycone [
46]; oleocanthal was also found to activate AMPK [
187]. Aging can be considered as a phenomenon driven by over-activation of the nutrient-sensor mTOR gerogene following a decline, or lack of responsiveness to activation signals of AMPK, an energy-sensing protein and a critical mTOR gerosuppressor [
69]. AMPK is a sort of node in the intricate signaling network of the cell, where several physiological and pathological pathways (autophagy, anabolism/catabolism, apoptosis, cell proliferation, inflammation, neurodegeneration) intersect [
188,
189,
190,
191,
192]. Accordingly, AMPK activation by olive polyphenols is a possible explanation for most of the pleiotropic activities of these substances. In general, specific receptor–polyphenol interactions have not been clearly identified; rather, it is believed that these compounds interact freely with the cell membrane bilayer [
185,
186], modifying its permeability properties, notably to calcium [
46]. However, a receptor, the TRPA1 receptor ion channel, spatially restricted to the throat, has recently been identified as the possible responsible for the pungency of oleocanthal sensed in the throat [
193].
Second, EVOO polyphenols directly participate to the redox balance of the cell. They perform this role not simply as antioxidants but, in certain circumstances, also as mild pro-oxidants, by up-regulating the antioxidant defenses of the cell, thus acting as hormetic factors [
69]. This has been shown for both HT that, in the presence of peroxidases, can undergo a redox cycling that generates superoxide, an inducer of Mn-SOD expression [
194] and for tyrosol, that increases
C. elegans lifespan also by activating the heat shock response [
195]. The role of EVOO phenols as “xenohormetic agents” has been analyzed by Menendez and coll. in their transcriptome analysis of cells exposed to crude EVOO polyphenol extracts highly enriched in secoiridoids OLE and decarboxymethyl OLE; their data confirm an involvement, among others, of the activation of anti-aging/cellular stress-like genes, including those for ER stress, the unfolded protein response, Sirt1 and Nrf2 signaling [
69].
Third, it is generally believed that plant polyphenols can directly interact with other molecules or molecular complexes through aromatic stacking, hydrophobic interaction, or chemical crosslinking. Actually, this mechanism seems to underlie the inhibition of toxic amyloid aggregation by EVOO phenols. In the case of oleocanthal, it usually promotes the conversion of monomers and oligomers of amyloidogenic proteins/peptides (such as Tau and Aβ) to high molecular weight aggregates by chemical crosslinking thanks to its couple of aldehyde groups [
136,
196,
197]. OLE aglycone differs from oleocanthal by the absence of aldehyde groups and the presence of a methoxycarbonyl group. Therefore, OLE is not a chemical crosslinker and its anti-amyloidogenic activity should occur differently [
130]. Structure comparison analysis suggests that the number of phenolic rings is a key responsible for the polyphenol’s efficacy in remodeling amyloid oligomers; in particular, two aromatic rings, at least one with a hydroxyl group, appears to be the minimal structural requirement needed by phenolic aglycones to remodel Aβ oligomers by interfering with aromatic group stacking [
132]. Actually, the main EVOO polyphenols do not respond to this description, since they have just one phenolic ring. Nonetheless, it has been clearly shown that OLE physically interacts with the Aβ peptide [
131] and that the Phe4–Glu11 sequence, together with the Leu17–Lys28 hydrophobic region, of Aβ40/42 are responsible for the non-covalent interaction that occurs in the 17–21 region of the peptide (Leu-Val-Phe-Phe-Ala), which includes two Phe residues [
197,
198]. Interestingly, the Aβ sequence critical for amyloid fibrillization overlaps such OLE-binding regions. This finding supports the hypothesis that aromatic stacking or, more generally, hydrophobic interactions, would be the molecular mechanism underlying inhibition of amyloid aggregation by this polyphenol [
197]. Hydrophobic interaction is probably also involved in the incorporation of olive polyphenols into LDL [
199].
Fourth, in spite of the reduced bioavailability due to incomplete intestinal absorption, microbiota metabolism and biotransformation in tissue, plant, notably olive, polyphenols are indeed distributed throughout the organism and have been found in tissues, including the brain, further supporting their ability to positively interfere with pathological states and/or their prodromal conditions.
Further efforts are needed to mechanistically define the biochemical and biological activities of EVOO and olive polyphenols as well as the pharmacokinetics and pharmacodynamics underlying their effective doses in humans and the dose-dependence of their effects. The structure–activity relationship of olive polyphenols must still be deciphered as well; the latter could be the basis of engineering new drugs starting from the molecular scaffolds of these substances. Finally, more clinical trials are needed to overcome the limits of those currently reported and some of their conflicts. However, we do believe that the four points that we have extracted from the large body of scientific literature on OLE, HT and oleocanthal could be useful landmarks with which to orient future research and organize present and future data into a coherent frame.







{kind=link}
{kind=link}
{kind=link}
{kind=link}