
Longitudinal Effects of Embryonic Exposure to Cocaine on Morphology, Cardiovascular Physiology, and Behavior in Zebrafish

Abstract
:
1. Introduction
2. Results
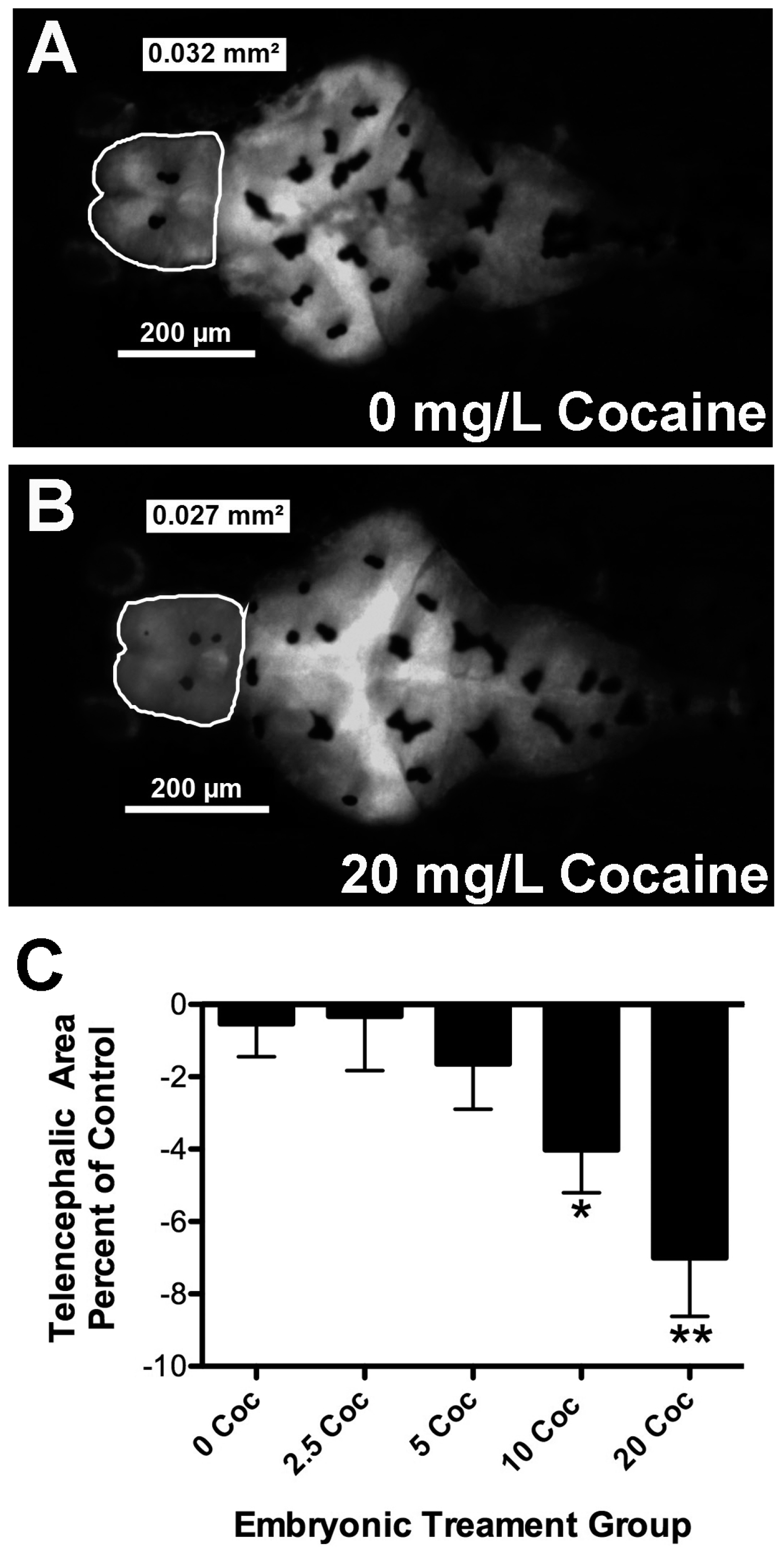
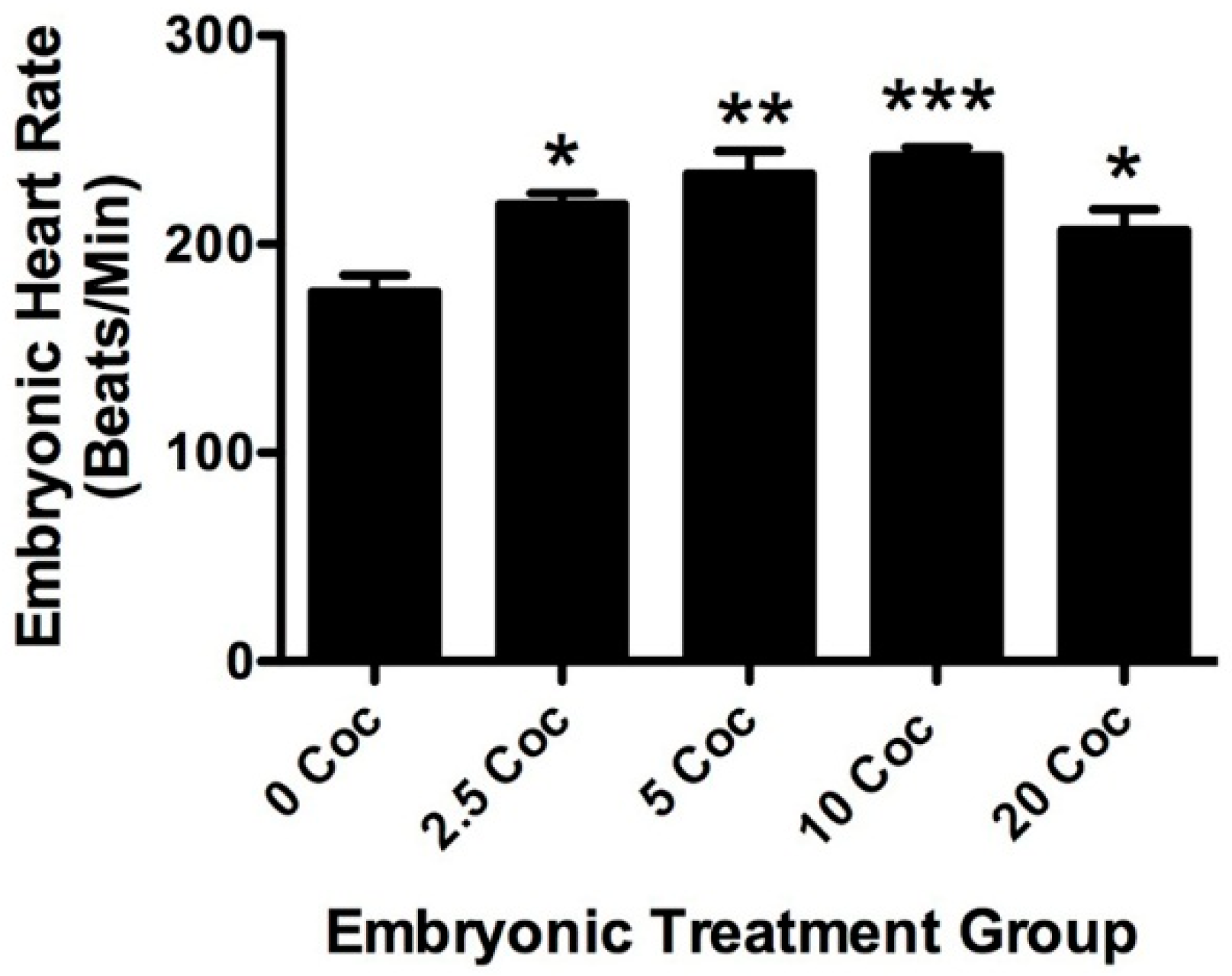
2.1. Cocaine Dosage Effects on Embryonic Health and Brain Development
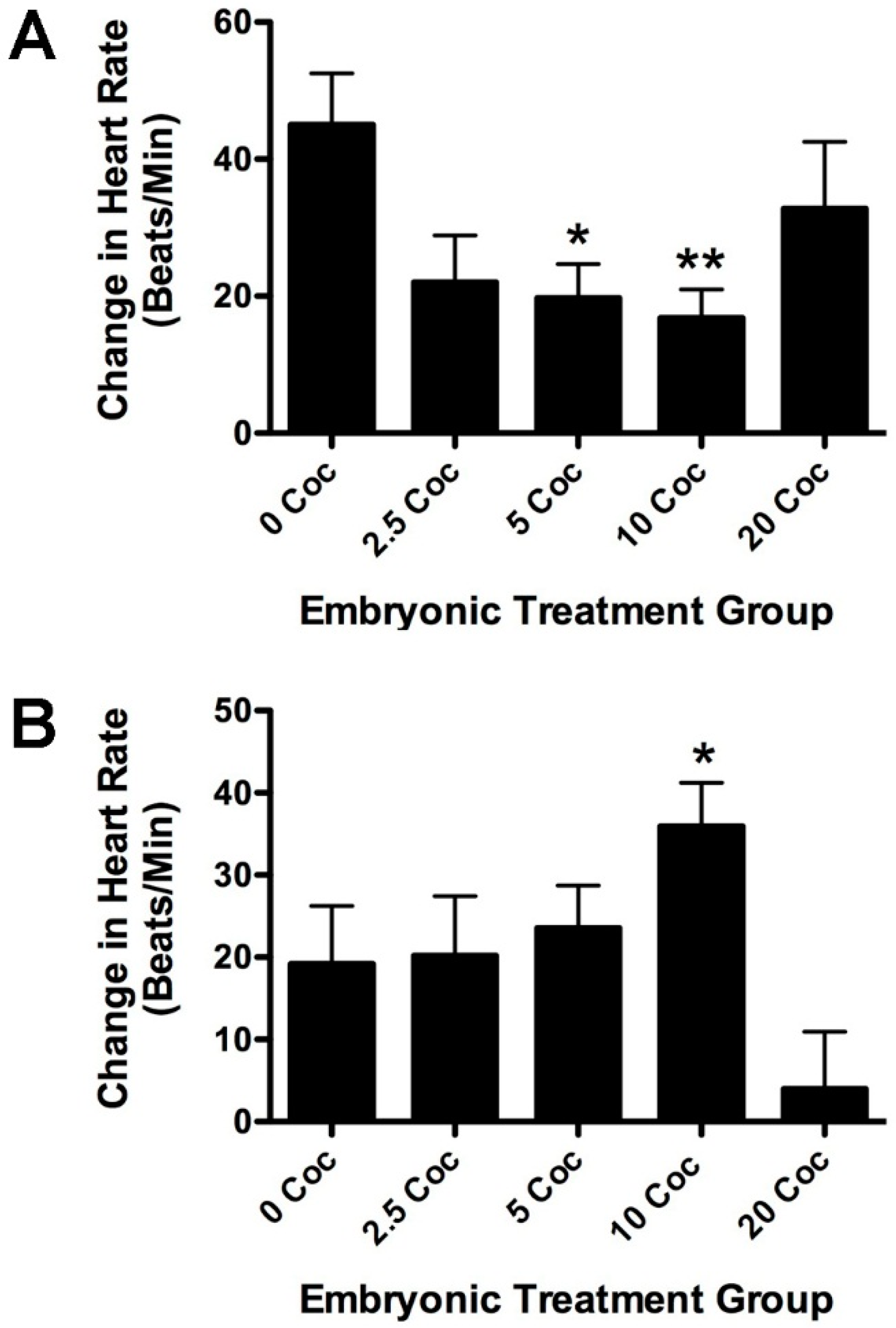
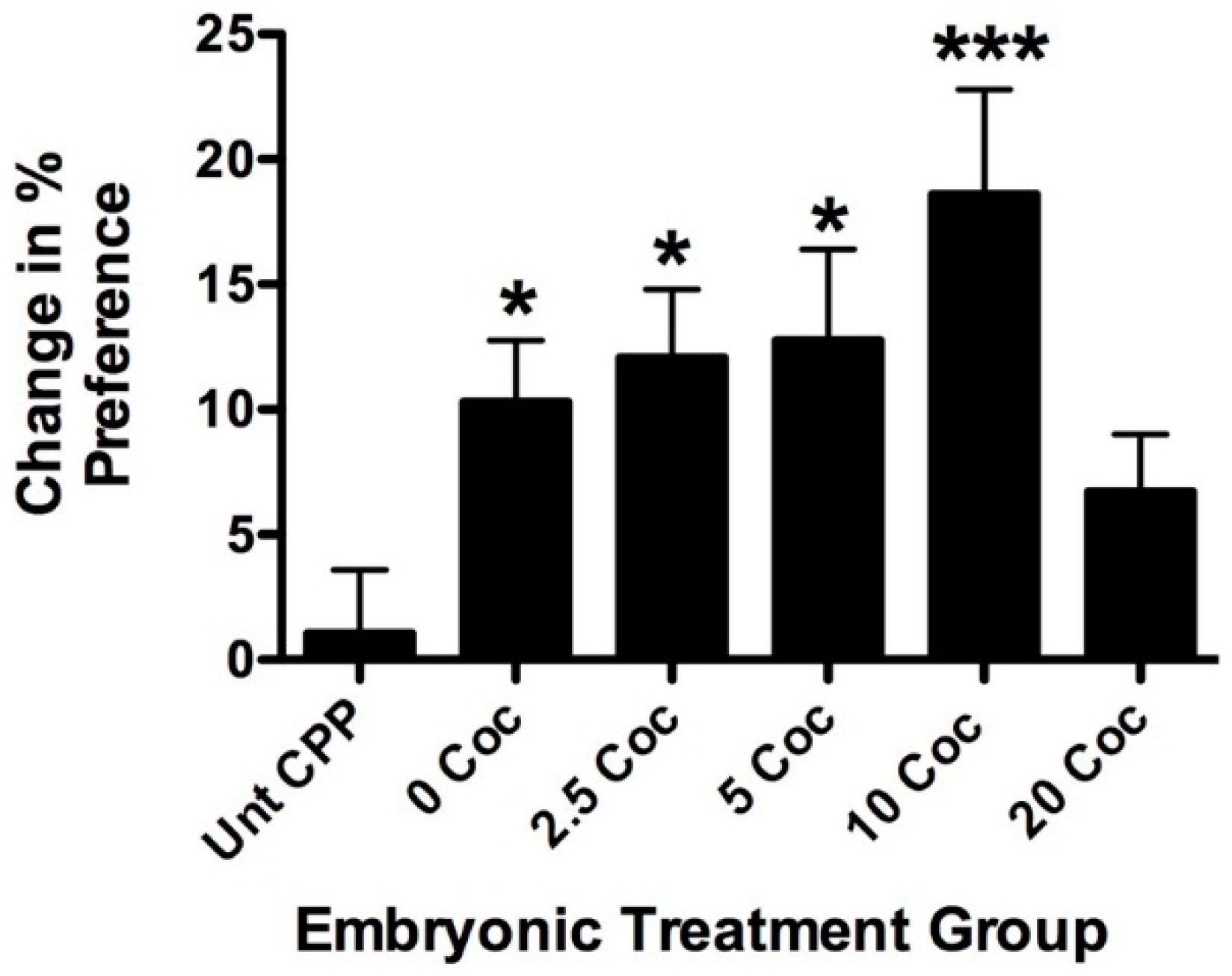
2.2. Longitudinal Effects of Cocaine on Zebrafish Health, Cardiovascular Physiology and Behavior
3. Discussion
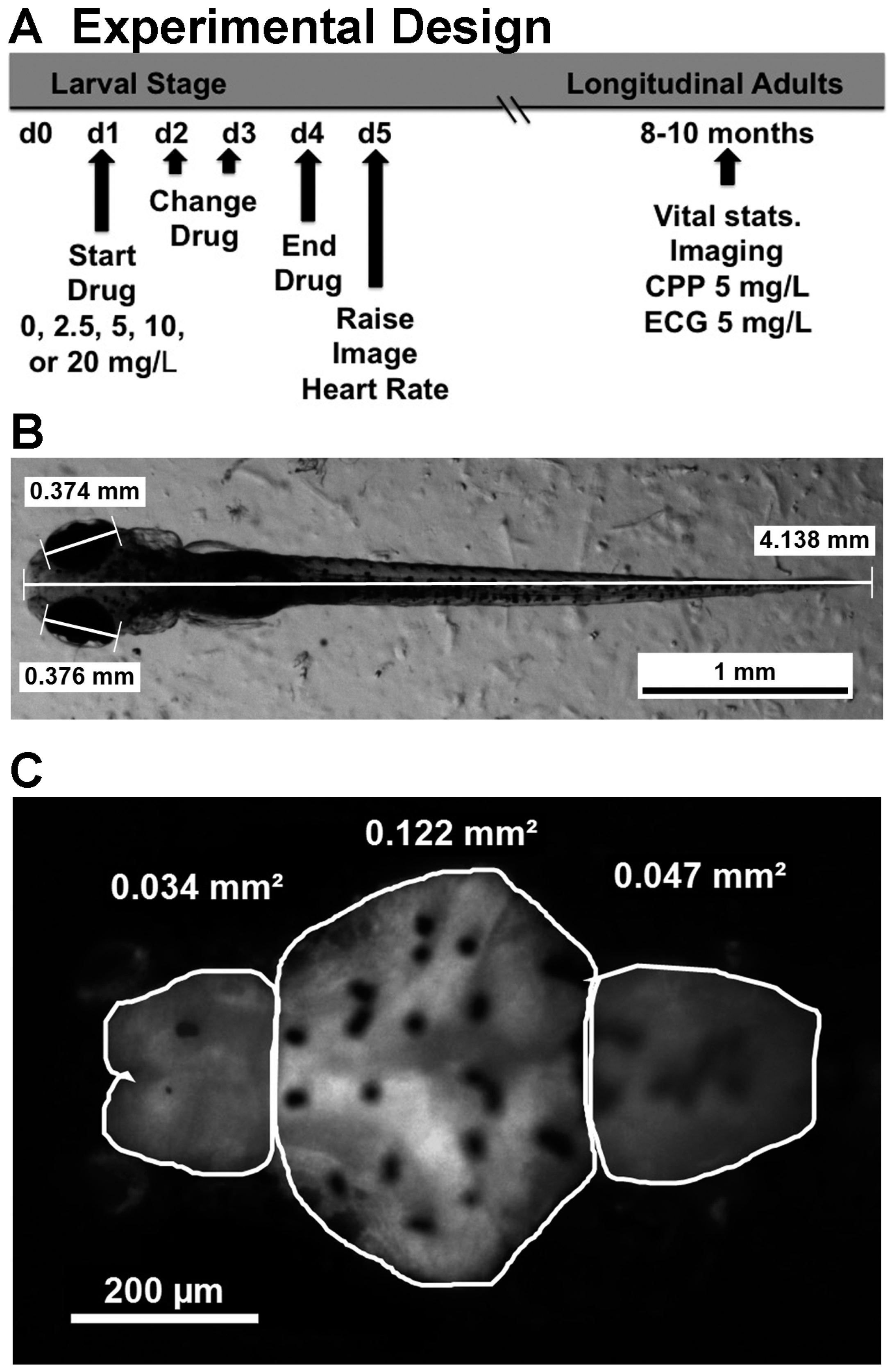
4. Materials and Methods
4.1. Fish Maintenance
4.2. Embryonic Drug Treatment
4.3. Imaging and Measuring Size and Brain Morphology
4.4. Measuring Embryonic Heart Rate
4.5. Conditioned Place Preference
4.6. Electrocardiograms in Adult Zebrafish
5. Statistics
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- NIDA: Drug Facts: Nationwide Trends. Available online: http://www.drugabuse.gov/publications/drugfacts/nationwide-trends (accessed on 30 May 2016).
- NIDA: Trends & Statistics. Available online: http://www.drugabuse.gov/related-topics/trends-statistics (accessed on 30 May 2016).
- NIDA: Prenatal Exposure to Drugs of Abuse. Available online: http://www.drugabuse.gov/sites/default/files/prenatal.pdf (accessed on 30 May 2016).
- Frank, D.A.; Augustyn, M.; Knight, W.G.; Pell, T.; Zuckerman, B. Growth, development, and behavior in early childhood following prenatal cocaine exposure: A systematic review. JAMA 2001, 285, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Grewen, K.; Burchinal, M.; Vachet, C.; Gouttard, S.; Gilmore, J.H.; Lin, W.; Johns, J.; Elam, M.; Gerig, G. Prenatal cocaine effects on brain structure in early infancy. Neuroimage 2014, 101, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J. Effect of cocaine use on the fetus. N. Engl. J. Med. 1992, 327, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q. Postnatal consequences of prenatal cocaine exposure and myocardial apoptosis: Does cocaine in utero imperil the adult heart? Br. J. Pharmacol. 2005, 144, 887–888. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Zhang, L. Short- and long-term adverse effects of cocaine abuse during pregnancy on the heart development. Ther. Adv. Cardiovasc. Dis. 2009, 3, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Minnes, S.; Singer, L.; Min, M.O.; Wu, M.; Lang, A.; Yoon, S. Effects of prenatal cocaine/polydrug exposure on substance use by age 15. Drug Alcohol Depend. 2014, 134, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.A.; Goldschmidt, L.; Larkby, C.; Day, N.L. Effects of prenatal cocaine exposure on adolescent development. Neurotoxicol. Teratol. 2015, 49, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Lidow, M.S. Consequences of prenatal cocaine exposure in nonhuman primates. Dev. Brain Res. 2003, 147, 23–36. [Google Scholar] [CrossRef]
- Malanga, C.J.; Kosofsky, B.E. Does drug abuse beget drug abuse? Behavioral analysis of addiction liability in animal models of prenatal drug exposure. Dev. Brain Res. 2003, 147, 47–57. [Google Scholar] [CrossRef]
- McCarthy, D.M.; Kabir, Z.D.; Bhide, P.G.; Kosofsky, B.E. Effects of prenatal exposure to cocaine on brain structure and function. Prog. Brain Res. 2014, 211, 277–289. [Google Scholar] [PubMed]
- McMurray, M.S.; Oguz, I.; Rumple, A.M.; Paniagua, B.; Styner, M.A.; Johns, J.M. Effects of prenatal cocaine exposure on early postnatal rodent brain structure and diffusion properties. Neurotoxicol. Teratol. 2015, 47, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Crandall, J.E.; Hackett, H.; Tobet, S.A.; Kosofsky, B.E.; Bhide, P.G. Cocaine exposure decreases GABA neuron migration from the ganglionic eminence to the cerebral cortex in embryonic mice. Cerebral Cortex 2004, 14, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.-Q.; Malanga, C.J.; Tabit, E.; Kosofsky, B.E. Neuropathological consequences of prenatal cocaine exposure in the mouse. Int. J. Dev. Neurosci. 2004, 22, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Rocha, B.A.; Mead, A.N.; Kosofsky, B.E. Increased vulnerability to self-administer cocaine in mice prenatally exposed to cocaine. Psychopharmacology 2002, 163, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Hall, Z.J.; De Serrano, A.R.; Rodd, F.H.; Tropepe, V. Casting a wider fish net on animal models in neuropsychiatric research. Prog. Neuro-Psychopharmacol. Biol. Psychiatr. 2014, 55, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Klee, E.W.; Schneider, H.; Clark, C.J.; Cousin, M.A.; Ebbert, J.O.; Hooten, W.M.; Karpyak, V.M.; Warner, D.O.; Ekker, S.C. Zebrafish: A model for the study of addiction genetics. Hum. Genet. 2012, 131, 977–1008. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueff, A.V. Zebrafish models for translational neuroscience research: From tank to bedside. Trends Neurosci. 2014, 37, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Ganz, J.; Kroehne, V.; Freudenreich, D.; Machate, A.; Geffarth, M.; Braasch, I.; Kaslin, J.; Brand, M. Subdivisions of the adult zebrafish pallium based on molecular marker analysis. F1000Res 2014, 3, 308. [Google Scholar] [CrossRef] [PubMed]
- Northcutt, R.G. Forebrain evolution in bony fishes. Brain Res. Bull. 2008, 75, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F. Secondary neurogenesis and telencephalic organization in zebrafish and mice: A brief review. Integr. Zool. 2009, 4, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Rink, E. The teleostean forebrain: A comparative and developmental view based on early proliferation, Pax6 activity and catecholaminergic organization. Brain Res. Bull. 2002, 57, 363–370. [Google Scholar] [CrossRef]
- Lau, B.Y.B.; Mathur, P.; Gould, G.G.; Guo, S. Identification of a brain center whose activity discriminates a choice behavior in zebrafish. PNAS 2011, 108, 2581–2586. [Google Scholar] [CrossRef] [PubMed]
- Von Trotha, J.W.; Vernier, P.; Bally-Cuif, L. Emotions and motivated behavior converge on a amygdala-like structure in the zebrafish. Eur. J. Neurosci. 2014, 40, 3302–3315. [Google Scholar] [CrossRef] [PubMed]
- Darland, T.; Dowling, J.E. Behavioral screening for cocaine sensitivity in mutagenized zebrafish. PNAS 2001, 98, 11691–11696. [Google Scholar] [CrossRef] [PubMed]
- Darland, T.; Mauch, J.T.; Meier, E.M.; Hagan, S.J.; Dowling, J.E.; Darland, D.C. Sulpiride, but not SCH23390, modifies cocaine-induced conditioned place preference and expression of tyrosine hydroxylase and elongation factor 1a in zebrafish. Pharmacol. Biochem. Behav. 2012, 103, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Mersereau, E.J.; Poitra, S.L.; Espinoza, A.; Crossley, D.A., II; Darland, T. The effects of cocaine on heart rate and electrocardiogram in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 172–173, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nicolaysen, L.C.; Justice, J.B., Jr. Effects of cocaine on release and reuptake of dopamine in vivo: Differentiation by mathematical modeling. Pharmacol. Biochem. Behav. 1988, 31, 327–335. [Google Scholar] [CrossRef]
- Taylor, D.; Ho, B.T. Comparison of inhibition of monoamine uptake by cocaine, methylphenidate and amphetamine. Res. Commun. Chem. Pathol. Pharmacol. 1978, 21, 67–75. [Google Scholar] [PubMed]
- Matthews, J.C.; Collins, A. Interactions of cocaine and cocaine congeners with sodium channels. Biochem. Pharmacol. 1983, 32, 455–460. [Google Scholar] [CrossRef]
- Hall, F.S.; Sora, I.; Drgonova, J.; Li, X.-F.; Goeb, M.; Uhl, G.R. Molecular mechanisms underlying the rewarding effects of cocaine. Annu. N. Y. Acad. Sci. 2004, 1025, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Kuhar, M.J.; Ritz, M.C.; Boja, J.W. The dopamine hypothesis of the reinforcing properties of cocaine. Trends Neurosci. 1991, 14, 299–302. [Google Scholar] [CrossRef]
- Kloner, R.A.; Hale, S.; Alker, K.; Rezkalla, S. The effects of acute and chronic cocaine use on the heart. Circulation 1992, 85, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, B.G.; Rezkalla, S.; Kloner, R.A. Cardiovascular effects of cocaine. Circulation 2010, 122, 2558–2569. [Google Scholar] [CrossRef] [PubMed]
- Bozzi, Y.; Borrelli, E. Dopamine in neurotoxicity and neuroprotection: What do D2 receptors have to do with it? Trends Neurosci. 2006, 29, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.L.; Stanic, D.; Drago, J.; Borrelli, E.; Finklestein, D.I.; Horne, M.K. Effects of long term treatment with dopamine agonists and antagonists on terminal arbor size. Eur. J. Neurosci. 2002, 16, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U.; Beyer, C.; Oestreicher, A.B.; Reisert, I.; Schilling, K.; Pilgrim, C. Activation of dopaminergic D1 receptors promotes morphogenesis of developing striatal neurons. Neuroscience 1996, 74, 453–460. [Google Scholar] [CrossRef]
- Todd, R.D. Neural development is regulated by classical neurotransmitters: Dopamine D2 receptor stimulation enhances neurite outgrowth. Biol. Psychiatr. 1992, 31, 794–807. [Google Scholar] [CrossRef]
- Lopez-Bellido, R.; Barreto-Valer, K.; Rodriguez, R.E. Substance P mRNA expression during zebrafish development: influence of mu opioid receptor and cocaine. Neuroscience 2013, 242, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Lillesaar, C. The serotonergic system in fish. J. Chem. Neuroanat. 2011, 41, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.M. Catecholaminergic systems in the zebrafish III. Organization and projection pattern of medullary dopaminergic and noradrenergic neurons. J. Comp. Neurol. 1997, 381, 411–427. [Google Scholar] [CrossRef]
- Rink, E.; Wullimann, M.F. The teleosten (zebrafish) dopaminergic system ascending to the subpallium (striatum) is located in the basal diencephalon (posterior tuberculum). Brain Res. 2001, 889, 316–330. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ruuskanen, J.O.; Wullimann, M.F.; Vernier, P. Differential expression of dopaminergic cell markers in the adult zebrafish forebrain. J. Comp. Neurol. 2011, 519, 576–598. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Strahle, U.; Scolpp, S. Neurogenesis in zebrafish-from embryo to adult. Neural Dev. 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.S. Perinatal cocaine exposure impairs myocardial B-adrenoceptor signaling in the neonatal rat. Anesth. Analg. 2000, 90, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.S.; Takuma, S.; Lui, R.; Homma, S. The effect of maternal cocaine exposure on neonatal rat cardiac function. Anesth. Analg. 2003, 97, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Schwerte, T.; Prem, C.; Mairosl, A.; Pelster, B. Development of the sympatho-vagal balance in the cardiovascular system in zebrafish (Danio rerio) characterized by power spectrum and classical signal analysis. J. Exp. Biol. 2006, 209, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Kastenhuber, E.; Kratochwil, C.F.; Ryu, S.; Schweitzer, J.; Driever, W. Genetic dissection of dopaminergic and noradrenergic contributions to catecholaminergic tracts in early larval zebrafish. J. Comp. Neurol. 2010, 518, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Boyer, B.; Ernest, S.; Rosa, F. Egr-1 induction provides a genetic response to food aversion in zebrafish. Front. Behav. Res. 2013, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hinz, F.I.; Aizenberg, M.; Tushev, G.; Schuman, E. Protein synthesis-dependent associative long-term memory in larval zebrafish. J. Neurosci. 2013, 33, 15382–15387. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.C.; Bill, B.R.; Glanzman, D.L. Learning and memory in zebrafish larvae. Front. Neural Circuits 2013, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, Y.; Rampersad, M.; Gerlai, R. Embryonic alcohol exposure impairs the dopaminergic system and social behavioral responses in adult zebrafish. Int. J. Neuropsychopharmacol. 2015, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Uhl, G.R.; Drgonova, J.; Hall, F.S. Curious cases: Altered dose-response relationships in addicition genetics. Pharmacol. Ther. 2014, 141, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Malanga, C.J.; Pejchal, M.; Kosofsky, B.E. Prenatal exposure to cocaine alters the development of conditioned place-preference in adult mice. Pharmacol. Biochem. Behav. 2007, 87, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Newton, T.F.; de La Garza, R., II; Kalechstein, A.D.; Liam, N. Cocaine and methamphetamine produce different patterns of subjective and cardiovascular effects. Pharmacol. Biochem. Behav. 2005, 82, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Mejias-Aponte, C.A.; Kiyatkin, E.A. Ventral tegmental area neurons are either excited or inhibited by cocaine’s actions in the peripheral nervous system. Neuroscience 2012, 207, 182–197. [Google Scholar] [CrossRef] [PubMed]
- Ruth, J.A.; Ullman, E.A.; Collins, A.C. An analysis of cocaine effects on locomotor activities and heart rate in four inbred mouse strains. Pharmacol. Biochem. Behav. 1988, 29, 157–162. [Google Scholar] [CrossRef]
- Thomsen, M.; Caine, S.B. Psychomotor stimulant effects of cocaine in rats and 15 mouse strains. Exp. Clin. Psychopharmacol. 2011, 19, 321–341. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D.; Hankin, M.; Li, Z.; Dai, X.; Ding, J. Transgenic zebrafish for studying nervous system development and regeneration. Transgenic Res. 2001, 10, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullman, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.M.; Perkin, B.D.; Amsterdam, A.; Egana, A.; Darland, T.; Matsui, J.I.; Sciascia, S.; Hopkins, N.; Dowling, J.E. Identification of zebrafish insertional mutants with defects in visual system development and function. Genetics 2005, 170, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Vassoler, F.M.; Sadri-Vakili, G. Mechanisms of transgenerational inheritance of addictive-like behaviors. Neuroscience 2014, 264, 198–206. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Embryonic Treatment Condition | 0 mg/L Cocaine | 2.5 mg/L Cocaine | 5.0 mg/L Cocaine | 10.0 mg/L Cocaine | 20.0 mg/L Cocaine |
|---|---|---|---|---|---|
| % Hatched by 48 hpf | 16 ± 15.5 | 39.5 ± 21.3 | 51.5 ± 13.8 * | 59.1 ± 19.5 ** | 65.5 ± 28.9 *** |
| Body size (mm) | 3.97 ± 0.16 | 3.94 ± 0.15 | 3.96 ± 0.19 | 3.95 ± 0.21 | 3.95 ± 0.13 |
| Eye Size (mm) | 0.352 ± 0.016 | 0.344 ± 0.014 | 0.351 ± 0.016 | 0.346 ± 0.023 | 0.351 ± 0.16 |
| Tel. Area (mm2) | 0.0306 ± .0028 | 0.0294 ± 0.0026 | 0.0292 ± 0.0026 | 0.0283 ± 0.0025 * | 0.0280 ± 0.0035 * |
| Dien. Area (mm2) | 0.117 ± 0.009 | 0.111 ± 0.006 | 0.112 ± 0.010 | 0.111 ± 0.011 | 0.113 ± 0.013 |
| Hind. Area (mm2) | 0.0499 ± 0.0060 | 0.0503 ± 0.0058 | 0.0495 ± 0.0057 | 0.0463 ± 0.0035 | 0.0494 ± 0.0049 |
| AO+ Tel AO+ Dien | 6.41 ± 0.97 5.27 ± 1.34 | 6.96 ± 1.09 5.68 ± 0.91 | 7.5 ± 0.87 9.00 ± 0.87 | 6.55 ± 0.83 7.57 ± 1.51 | 6.36 ± 0.73 6.64 ± 0.69 |
| Embryonic Treatment Condition | 0 mg/L Cocaine | 2.5 mg/L Cocaine | 5.0 mg/L Cocaine | 10.0 mg/L Cocaine | 20.0 mg/L Cocaine |
|---|---|---|---|---|---|
| Survival at 8–10 months | 61.5% ± 7.8 | 63.5% ± 7.4 | 62.7% ± 7.6 | 63.2% ± 13.2 | 66.8% ± 11.2 |
| Percentage of Males | 34.9% ± 14.1 | 50.3% ± 8.7 | 55.2% ± 11.0 | 43.8% ± 12.9 | 62.5% ± 9.2 |
| Body Len. (cm) | 3.6 ± 0.1 | 3.8 ± 0.3 | 3.7 ± 0.2 | 3.8 ± 0.2 | 3.6 ± 0.2 |
| Body Wt. (g) | 0.45 ± 0.07 | 0.44 ± 0.10 | 0.48 ± 0.10 | 0.43 ± 0.09 | 0.44 ± 0.13 |
| Tel. Area (mm2) | 1.86 ± 0.21 | 1.98 ± 0.22 | 1.97 ± 0.3 | 2.09 ± 0.27 | 2.11 ± 0.38 |
| Baseline HR | 138.3 ± 9.5 | 144.5 ± 8.0 | 132.9 ± 6.2 | 138.6 ± 6.5 | 130.5 ± 9.8 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mersereau, E.J.; Boyle, C.A.; Poitra, S.; Espinoza, A.; Seiler, J.; Longie, R.; Delvo, L.; Szarkowski, M.; Maliske, J.; Chalmers, S.; et al. Longitudinal Effects of Embryonic Exposure to Cocaine on Morphology, Cardiovascular Physiology, and Behavior in Zebrafish. Int. J. Mol. Sci. 2016, 17, 847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060847
Mersereau EJ, Boyle CA, Poitra S, Espinoza A, Seiler J, Longie R, Delvo L, Szarkowski M, Maliske J, Chalmers S, et al. Longitudinal Effects of Embryonic Exposure to Cocaine on Morphology, Cardiovascular Physiology, and Behavior in Zebrafish. International Journal of Molecular Sciences. 2016; 17(6):847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060847
Chicago/Turabian StyleMersereau, Eric J., Cody A. Boyle, Shelby Poitra, Ana Espinoza, Joclyn Seiler, Robert Longie, Lisa Delvo, Megan Szarkowski, Joshua Maliske, Sarah Chalmers, and et al. 2016. "Longitudinal Effects of Embryonic Exposure to Cocaine on Morphology, Cardiovascular Physiology, and Behavior in Zebrafish" International Journal of Molecular Sciences 17, no. 6: 847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060847