
Stable Toll-Like Receptor 10 Knockdown in THP-1 Cells Reduces TLR-Ligand-Induced Proinflammatory Cytokine Expression

Abstract
:1. Introduction
2. Results
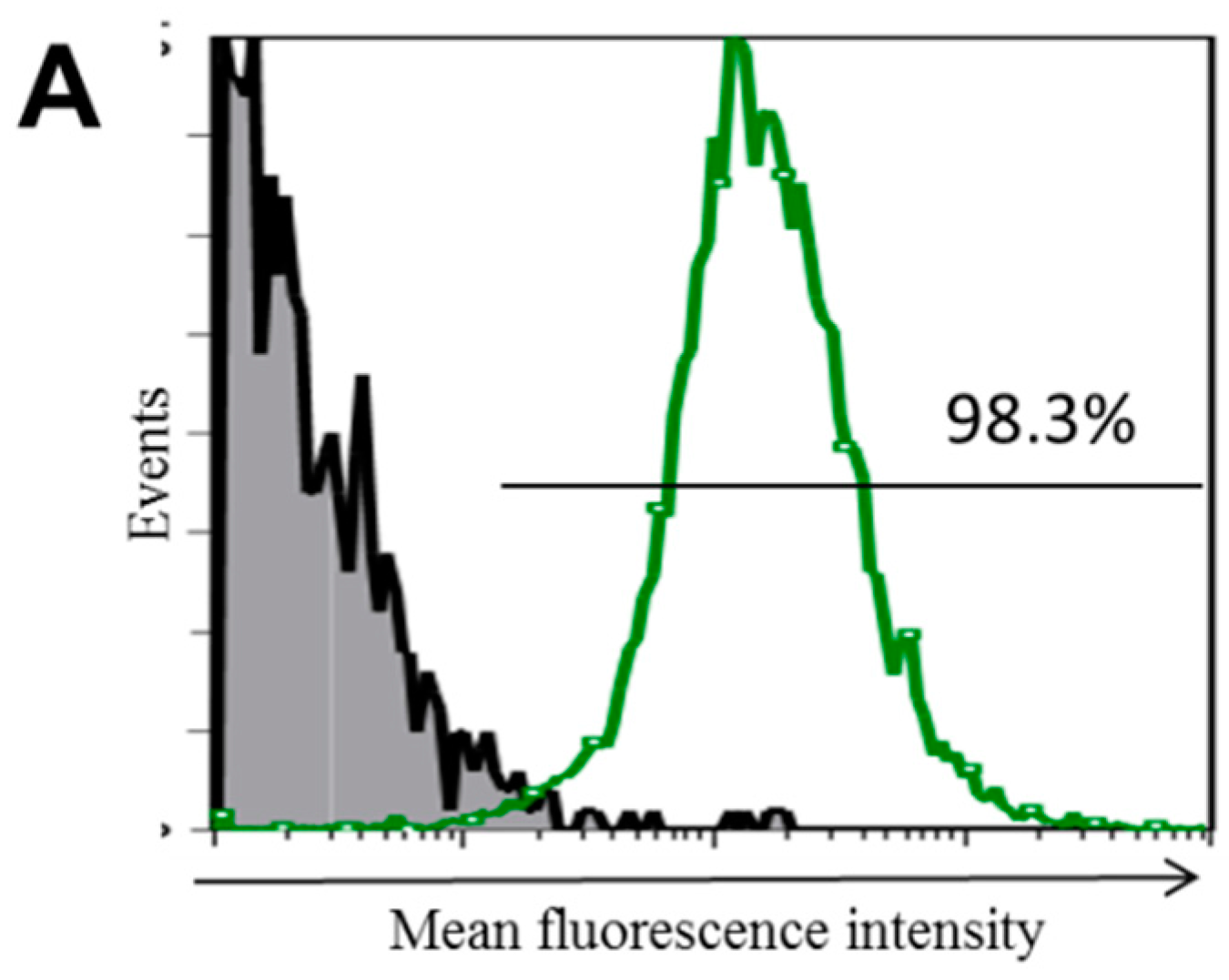
2.1. Toll-Like Receptor 10 (TLR10) Short Hairpin RNA (shRNA) Lentiviral Particle-Mediated TLR10 Knockdown in THP-1 Cells
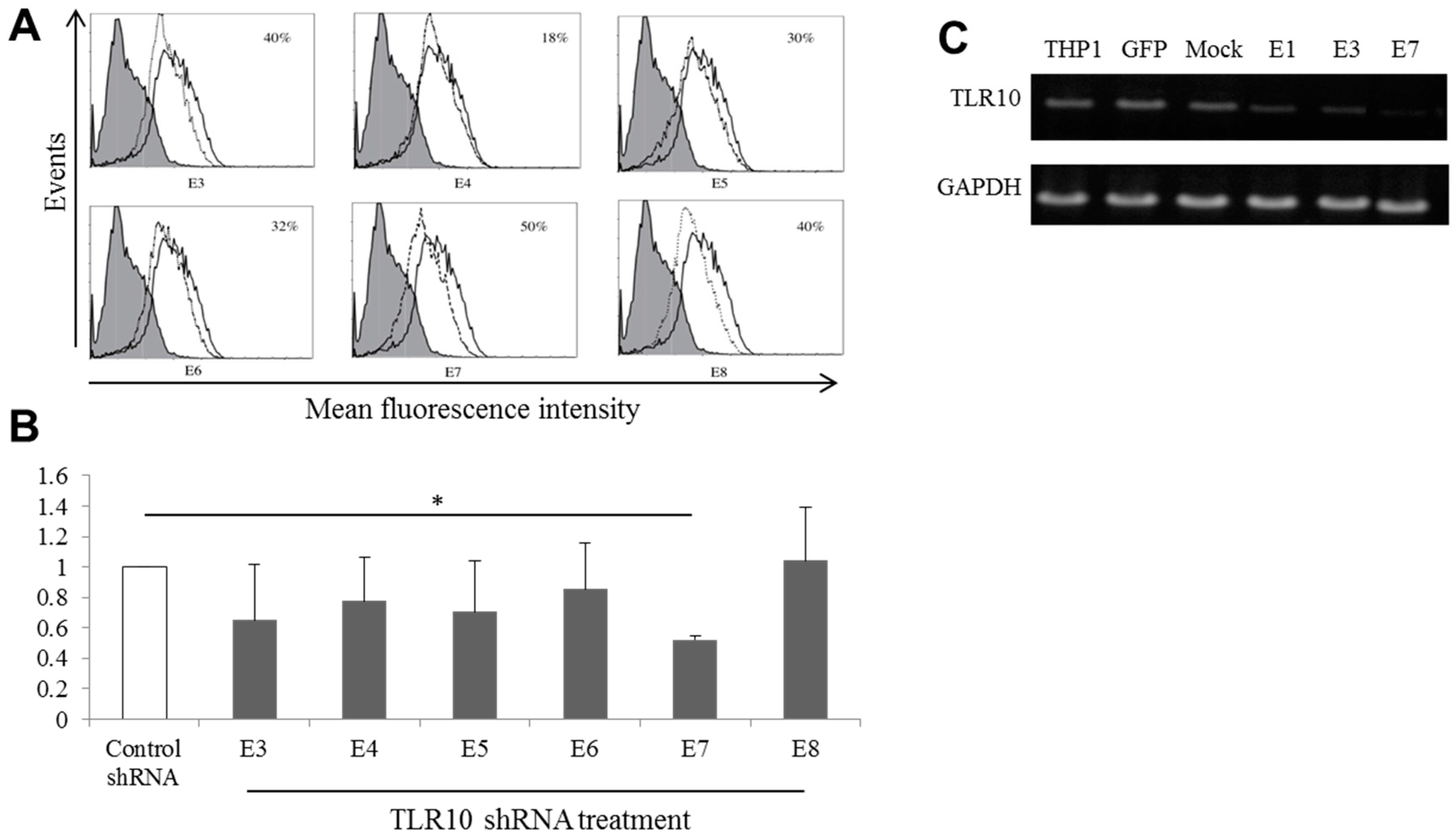
2.2. Generation of Stable TLR10 Knockdown THP-1 Clones
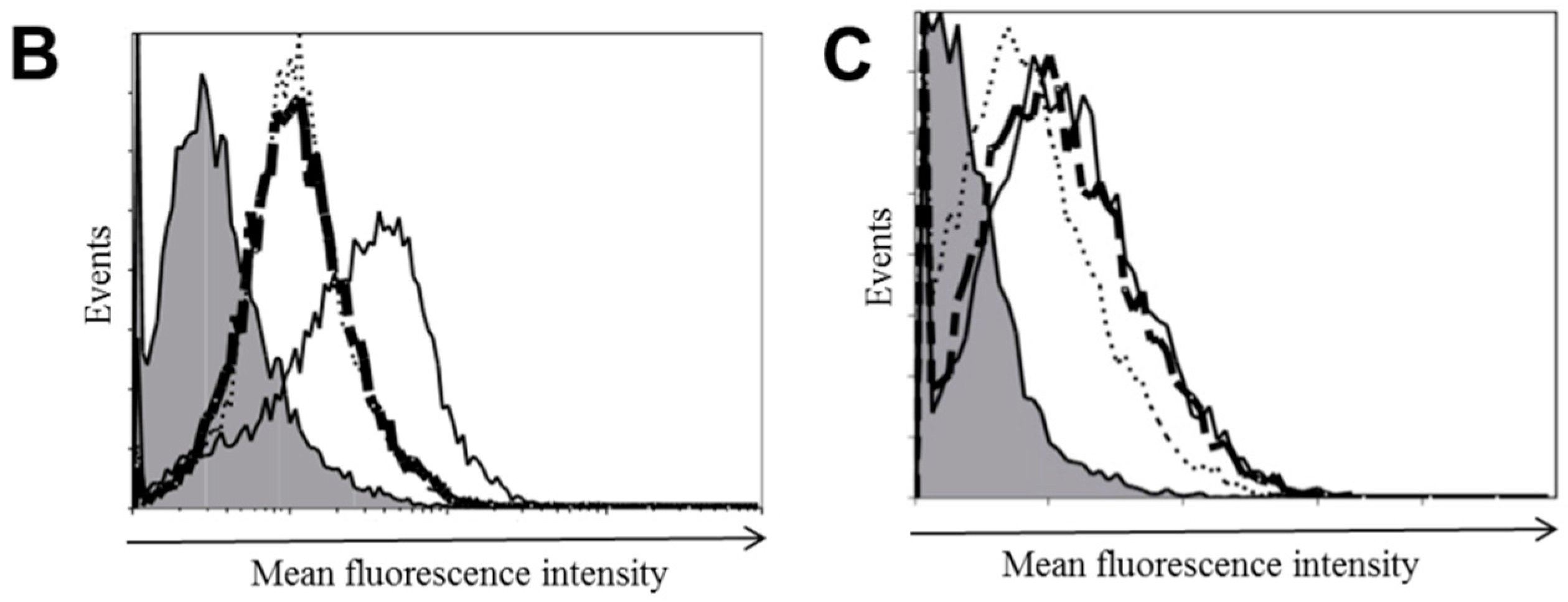
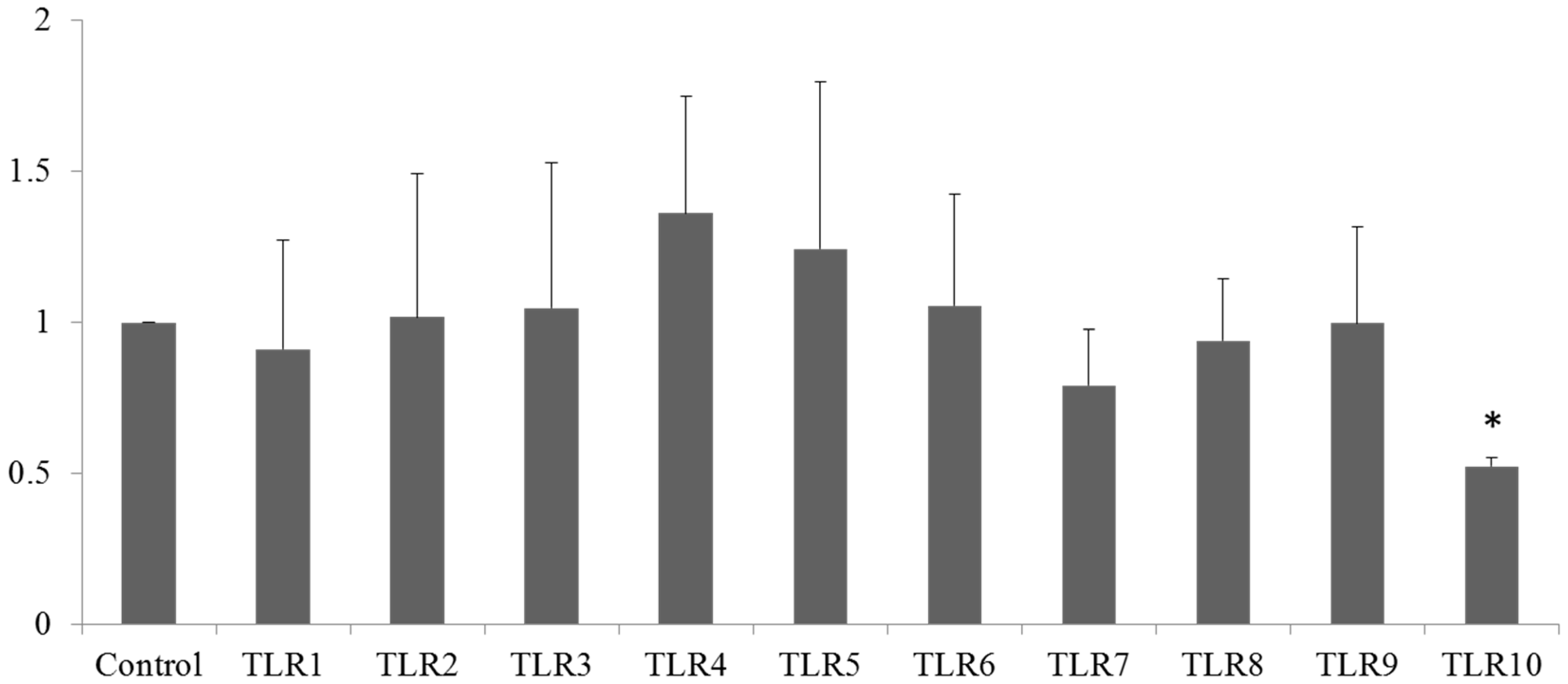
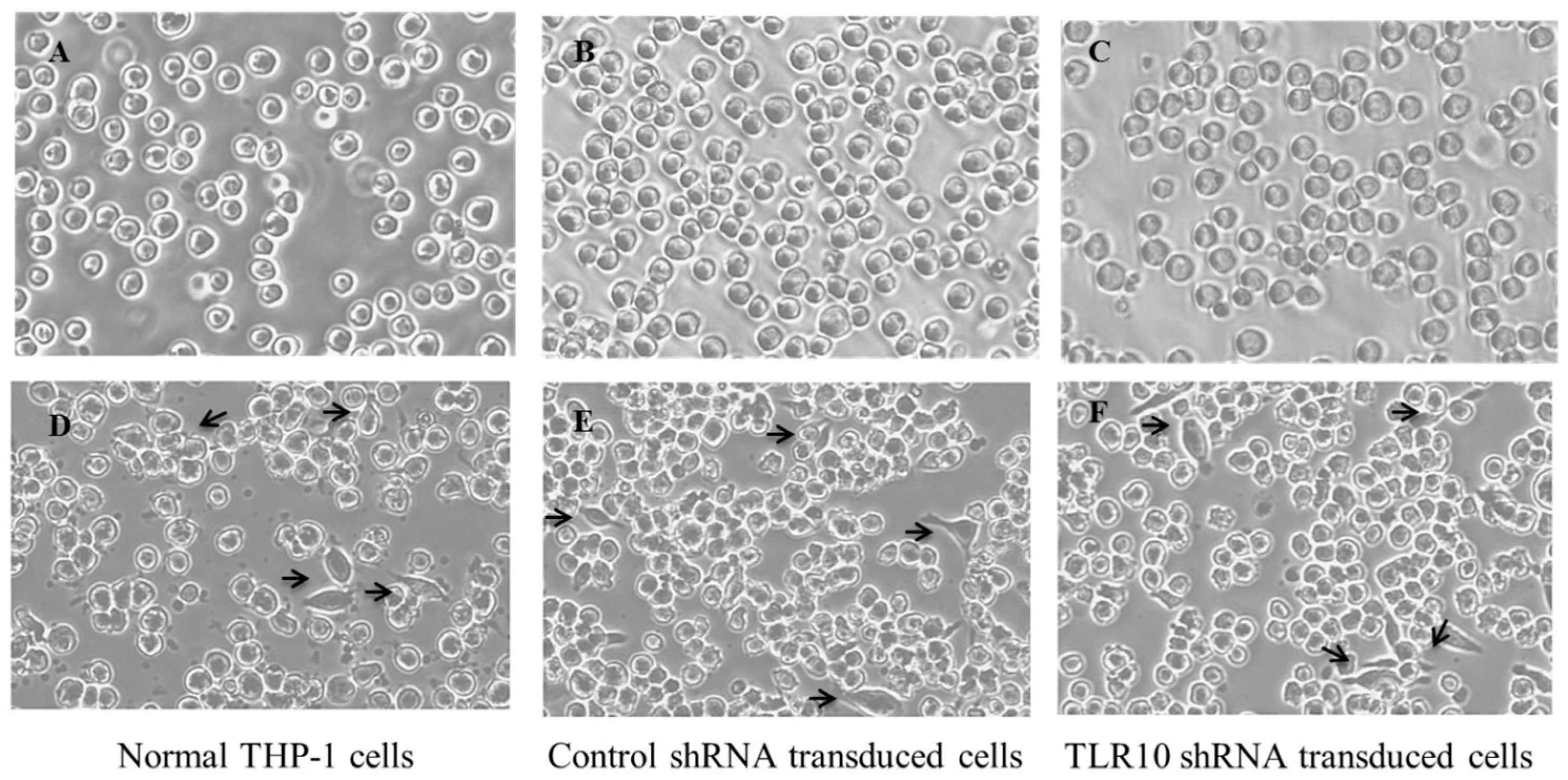
2.3. Stable Knockdown of TLR10 Does Not Affect the Expression of Other TLRs and Phorbol 12-Myristate 13-Acetate (PMA)-Induced Differentiation of THP-1 Cells
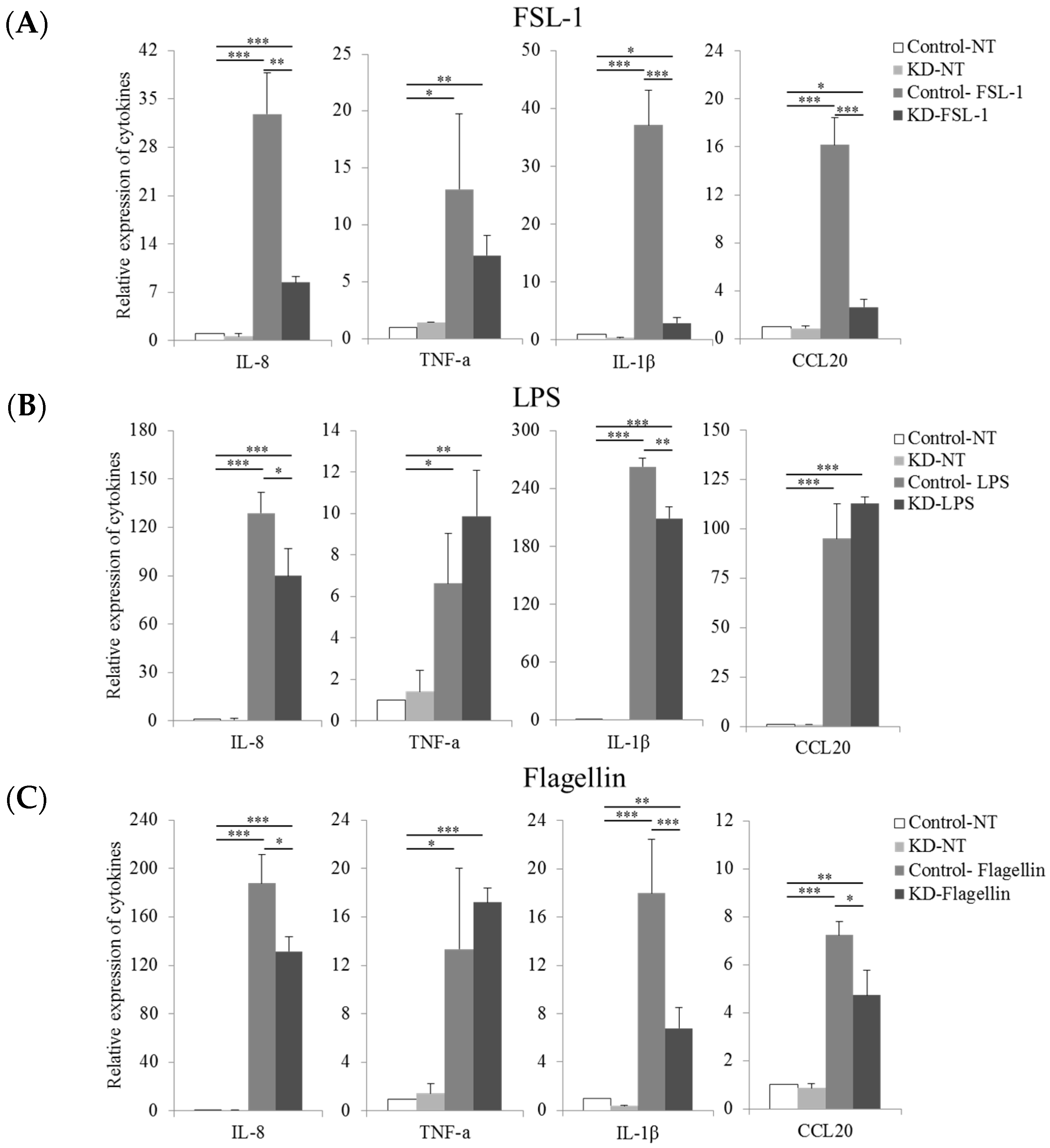
2.4. Knockdown of TLR10 Reduces TLR Ligand-Induced Pro-Inflammatory Cytokine Expression
2.5. Putative Binding Sites of the Nuclear Factor NF-κB, Activator Protein 1 (AP-1) and c-Jun on Promoters of Interleukin-8 (IL-8), Tumor Necrosis Factor-α (TNF-α), IL-1β, and Chemokine (C-C Motif) Ligand 20 (CCL20)
3. Discussion
4. Experimental Section
4.1. Reagents and Chemicals
4.2. Cell Culture and Diffrentiation
4.3. shRNA-Mediated Gene Silencing
4.4. Flow Cytometric Analysis
4.5. Quantitative Real-Time PCR and Semi-Quantitative PCR
4.6. Prediction of Transcription Factor Binding Sites
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guan, Y.; Ranoa, D.R.; Jiang, S.; Mutha, S.K.; Li, X.; Baudry, J.; Tapping, R.I. Human TLRs 10 and 1 share common mechanisms of innate immune sensing but not signaling. J. Immunol. 2010, 184, 5094–5103. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.; Ulevitch, R.J. Identification of hTLR10: A novel human Toll-like receptor preferentially expressed in immune cells. Biochim. Biophys. Acta 2001, 1518, 157–161. [Google Scholar] [CrossRef]
- Hasan, U.; Chaffois, C.; Gaillard, C.; Saulnier, V.; Merck, E.; Tancredi, S.; Guiet, C.; Briere, F.; Vlach, J.; Lebecque, S.; et al. Human TLR10 is a functional receptor, expressed by B cells and plasmacytoid dendritic cells, which activates gene transcription through MyD88. J. Immunol. 2005, 174, 2942–2950. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, R.G.; Manavalan, B.; Lee, G.; Choi, S. Molecular modeling-based evaluation of hTLR10 and identification of potential ligands in Toll-like receptor signaling. PLoS ONE 2010, 5, e12713. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kok, K.H.; Jaume, M.; Cheung, T.K.; Yip, T.F.; Lai, J.C.; Guan, Y.; Webster, R.G.; Jin, D.Y.; Peiris, J.S. Toll-like receptor 10 is involved in induction of innate immune responses to influenza virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 3793–3798. [Google Scholar] [CrossRef] [PubMed]
- Regan, T.; Nally, K.; Carmody, R.; Houston, A.; Shanahan, F.; Macsharry, J.; Brint, E. Identification of TLR10 as a key mediator of the inflammatory response to Listeria monocytogenes in intestinal epithelial cells and macrophages. J. Immunol. 2013, 191, 6084–6092. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, H.; Iwatani, S.; Cruz, M.; Jimenez Abreu, J.A.; Uchida, T.; Mahachai, V.; Vilaichone, R.K.; Graham, D.Y.; Yamaoka, Y. Toll-like Receptor 10 in Helicobacter pylori Infection. J. Infect. Dis. 2015, 212, 1666–1676. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tay, P.N.; Cao, W.; Li, W.; Lu, J. Integrin-Nucleated Toll-like receptor (TLR) dimerization reveals subcellular targeting of TLRs and distinct mechanisms of TLR4 activation and signaling. FEBS Lett. 2002, 532, 171–176. [Google Scholar] [CrossRef]
- Opsal, M.A.; Vage, D.I.; Hayes, B.; Berget, I.; Lien, S. Genomic organization and transcript profiling of the bovine toll-like receptor gene cluster TLR6-TLR1-TLR10. Gene 2006, 384, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, H.; Muneta, Y.; Suzuki, K.; Eguchi-Ogawa, T.; Awata, T.; Uenishi, H. Porcine Toll-like receptor 1, 6, and 10 genes: Complete sequencing of genomic region and expression analysis. Mol. Immunol. 2006, 43, 1474–1480. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.P.; Svingen, P.A.; Rahman, M.K.; Xiong, Y.; Faubion, W.A., Jr. FOXP3 regulates TLR10 expression in human T regulatory cells. J. Immunol. 2007, 179, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Oosting, M.; Cheng, S.C.; Bolscher, J.M.; Vestering-Stenger, R.; Plantinga, T.S.; Verschueren, I.C.; Arts, P.; Garritsen, A.; van Eenennaam, H.; Sturm, P.; et al. Human TLR10 is an anti-inflammatory pattern-recognition receptor. Proc. Natl. Acad. Sci. USA 2014, 111, E4478–E4484. [Google Scholar] [CrossRef] [PubMed]
- Krutzik, S.R.; Tan, B.; Li, H.; Ochoa, M.T.; Liu, P.T.; Sharfstein, S.E.; Graeber, T.G.; Sieling, P.A.; Liu, Y.J.; Rea, T.H.; et al. TLR activation triggers the rapid differentiation of monocytes into macrophages and dendritic cells. Nat. Med. 2005, 11, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Jung, J.H.; Kim, J.Y. 1,25-Dihydroxyvitamin D3 up-regulates TLR10 while down-regulating TLR2, 4, and 5 in human monocyte THP-1. J. Steroid Biochem. Mol. Biol. 2014, 141, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.S.T.; Ho, B.; Leung, B.P.; Ding, J.L. TLR cross-talk confers specificity to innate immunity. Int. Rev. Immunol. 2014, 33, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sidiropoulos, P.; Song, G.; Pagliari, L.J.; Birrer, M.J.; Stein, B.; Anrather, J.; Pope, R.M. TNF-α gene expression in macrophages: Regulation by NF-κB is independent of c-Jun or C/EBP β. J. Immunol. 2000, 164, 4277–4285. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, Y.J.; Koh, H.S.; Jang, T.Y.; Park, H.E.; Kim, J.Y. Reactive oxygen species enhance TLR10 expression in the human monocytic cell line THP-1. Int. J. Mol. Sci. 2010, 11, 3769–3782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, B.; Sumner, S.; Maitland, N.; Lewis, C.E. Macrophages in gene therapy: Cellular delivery vehicles and in vivo targets. J. Leukoc. Biol. 2002, 72, 417–428. [Google Scholar] [PubMed]
- Kusumawati, A.; Commes, T.; Liautard, J.P.; Widada, J.S. Transfection of myelomonocytic cell lines: Cellular response to a lipid-based reagent and electroporation. Anal. Biochem. 1999, 269, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, S.L.; Standiford, T.; Kasahara, K.; Strieter, R.M. Interleukin-8 (IL-8): The major neutrophil chemotactic factor in the lung. Exp. Lung Res. 1991, 17, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Hieshima, K.; Imai, T.; Baba, M.; Shoudai, K.; Ishizuka, K.; Nakagawa, T.; Tsuruta, J.; Takeya, M.; Sakaki, Y.; Takatsuki, K.; et al. A novel human CC chemokine PARC that is most homologous to macrophage-inflammatory protein-1 α/LD78 α and chemotactic for T lymphocytes, but not for monocytes. J. Immunol. 1997, 159, 1140–1149. [Google Scholar] [PubMed]
- Rider, P.; Carmi, Y.; Guttman, O.; Braiman, A.; Cohen, I.; Voronov, E.; White, M.R.; Dinarello, C.A.; Apte, R.N. IL-1α and IL-1β recruit different myeloid cells and promote different stages of sterile inflammation. J. Immunol. 2011, 187, 4835–4843. [Google Scholar] [CrossRef] [PubMed]
- Le, J.; Vilcek, J. Tumor necrosis factor and interleukin 1: Cytokines with multiple overlapping biological activities. Lab. Investig. 1987, 56, 234–248. [Google Scholar] [PubMed]
- Farre, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Rosello, L.; Alba, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [PubMed]
- ALGGEN. Available online: http://alggen.lsi.upc.es (accessed on 31 May 2016).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factors | IL-8 | IL-1β | CCL20 | TNF-α |
|---|---|---|---|---|
| NF-κB | nd | nd | a 38–49 72–81 154–165 | nd |
| NF-κB1 | 45–55 | nd | 62–72 71–81 197–207 | 191–201 |
| AP-1 | 66–74 | 18–26 234–241 | nd | nd |
| c-Jun | 11–17 68–74 112–118 243–249 | 18–24 34–40 236–242 | 101–107 166–172 | nd |
| Construct | 5′-Antisense (19 nt)-Hairpin Loop (9 nt)-Sense (19 nt)-Termination (5 Ts)-3′ |
|---|---|
| sc-40272-VA | CTCTGTGGGTGAAGAATGATTCAAGAGATCATTCTTCACCCACAGAGTTTTT |
| sc-40272-VB | GGAACCCATTCCATTCTATTTCAAGAGAATAGAATGGAATGGGTTCCTTTTT |
| sc-40272-VC | CCAGAGAAATGTATGAACTTTCAAGAGAAGTTCATACATTTCTCTGGTTTTT |
| Gene | Sequence (5′~3′) |
|---|---|
| TLR1-F | AGTGTTTTCAAATTCAACCAGGA |
| TLR1-R | AATGACAGGTCCAAGTGCTT |
| TLR2-F | CATTCTATGCCTGAAACTTGTCA |
| TLR2-R | CCAGTGTCTTGGGAATGCAG |
| TLR3-F | GTTGACTCAGGTACCCGATGA |
| TLR3-R | ATACCTTGTGAAGTTGGCGG |
| TLR4-F | CCATTGAAGAATTCCGATTAGCA |
| TLR4-R | CAATAGTCACACTCACCAGGG |
| TLR5-F | CTTCCTGCTCCTTTGATGGC |
| TLR5-R | TCCTGATATAGTTGAAGCTCAGC |
| TLR6-F | TCATGTTCCAAAAGACCTACCG |
| TLR6-R | ACTCTGATAGAAAGCTCATGTCA |
| TLR7-F | AACTCTGCCCTGTGATGTCA |
| TLR7-R | TGAGGTTCGTGGTGTTCGT |
| TLR8-F | TCATTTCAAGGGCTGCAAAATC |
| TLR8-R | CCCGTCTGTGATATTCAAGCC |
| TLR9-F | GCTGTTCCTGAAGTCTGTGC |
| TLR9-R | ATCATGGAGGTGGTGGATGC |
| TLR10-F | TCCAGAGCTGCCAGAAGAAA |
| TLR10-R | AAATCCAGTGTCGTTGTGGC |
| TNF-α-F | TGAGCACTGAAAGCATGATCC |
| TNF-α-R | GGAGAAGAGGCTGAGGAACA |
| IL-8-F | TGCCAAGGAGTGCTAAAGAAC |
| IL-8-R | TCCACTCTCAATCACTCTCAGT |
| IL-1β-F | GGGATAACGAGGCTTATGTGC |
| IL-1β-R | AGGTGGAGAGCTTTCAGTTCA |
| CCL20-F | ACTGTGGCTTTTCTGGAATGG |
| CCL20-R | AGCACTAAACCCTCCATGATG |
| β-actin-F | GGGACCTGACTGACTACCTC |
| β-actin-R | AGCTTCTCCTTAATGTCACGC |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, H.V.; Kim, J.Y. Stable Toll-Like Receptor 10 Knockdown in THP-1 Cells Reduces TLR-Ligand-Induced Proinflammatory Cytokine Expression. Int. J. Mol. Sci. 2016, 17, 859. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060859
Le HV, Kim JY. Stable Toll-Like Receptor 10 Knockdown in THP-1 Cells Reduces TLR-Ligand-Induced Proinflammatory Cytokine Expression. International Journal of Molecular Sciences. 2016; 17(6):859. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060859
Chicago/Turabian StyleLe, Hai Van, and Jae Young Kim. 2016. "Stable Toll-Like Receptor 10 Knockdown in THP-1 Cells Reduces TLR-Ligand-Induced Proinflammatory Cytokine Expression" International Journal of Molecular Sciences 17, no. 6: 859. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060859