
Hypoxia Potentiates Anabolic Effects of Exogenous Hyaluronic Acid in Rat Articular Cartilage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
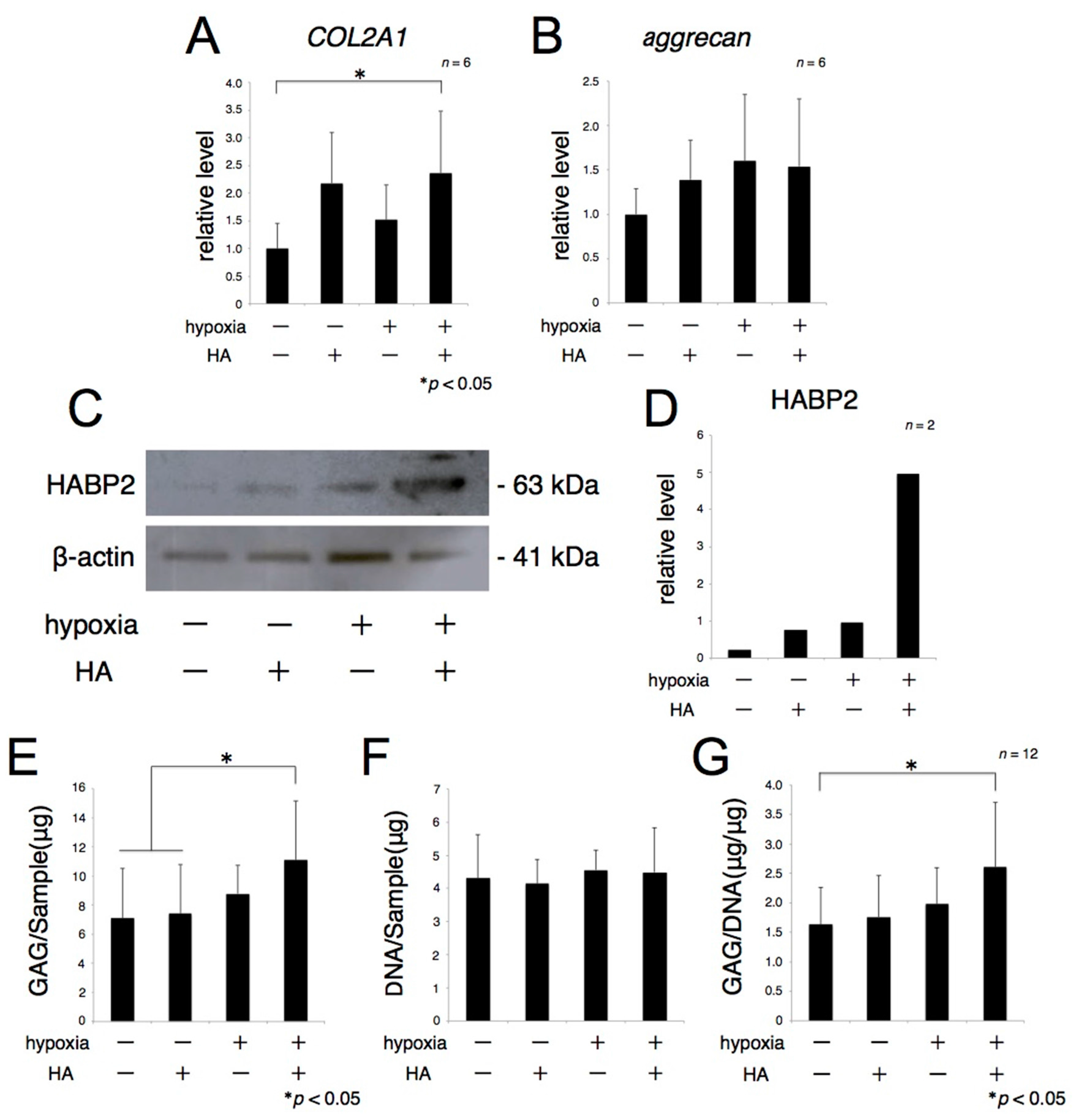
2.1. Hypoxia with Hyaluronic Acid (HA) Significantly Potentiates the Synthesis of Extracellular Matrix (ECM)
2.2. The Expression of Sex-Determining Region Y Box 9 (SOX9) Is Enhanced by Hypoxia with HA
2.3. Hypoxia with HA Promotes the Redifferentiation of Rat Articular Chondrocytes (rACs) and the Synthesis of Hyaline Cartilagenous ECM
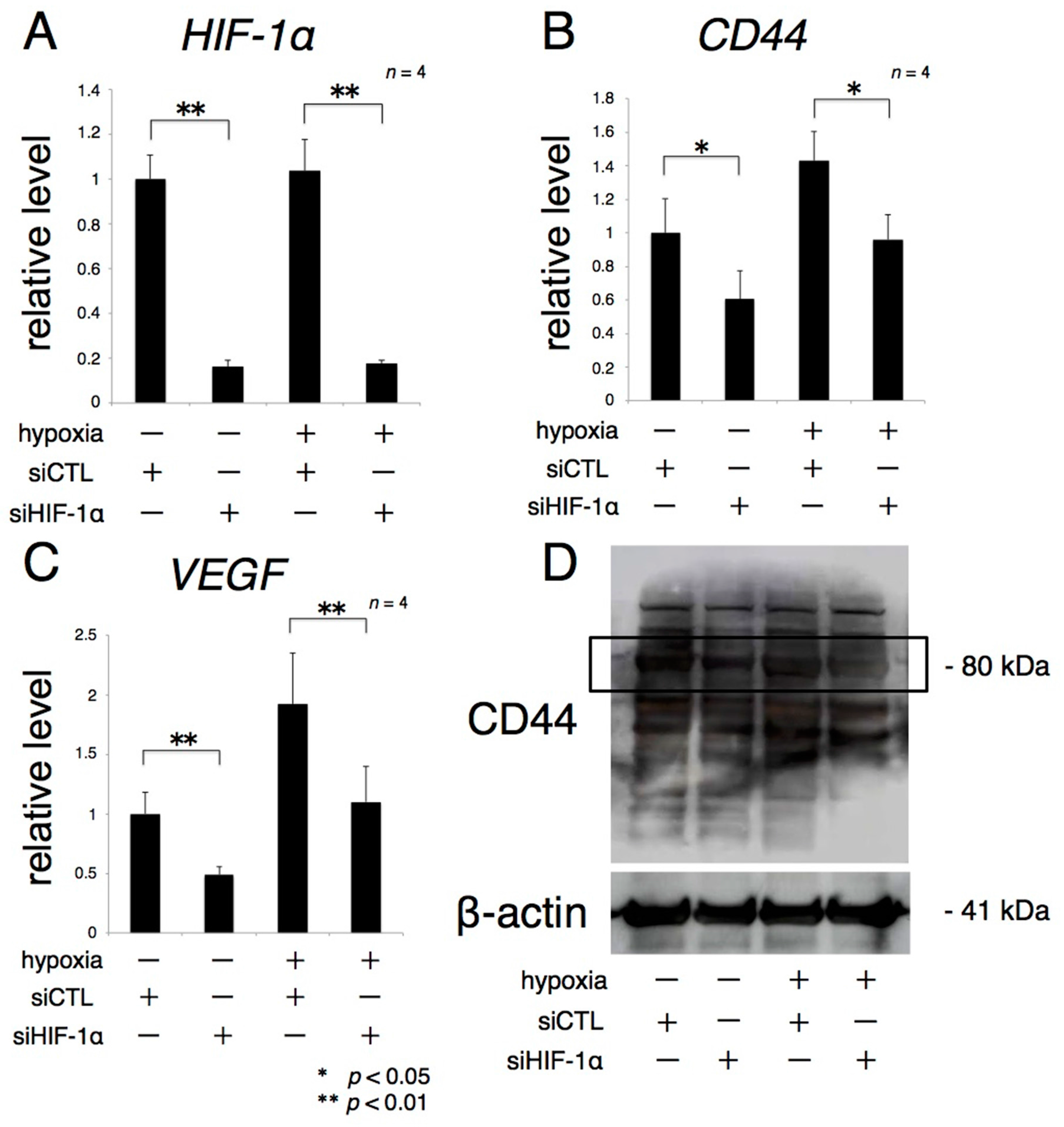
2.4. The Expression of CD44 Significantly Increases under Hypoxia
2.5. HIF-1α Controls CD44 Expression
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cartilage Samples
4.3. Isolation and Culture of Rat Articular Chondrocytes (rACs)
4.4. 3D Pellet Culture of rACs
4.5. Hyaluronic Acid Treatment and Hypoxia Experiments
4.6. Transfection with Small Interfering RNA (siRNA)
4.7. Total RNA Extraction and Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
4.8. Protein Extraction
4.9. Western Blot Analyses
4.10. Biochemical Analyses
4.11. Histological and Immunohistochemical Analyses
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Felson, D.T.; Zhang, Y. An update on the epidemiology of knee and hip osteoarthritis with a view to prevention. Arthritis Rheum. 1998, 41, 1343–1355. [Google Scholar] [CrossRef]
- Saito, T.; Fukai, A.; Mabuchi, A.; Ikeda, T.; Yano, F.; Ohba, S.; Nishida, N.; Akune, T.; Yoshimura, N.; Nakagawa, T.; et al. Transcriptional regulation of endochondral ossification by HIF-2α during skeletal growth and osteoarthritis development. Nat. Med. 2010, 16, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Muraki, S.; Akune, T.; En-Yo, Y.; Yoshida, M.; Suzuki, T.; Yoshida, H.; Ishibashi, H.; Tokimura, F.; Yamamoto, S.; Tanaka, S.; et al. Joint space narrowing, body mass index, and knee pain: The ROAD study (OAC1839R1). Osteoarthr. Cartil. 2015, 23, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Yorimitsu, M.; Nishida, K.; Shimizu, A.; Doi, H.; Miyazawa, S.; Komiyama, T.; Nasu, Y.; Yoshida, A.; Watanabe, S.; Ozaki, T. Intra-articular injection of interleukin-4 decreases nitric oxide production by chondrocytes and ameliorates subsequent destruction of cartilage in instability-induced osteoarthritis in rat knee joints. Osteoarthr. Cartil. 2008, 16, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Mero, A.; Campisi, M.; Favero, M.; Barbera, C.; Secchieri, C.; Dayer, J.M.; Goldring, M.B.; Goldring, S.R.; Pasut, G. A hyaluronic acid-salmon calcitonin conjugate for the local treatment of osteoarthritis: Chondro-protective effect in a rabbit model of early OA. J. Control. Release 2014, 187, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, Y.; Saito, T.; Sugita, S.; Hikata, T.; Kobayashi, H.; Fukai, A.; Taniguchi, Y.; Hirata, M.; Akiyama, H.; Chung, U.I.; et al. Notch signaling in chondrocytes modulates endochondral ossification and osteoarthritis development. Proc. Natl. Acad. Sci. USA 2013, 110, 1875–1880. [Google Scholar] [CrossRef] [PubMed]
- Dahl, L.B.; Dahl, I.M.; Engstrom-Laurent, A.; Granath, K. Concentration and molecular weight of sodium hyaluronate in synovial fluid from patients with rheumatoid arthritis and other arthropathies. Ann. Rheum. Dis. 1985, 44, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Balazs, E.A.; Denlinger, J.L. Viscosupplementation—A new concept in the treatment of osteoarthritis. J. Rheumatol. Suppl. 1993, 39, 3–9. [Google Scholar] [PubMed]
- Julovi, S.M.; Yasuda, T.; Shimizu, M.; Hiramitsu, T.; Nakamura, T. Inhibition of interleukin-1β-stimulated production of matrix metalloproteinases by hyaluronan via CD44 in human articular cartilage. Arthritis Rheum. 2004, 50, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Sliver, I.A. Measurement of pH and ionic composition of pericellular sites. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1975, 271, 261–272. [Google Scholar] [CrossRef]
- Svalastoga, E.; Kiaer, T. Oxygen consumption, diffusing capacity and blood flow of the synovial membrane in osteoarthritic rabbit knee joints. Acta Vet. Scand. 1989, 30, 121–125. [Google Scholar] [PubMed]
- Bruick, R.K.; McKnight, S.L. Oxygen sensing gets a second wind. Science 2002, 295, 807–808. [Google Scholar] [CrossRef] [PubMed]
- Schipani, E. Hypoxia and HIF-1α in chondrogenesis. Ann. N. Y. Acad. Sci. 2006, 1068, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Yudoh, K.; Nakamura, H.; Masuko-Hongo, K.; Kato, T.; Nishioka, K. Catabolic stress induces expression of hypoxia-inducible factor (HIF)-1 α in articular chondrocytes: Involvement of HIF-1 α in the pathogenesis of osteoarthritis. Arthritis Res. Ther. 2005, 7, R904–R914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubb, A.M.; Buffa, F.M.; Harris, A.L. Assessment of tumour hypoxia for prediction of response to therapy and cancer prognosis. J. Cell. Mol. Med. 2010, 14, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Knudson, C.B.; Thankamony, S.; Ariyoshi, W.; Mellor, L.; Im, H.J.; Knudson, W. Induction of CD44 cleavage in articular chondrocytes. Arthritis Rheum. 2010, 62, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Aibe, K.; Ryu, J.; Sano, S. Effects of hyaluronic acid on cartilage metabolism in free chondrocytes. J. Orthop. Sci. 1996, 1, 268–276. [Google Scholar] [CrossRef]
- Akmal, M.; Singh, A.; Anand, A.; Kesani, A.; Asiam, N.; Goodship, A.; Bently, G. The effects of hyaluronic acid on articular chondrocytes. J. Bone Jt. Surg. Br. 2005, 87, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Myllyharju, J.; Schipani, E. Extracellular matrix genes as hypoxia-inducible targets. Cell Tissue Res. 2010, 339, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Strobel, S.; Loparic, M.; Wendt, D.; Schenk, A.D.; Candrian, C.; Lindberg, R.L.; Moldovan, F.; Barbero, A.; Martin, I. Anabolic and catabolic responses of human articular chondrocytes to varying oxygen percentages. Arthritis Res. Ther. 2010, 12, R34. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, S.; Arai, Y.; Takahashi, K.A.; Kishida, T.; Terauchi, R.; Honjo, K.; Nakagawa, S.; Inoue, H.; Ikoma, K.; Ueshima, K.; et al. HIF-1α-induced HSP70 regulates anabolic responses in articular chondrocytes under hypoxic conditions. J. Orthop. Res. 2014, 32, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Markway, B.D.; Cho, H.; Johnstone, B. Hypoxia promotes redifferentiation and suppresses markers of hypertrophy and degeneration in both healthy and osteoarthritic chondrocytes. Arthritis Res. Ther. 2013, 15, R92. [Google Scholar] [CrossRef] [PubMed]
- Caron, M.M.; Emans, P.J.; Coolsen, M.M.; Voss, L.; Surtel, D.A.; Cremers, A.; van Rhijn, L.W.; Welting, T.J. Redifferentiation of dedifferentiated human articular chondrocytes: Comparison of 2D and 3D cultures. Osteoarthr. Cartil. 2012, 20, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Tallheden, T.; Karlsson, C.; Brunner, A.; van der Lee, J.; Hagg, R.; Tommasini, R.; Lindahl, A. Gene expression during redifferentiation of human articular chondrocytes. Osteoarthr. Cartil. 2004, 12, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Willers, C.; Xu, J.; Zheng, M.H. The chondrocyte: Biology and clinical application. Tissue Eng. 2006, 12, 1971–1984. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [PubMed]
- Akiyama, H.; Kim, J.E.; Nakashima, K.; Baimes, G.; Iwai, N.; Deng, J.M.; Zhang, Z.; Martin, J.F.; Behringer, R.R.; Nakamura, T.; et al. Osteo-chondroprogenitor cells are derived from Sox9 expressing precursors. Proc. Natl. Acad. Sci. USA 2005, 102, 14665–14670. [Google Scholar] [CrossRef] [PubMed]
- Amarilio, R.; Viukov, S.V.; Sharir, A.; Eshkar-Oren, I.; Johnson, R.S.; Zelzer, E. HIF1α regulation of Sox9 is necessary to maintain differentiation of hypoxic prechondrogenic cells during early skeletogenesis. Development 2007, 134, 3917–3928. [Google Scholar] [CrossRef] [PubMed]
- Turley, E.A. Purification of a hyluronate-binding protein fraction that modifies cell social behavior. Biochem. Biophys. Res. Commun. 1982, 108, 1016–1024. [Google Scholar] [CrossRef]
- Aruffo, A.; Stamenkovic, I.; Melnick, M.; Underhill, C.B.; Seed, B. CD44 is the principal cell surface receptor for hyaluronate. Cell 1990, 61, 1303–1313. [Google Scholar] [CrossRef]
- McCourt, P.A.; Ek, B.; Forsberg, N.; Gustafson, S. Intercellular adhesion molecule-1 is a cell surface receptor for hyaluronan. J. Biol. Chem. 1994, 269, 30081–30084. [Google Scholar] [PubMed]
- Turley, E.A.; Noble, P.W.; Bourguignon, L.Y. Signaling properties of hyaluronan receptors. J. Biol. Chem. 2002, 277, 4589–4592. [Google Scholar] [CrossRef] [PubMed]
- Krishnamachary, B.; Penet, M.F.; Nimmagadda, S.; Mironchik, Y.; Raman, V.; Solaiyappan, M.; Semenza, G.L.; Pomper, M.G.; Bhujwalla, Z.M. Hypoxia regulates CD44 and its variant isoforms through HIF-1α in triple negative breast cancer. PLoS ONE 2012, 7, e44078. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Imai, S.; Isoya, E.; Kubo, M.; Mimura, T.; Shioji, S.; Ueyama, H.; Matsusue, Y. Effect of dynamic compressive loading and its combination with a growth factor on the chondrocytic phenotype of 3-dimensional scaffold-embedded chondrocytes. Acta Orthop. 2009, 80, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Nakayama, K.; Akieda, S.; Iwamoto, Y. Simultaneous regeneration of full-thickness cartilage and subchondral bone defects in vivo using a three-dimensional scaffold-free autologous construct derived from high-density bone marrow-derived mesenchymal stem cells. J. Orthop. Surg. Res. 2014, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Farndale, R.W.; Buttle, D.J.; Barrett, A.J. Improved quantitation and discrimination of sulphated glycosaminoglycans by use of dimethylmethylene blue. Biochim. Biophys. Acta 1986, 883, 173–177. [Google Scholar] [CrossRef]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ichimaru, S.; Nakagawa, S.; Arai, Y.; Kishida, T.; Shin-Ya, M.; Honjo, K.; Tsuchida, S.; Inoue, H.; Fujiwara, H.; Shimomura, S.; et al. Hypoxia Potentiates Anabolic Effects of Exogenous Hyaluronic Acid in Rat Articular Cartilage. Int. J. Mol. Sci. 2016, 17, 1013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071013
Ichimaru S, Nakagawa S, Arai Y, Kishida T, Shin-Ya M, Honjo K, Tsuchida S, Inoue H, Fujiwara H, Shimomura S, et al. Hypoxia Potentiates Anabolic Effects of Exogenous Hyaluronic Acid in Rat Articular Cartilage. International Journal of Molecular Sciences. 2016; 17(7):1013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071013
Chicago/Turabian StyleIchimaru, Shohei, Shuji Nakagawa, Yuji Arai, Tsunao Kishida, Masaharu Shin-Ya, Kuniaki Honjo, Shinji Tsuchida, Hiroaki Inoue, Hiroyoshi Fujiwara, Seiji Shimomura, and et al. 2016. "Hypoxia Potentiates Anabolic Effects of Exogenous Hyaluronic Acid in Rat Articular Cartilage" International Journal of Molecular Sciences 17, no. 7: 1013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071013