
Digital Gene Expression Analysis of Ponkan Mandarin (Citrus reticulata Blanco) in Response to Asia Citrus Psyllid-Vectored Huanglongbing Infection

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Development of Citrus Huanglongbing (HLB) Symptoms on Experimental Trees
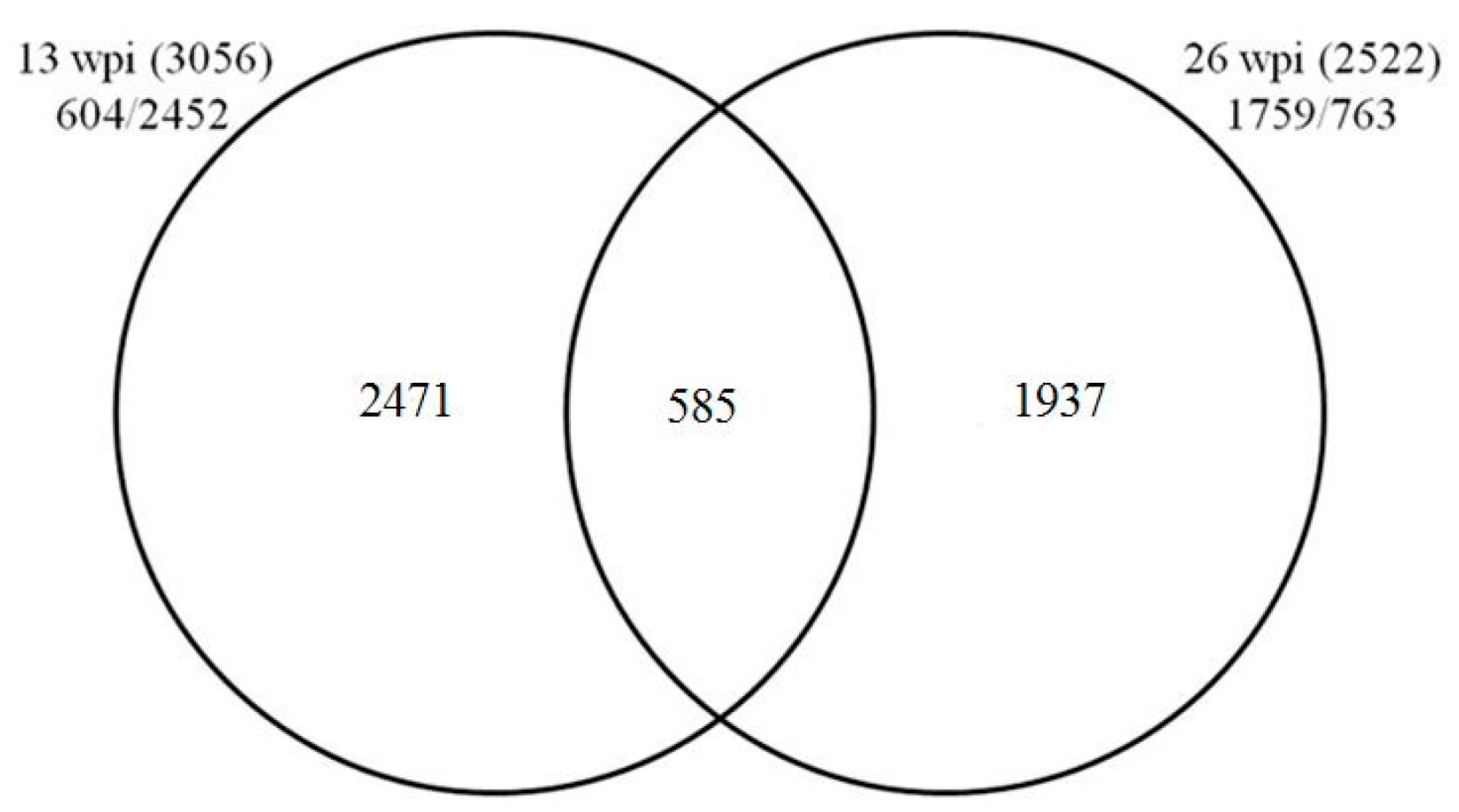
2.2. Digital Gene Expression (DGE) Profiling Result
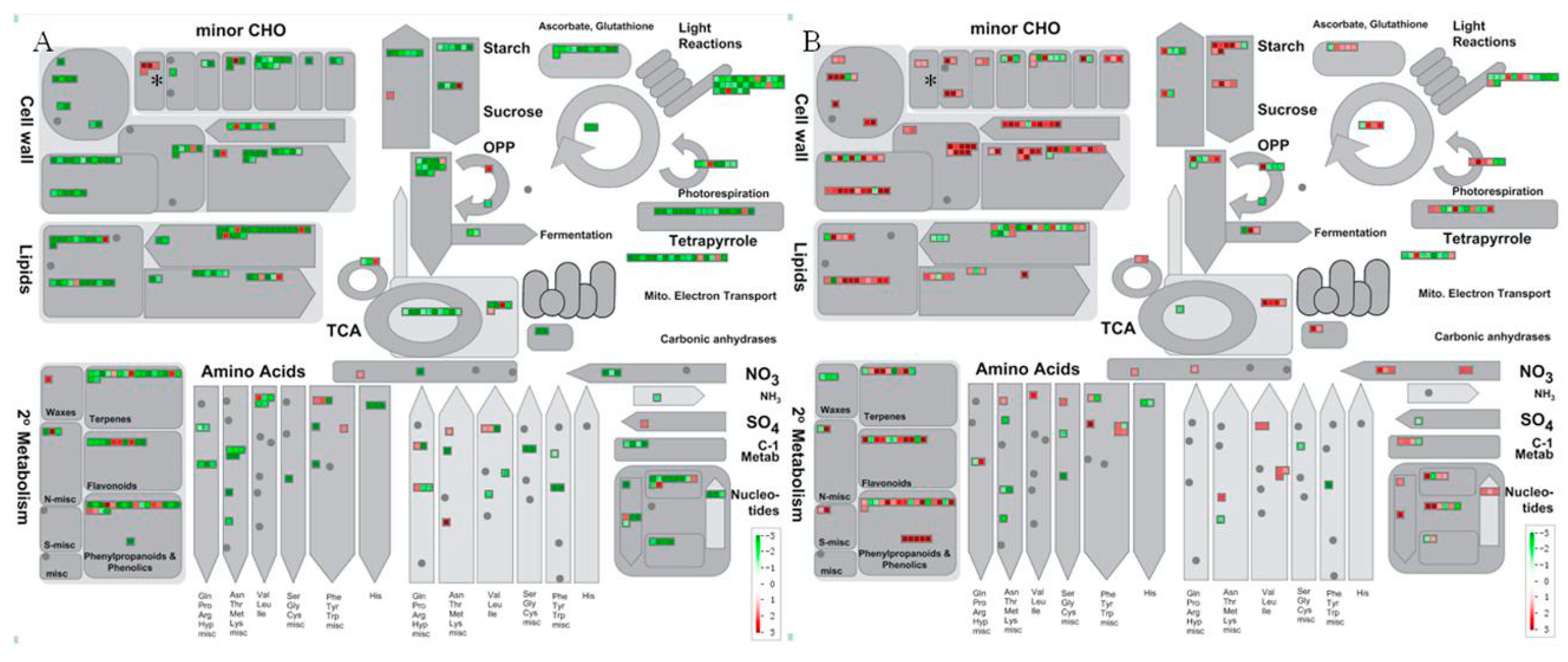
2.3. Functional Analysis of DGE Profiling Data
2.4. Gene Pathway Enrichment Analysis of HLB-Modulated Pathways
2.5. Carbohydrate Metabolism Was Significantly Regulated by HLB Infection
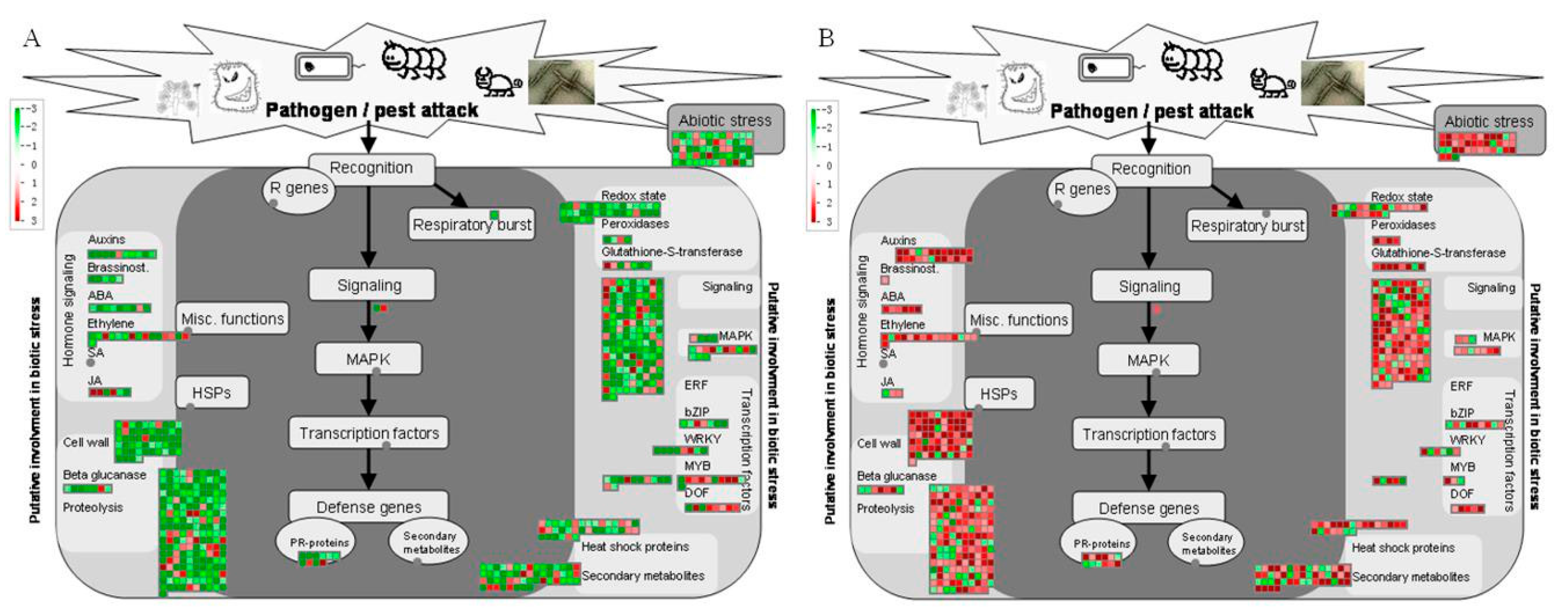
2.6. Stressresponse-Related Genes
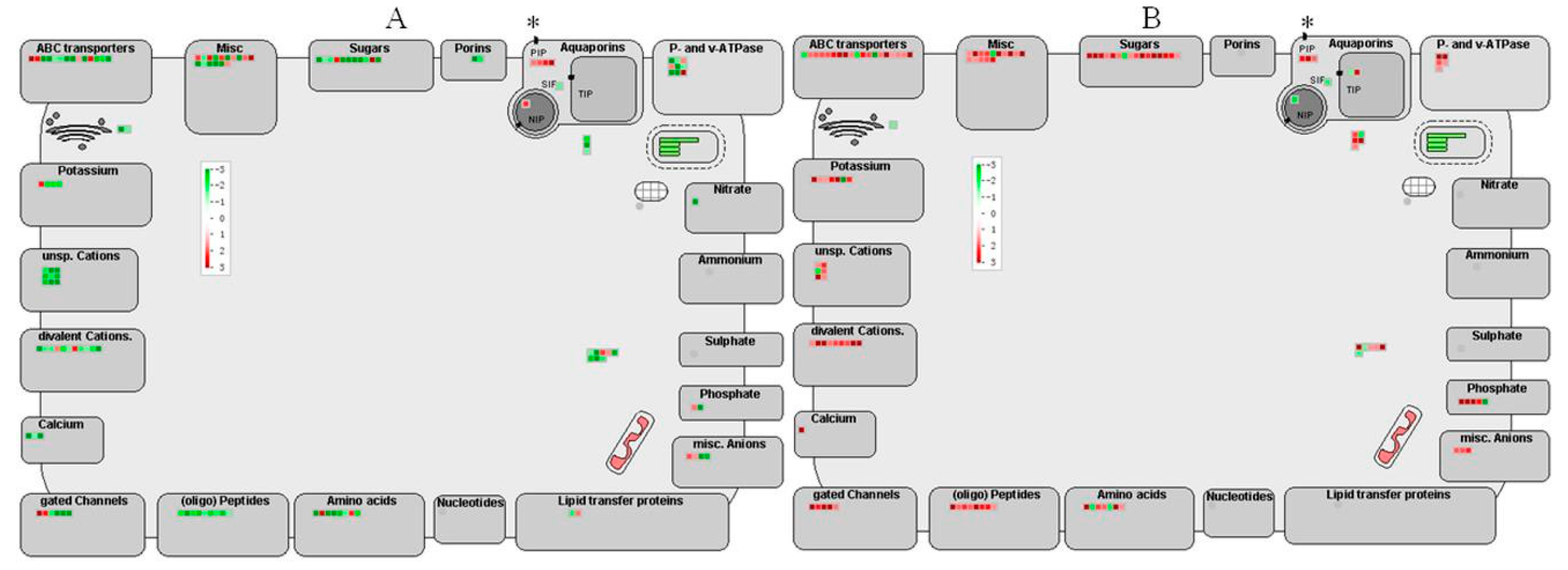
2.7. Transport Related Genes Regulated by HLB Infection
2.8. Cell Organization Related Genes
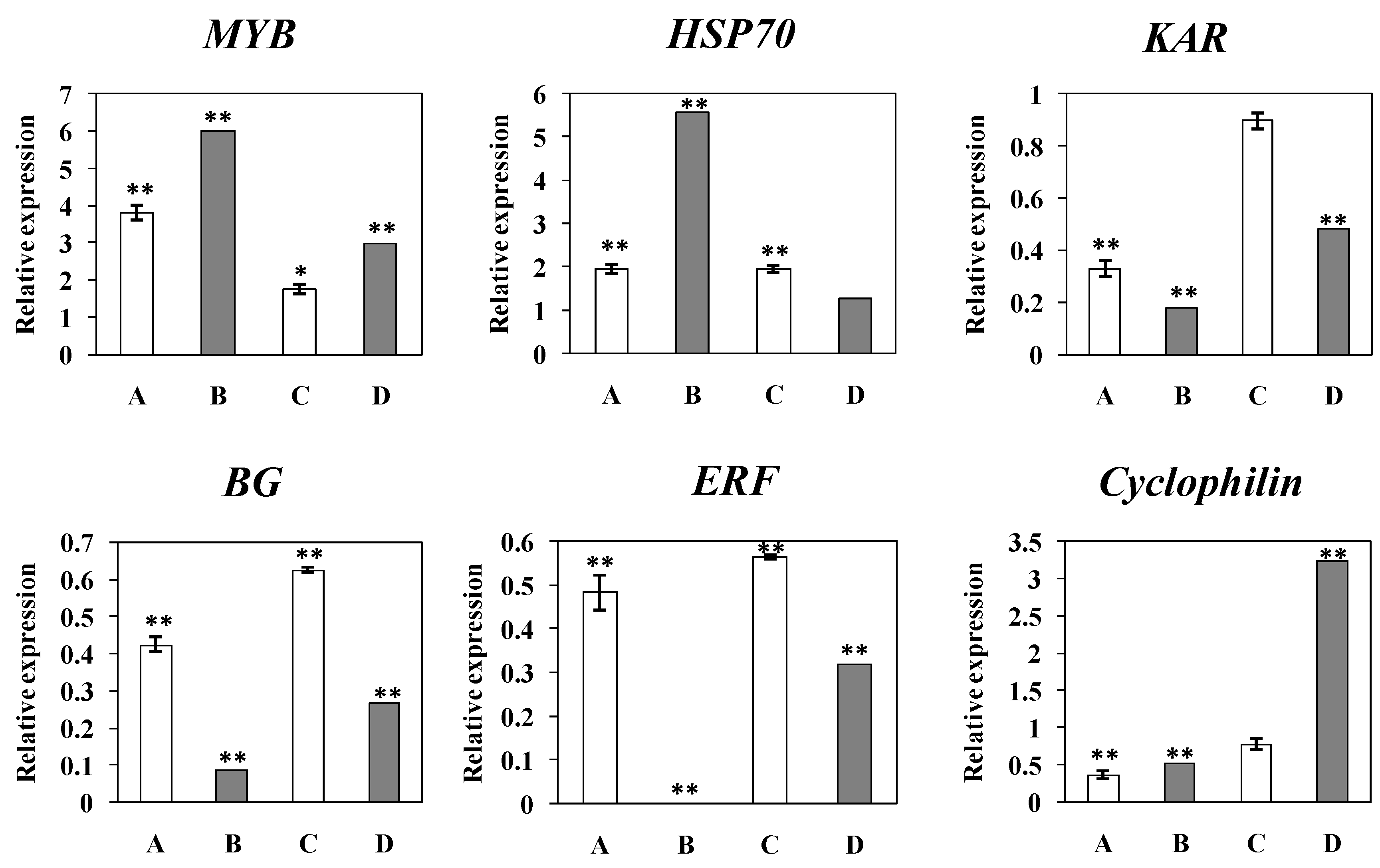
2.9. Quantitative Real Time PCR (qRT-PCR) Result
3. Discussion
Transport Related Genes Regulated by HLB Infection
4. Materials and Methods
4.1. Asian Citrus Psyllids Collection and Feeding
4.2. HLB Inoculation and Detection
4.3. RNA Preparation and Digital Gene Expression Profiling
4.4. Differential Gene Expression Analysis
4.5. Quantitative Real Time PCR (qRT-PCR) Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HLB | Citrus Huanglongbing |
| WPI | weeks post inoculation |
| DGE | digital gene expression |
| ACP | Asian citrus psyllids |
| DEG | Differentially expressed gene |
| CLas | Candidatus L. asiaticus |
| Clam | Candidatus L. americanus |
| CLaf | Candidatus L. africanus |
| SPS | sucrose-phosphate synthase |
| SPP1 | sucrose-phosphatase 1 |
| LHY | late elongated hypocotyls |
| EPR1 | early-phytochrome responsive 1 |
| GGPS1 | geranylgeranyl pyrophosphate synthase 1 |
| HPT1 | homogentisate phytyl transferase 1 |
| IRX12 | irregular xylem 12 |
| LAC | laccase |
| RLK | receptor like kinases |
| HSP | heat shock protein |
| PIP | plasma membrane intrinsic protein |
| PP | phloem protein |
| UPS | ubiquitin/proteasome system |
References
- Lin, K.H. Etiological studies of yellow shoot of citrus. Acta Phytopathol. Sin. 1956, 2, 13–42. [Google Scholar]
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Dutt, M.; Barthe, G.; Irey, M.; Grosser, J. Transgenic citrus expressing an Arabidopsis NPR1 gene exhibit enhanced resistance against Huanglongbing (HLB; Citrus Greening). PLoS ONE 2015, 10, e0137134. [Google Scholar] [CrossRef] [PubMed]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greending disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar]
- Mafra, V.; Martins, P.K.; Francisco, C.S.; Ribeiro-alves, M.; Freitas-astúa, J.; Machado, M.A. Candidatus Liberibacter americanus induces significant reprogramming of the transcriptome of the susceptible citrus genotype. BMC Genom. 2013, 14, 247. [Google Scholar] [CrossRef] [PubMed]
- Masaoka, Y.; Pustika, A.; Subandiyah, S.; Okada, A.; Hanundin, E.; Purwanto, B.; Okuda, M.; Okada, Y.; Saito, A.; Holford, P.; et al. Lower concentrations of microelements in leaves of citrus infected with “Candidatus Liberibacter asiaticus”. Jpn. Agric. Res. Q. 2011, 45, 269–275. [Google Scholar] [CrossRef]
- Folimonova, S.Y.; Achor, D.S. Early events of citrus greening (Huanglongbing) disease development at the ultrastructural level. Phytopathology 2010, 100, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Sagaram, U.S.; Burns, J.K.; Li, J.; Wang, N. Response of sweet orange (Citrus sinensis) to “Candidatus Liberibacter asiaticus” infection: Microscopy and microarray analyses. Phytopathology 2009, 99, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, E.; Gonzalez, P.; Achor, D.; Albrigo, G. Anatomical distribution of abnormally high levels of starch in HLB-affected Valencia orange trees. Physiol. Mol. Plant Pathol. 2009, 74, 76–83. [Google Scholar] [CrossRef]
- Achor, D.S.; Etxeberria, E.; Wang, N.; Folimonova, S.Y.; Chung, K.R.; Albrigo, L.G. Sequence of anatomical symptom observations in citrus affected with Huanglongbing disease. Plant Pathol. J. 2010, 9, 56–64. [Google Scholar] [CrossRef]
- Fan, J.; Chen, C.; Brlansky, R.H.; Gmitter, F.G., Jr.; Li, Z.-G. Changes in carbohydrate metabolism in Citrus sinensis infected with “Candidatus Liberibacter asiaticus”. Plant Pathol. 2010, 59, 1037–1043. [Google Scholar] [CrossRef]
- Donnell, P.J.O.; Schmelz, E.A.; Moussatche, P.; Lund, S.T.; Jones, J.B.; Klee, H.J. Susceptible to intolerance—Arange of hormonal actions in a susceptible Arabidopsis pathogen response. Plant J. 2003, 33, 245–257. [Google Scholar] [CrossRef]
- Martinelli, F.; Reagan, R.L.; Uratsu, S.L.; Phu, M.L.; Albrecht, U.; Zhao, W.; Davis, C.E.; Bowman, K.D.; Dandekar, A.M. Gene regulatory networks elucidating Huanglongbing disease mechanisms. PLoS ONE 2013, 8, e74256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albrecht, U.; Bowman, K. Gene expression in Citrus sinensis (L.) Osbeck following infection with the bacterial pathogen Candidatus Liberibacter asiaticus causing Huanglongbing in Florida. Plant Sci. 2008, 175, 291–306. [Google Scholar] [CrossRef]
- Albrecht, U.; Bowman, K.D. Transcriptional response of susceptible and tolerant citrus to infection with Candidatus Liberibacter asiaticus. Plant Sci. 2012, 185–186, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Chen, C.; Yu, Q.; Brlansky, R.H.; Li, Z.-G.; Gmitter, F.G. Comparative iTRAQ proteome and transcriptome analyses of sweet orange infected by “Candidatus Liberibacter asiaticus”. Physiol. Plant. 2011, 143, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Chen, C.; Yu, Q.; Khalaf, A.; Achor, D.S.; Brlansky, R.H.; Moore, G.A.; Li, Z.; Gmitter, F.G. Comparative transcriptional and anatomical analyses of tolerant rough lemon and susceptible sweet orange in response to “Candidatus Liberibacter asiaticus” infection. Mol. Plant Microbe Interact. 2012, 25, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Burns, J.K. Gene expression in Citrus sinensis fruit tissues harvested from Huanglongbing-infected trees: Comparison with girdled fruit. J. Exp. Bot. 2012, 63, 3307–3319. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, F.; Uratsu, S.L.; Albrecht, U.; Reagan, R.L.; Phu, M.L.; Britton, M.; Buffalo, V.; Fass, J.; Leicht, E.; Zhao, W.; et al. Transcriptome profiling of citrus fruit response to Huanglongbing disease. PLoS ONE 2012, 7, e38039. [Google Scholar] [CrossRef] [PubMed]
- Aritua, V.; Achor, D.; Gmitter, F.G.; Albrigo, G.; Wang, N. Transcriptional and microscopic analyses of citrus stem and root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 2013, 8, e73742. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Cheng, C.-Z.; Jiang, N.-H.; Jiang, B.; Zhang, Y.-Y.; Wu, B.; Hu, M.-L.; Zeng, J.-W.; Yan, H.-X.; Yi, G.-J.; et al. Comparative transcriptome and iTRAQ proteome analyses of citrus root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 2015, 10, e0126973. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhao, Y. Transcriptome comparison and gene coexpression network analysis provide a systems view of citrus response to “Candidatus Liberibacter asiaticus” infection. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Rawat, N.; Kiran, S.P.; Du, D.; Gmitter, F.G.; Deng, Z. Comprehensive meta-analysis, co-expression, and miRNA nested network analysis identifies gene candidates in citrus against Huanglongbing disease. BMC Plant Biol. 2015, 15, 184. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Rawat, N.; Deng, Z.; Gmitter, F.G. Construction of citrus gene coexpression networks from microarray data using random matrix theory. Horticult. Res. 2015, 2, e15026. [Google Scholar] [CrossRef]
- Duan, Y.; Zhou, L.; Hall, D.G.; Li, W.; Doddapaneni, H.; Lin, H.; Liu, L.; Vahling, C.M.; Gabriel, D.W.; Williams, K.P.; et al. Complete genome sequence of citrus Huanglongbing bacterium, “Candidatus Liberibacter asiaticus” obtained through metagenomics. Mol. Plant Microbe Interact. 2009, 22, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Hui, W.; Ding, F.; Zhong, Y.; Jiang, B.; Yi, G.; Wang, G. Comparison of different methods to transmit Candidatus liberibacter asiaticusto Murraya paniculata. J. Fruit Sci. 2011, 2, 268–272. [Google Scholar]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2010, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Translocation in the phloem. In Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2010; pp. 271–304. [Google Scholar]
- Salzman, R.A.; Tikhonova, I.; Bordelon, B.P.; Hasegawa, P.M.; Bressan, R.A. Coordinate accumulation of antifungal proteins and hexoses constitutes a developmentally controlled defense response during fruit ripening in grape. Plant Physiol. 1998, 117, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Herbers, K.; Takahata, Y.; Melzer, M.; Mock, H.P.; Hajirezaei, M.; Sonnewald, U. Regulation of carbohydrate partitioning during the interaction. Mol. Plant Pathol. 2000, 1, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ballicora, M.A.; Iglesias, A.A.; Preiss, J. ADP-glucose pyrophosphorylase, a regulatory enzyme for bacterial glycogen synthesis. Microbiol. Mol. Biol. Rev. 2003, 67, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.; Zhou, L.; Williams, D.S. Callose deposition in the phloem plasmodesmata and inhibition of phloem transport in citrus leaves infected with “Candidatus Liberibacter asiaticus”. Protoplasma 2010, 249, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Botha, C.E.J.; Cross, R.H.M.; van Bel, A.J.E.; Peter, C.I. Phloem loading in the sucrose-export-defective (SXD-1) mutant maize is limited by callose deposition at plasmodesmata in bundle sheath–vascular parenchyma interface. Protoplasma 2000, 214, 65–72. [Google Scholar] [CrossRef]
- Dinant, S.; Clark, A.M.; Zhu, Y.; Palauqui, J.; Kusiak, C.; Thompson, G.A. Diversity of the superfamily of phloem lectins (phloem protein 2) in angiosperms. Plant Physiol. 2003, 131, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Wightman, R.; Zhang, Z.; Gomez, L.D.; Atanassov, I.; Bukowski, J.-P.; Tryfona, T.; McQueen-Mason, S.J.; Dupree, P.; Turner, S. Arabidopsis genes IRREGULAR XYLEM (IRX15) and IRX15L encode DUF579-containing proteins that are essential for normal xylan deposition in the secondary cell wall. Plant J. Cell Mol. Biol. 2011, 66, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Chiniquy, D.; Varanasi, P.; Oh, T.; Harholt, J.; Katnelson, J.; Singh, S.; Auer, M.; Simmons, B.; Adams, P.D.; Scheller, H.V.; et al. Three novel rice genes closely related to the Arabidopsis IRX9, IRX9L, and IRX14 genes and their roles in xylan biosynthesis. Front. Plant Sci. 2013, 4, 83. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Staples, R.C. Laccase: New functions for an old enzyme. Phytochemistry 2002, 60, 551–565. [Google Scholar] [CrossRef]
- Nomura, K.; Melotto, M.; He, S. Suppression of host defense in compatible plant–Pseudomonas syringae interactions. Curr. Opin. Plant Biol. 2005, 8, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, N. Ultrastructural and cytochemical aspects of the response of eggplant parenchyma cells in direct contact with Verticillium-infected xylem vessels. Physiol. Mol. Plant Pathol. 1995, 46, 321–338. [Google Scholar] [CrossRef]
- Nagaoka, S.; Takano, T. Salt tolerance-related protein STO binds to a MYB transcription factor homologue and confers salt tolerance in Arabidopsis. J. Exp. Bot. 2003, 54, 2231–2237. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, R.; Ramsay, N.; Samach, A.; Corden, S.; Putterill, J.; Carre, I.A.; Centre, J.I.; Lane, C. The late elongated hypocotyl mutation of Arabidopsis disrupts circadian rhythms and the photoperiodic control of flowering. Cell 1998, 93, 1219–1229. [Google Scholar] [CrossRef]
- Kuno, N.; Møller, S.G.; Shinomura, T.; Xu, X.; Chua, N. The novel MYB protein EARLY-PHYTOCHROME-RESPONSIVE1 is a component of a slave circadian oscillator in Arabidopsis. Plant Cell 2003, 15, 2476–2488. [Google Scholar] [CrossRef] [PubMed]
- Harmer, S.L.; Hogenesch, J.B.; Straume, M.; Chang, H.S.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.A.; Kay, S.A. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Goodwin, S.M.; Boroff, V.L.; Liu, X.; Jenks, M.A. Cloning and characterization of the WAX2 gene of Arabidopsis involved in cuticle membrane and wax production. Plant Cell 2003, 15, 1170–1185. [Google Scholar] [CrossRef] [PubMed]
- Rowland, O.; Lee, R.; Franke, R.; Schreiber, L.; Kunst, L. The CER3 wax biosynthetic gene from Arabidopsis thaliana is allelic to WAX2/YRE/FLP1. FEBS Lett. 2007, 581, 3538–3544. [Google Scholar] [CrossRef] [PubMed]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; et al. Overexpression of Arabidopsis ECERIFERUM1 promotes wax very-long-chain alkane biosynthesis and influences plant response to biotic and abiotic stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Hofius, D.; Hajirezaei, M.; Geiger, M.; Tschiersch, H.; Melzer, M. RNAi-mediated tocopherol deficiency impairs photoassimilate export in transgenic potato plants. Plant Physiol. 2004, 135, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- Ament, K.; van Schie, C.C.; Bouwmeester, H.J.; Haring, M.A.; Schuurink, R.C. Induction of a leaf specific geranylgeranyl pyrophosphate synthase and emission of (E,E)-4,8,12-trimethyltrideca-1,3,7,11-tetraene in tomato are dependent on both jasmonic acid and salicylic acid signaling pathways. Planta 2006, 224, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Kraut, D.A.; Prakash, S.; Matouschek, A. To degrade or release: Ubiquitin-chain remodeling. Trends Cell Biol. 2007, 17, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Schulze-lefert, P. Plant immunity: The origami of receptor activation. Curr. Biol. 2004, 14, 22–24. [Google Scholar] [CrossRef]
- Khabudaev, K.V.; Petrova, D.P.; Grachev, M.A.; Likhoshway, Y.V. A new subfamily LIP of the major intrinsic proteins. BMC Genom. 2014, 15, 173. [Google Scholar] [CrossRef] [PubMed]
- Sabir, F.; Leandro, M.J.; Martins, A.P.; Loureiro-Dias, M.C.; Moura, T.F.; Soveral, G.; Prista, C. Exploring three PIPs and three TIPs of grapevine for transport of water and atypical substrates through heterologous expression in aqy-null Yeast. PLoS ONE 2014, 9, e102087. [Google Scholar] [CrossRef] [PubMed]
- Hocquellet, A.; Toorawa, P.; Bové, J.M.; Garnier, M. Detection and identification of the two Candidatus Liberobacter species associated with citrus Huanglongbing by PCR amplification of ribosomal protein genes of the β operon. Mol. Cell. Probes 1999, 13, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, C.; Li, Y.; Lam, T.; Yiu, S.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Krüger, P.; Selbig, J.; Müller, L.A.; Rhee, S.Y.; Stitt, M. MAPMAN: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. Cell Mol. Biol. 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Bioinform. Methods Protoc. 1999, 132, 365–386. [Google Scholar]
- Cheng, C.Z.; Yang, J.W.; Yan, H.B.; Zhang, Y.Y.; Lu, Z.M.; Zhong, G.Y. Expressing p20 hairpin RNA of Citrus tristeza virus confers Citrus aurantium with tolerance/resistance against stem pitting and seedling yellow CTV strains. J. Integr. Agric. 2015, 14, 1767–1777. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 408, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summary Detail | M1 | M2 | H1 | H2 |
|---|---|---|---|---|
| Number of total raw tags | 3,587,092 | 3,696,000 | 3,704,176 | 3,587,500 |
| Number of distinct tags | 200,301 | 207,328 | 191,280 | 230,525 |
| Number of total clean tags | 3,479,412 | 3,558,497 | 3,602,015 | 3,449,820 |
| Number of distinct tags | 92,796 | 89,297 | 89,324 | 110,577 |
| Total clean tags mapped to Gene | 2,715,908 | 2,778,658 | 2,506,722 | 2,668,438 |
| Percentage of clean tags mapped to Gene | 78.06% | 78.09% | 69.59% | 77.35% |
| Number of distinct tags mapped to Gene | 55,259 | 57,768 | 48,500 | 68,912 |
| Percentage of distinct tags mapped to Gene | 59.55% | 64.69% | 54.30% | 62.32% |
| Bin ID | Pathway Name | DEG Numbers | p-Value |
|---|---|---|---|
| 29.2.1 | protein. synthesis. ribosomal protein | 40 | 2.81 × 10−7 |
| 29.2 | protein. synthesis | 55 | 1.22 × 10−6 |
| 10 | cell wall | 71 | 6.35 × 10−5 |
| 29.2.1.2 | protein. synthesis. ribosomal protein. eukaryotic | 21 | 3.46 × 10−4 |
| 34 | transport | 126 | 5.49 × 10−3 |
| 28.1 | DNA. synthesis/chromatin structure | 26 | 7.96 × 10−3 |
| 29.2.1.2.1 | protein. synthesis. ribosomal protein. Eukaryotic. 40S subunit | 11 | 2.24 × 10−2 |
| 28.1.3 | DNA. synthesis/chromatin structure. histone | 7 | 2.65 × 10−2 |
| 10.2 | cell wall. cellulose synthesis | 9 | 3.18 × 10−2 |
| 16.1 | secondary metabolism. simple phenols | 5 | 4.46 × 10−2 |
| 29.2.1.1 | protein. synthesis. ribosomal protein. prokaryotic | 15 | 4.46 × 10−2 |
| 17.2 | hormone metabolism. auxin | 24 | 4.62 × 10−2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Cheng, C.; Jiang, B.; Jiang, N.; Zhang, Y.; Hu, M.; Zhong, G. Digital Gene Expression Analysis of Ponkan Mandarin (Citrus reticulata Blanco) in Response to Asia Citrus Psyllid-Vectored Huanglongbing Infection. Int. J. Mol. Sci. 2016, 17, 1063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071063
Zhong Y, Cheng C, Jiang B, Jiang N, Zhang Y, Hu M, Zhong G. Digital Gene Expression Analysis of Ponkan Mandarin (Citrus reticulata Blanco) in Response to Asia Citrus Psyllid-Vectored Huanglongbing Infection. International Journal of Molecular Sciences. 2016; 17(7):1063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071063
Chicago/Turabian StyleZhong, Yun, Chunzhen Cheng, Bo Jiang, Nonghui Jiang, Yongyan Zhang, Minlun Hu, and Guangyan Zhong. 2016. "Digital Gene Expression Analysis of Ponkan Mandarin (Citrus reticulata Blanco) in Response to Asia Citrus Psyllid-Vectored Huanglongbing Infection" International Journal of Molecular Sciences 17, no. 7: 1063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071063