
Isolation and Characterization of a Glycosyl Hydrolase Family 16 β-Agarase from a Mangrove Soil Metagenomic Library

Abstract
:1. Introduction
2. Results and Discussion
2.1. Construction of Metagenomic Library and Isolation of Agarase Gene
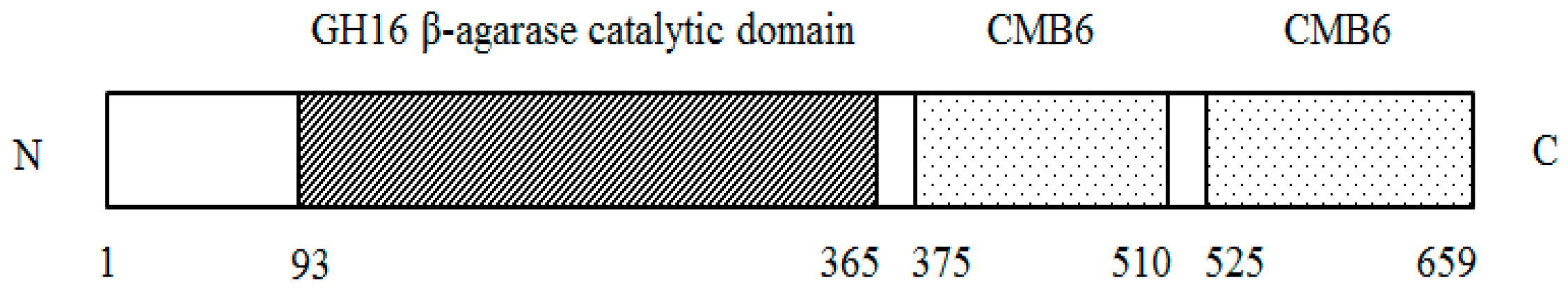
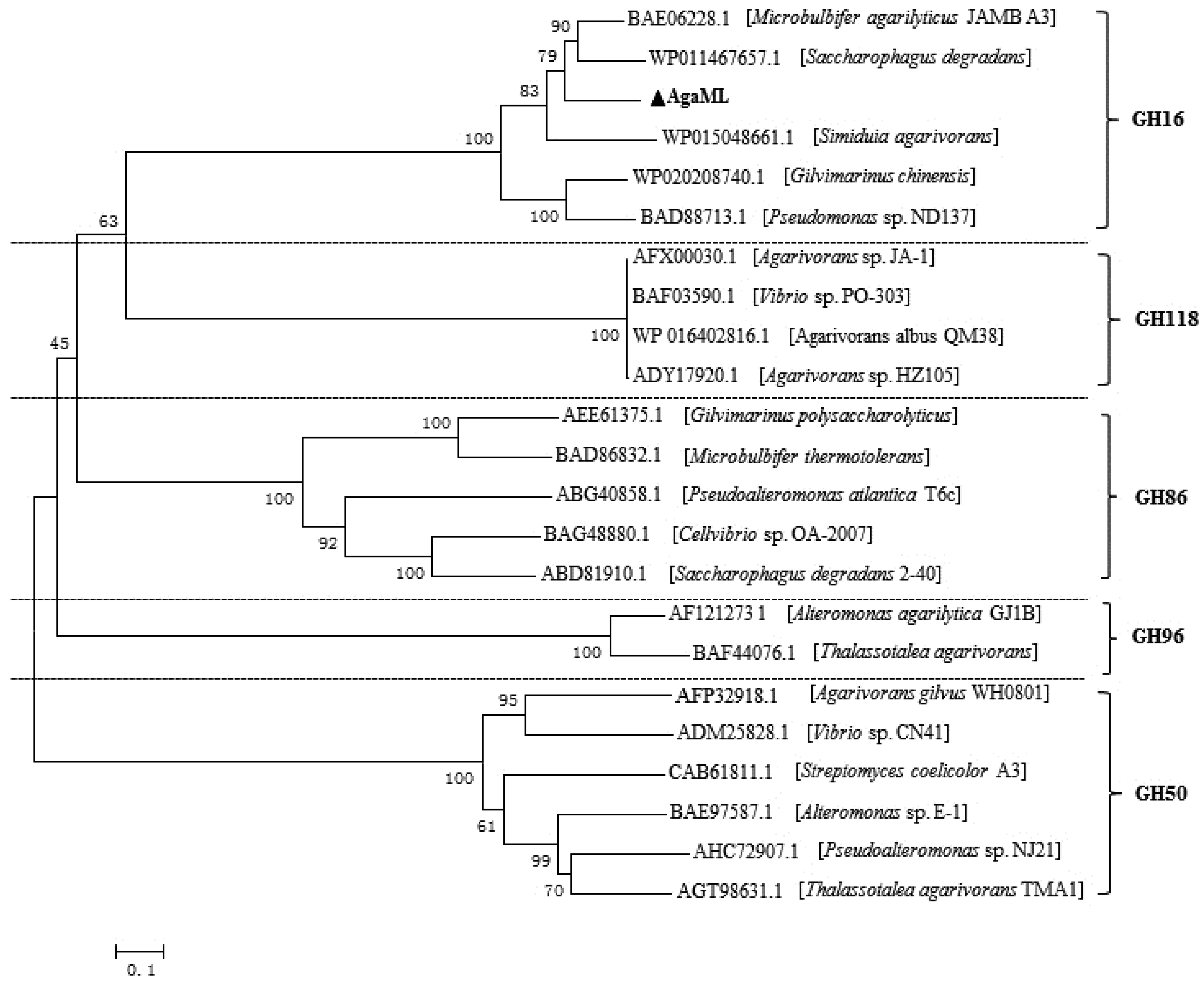
2.2. Sequence Analysis
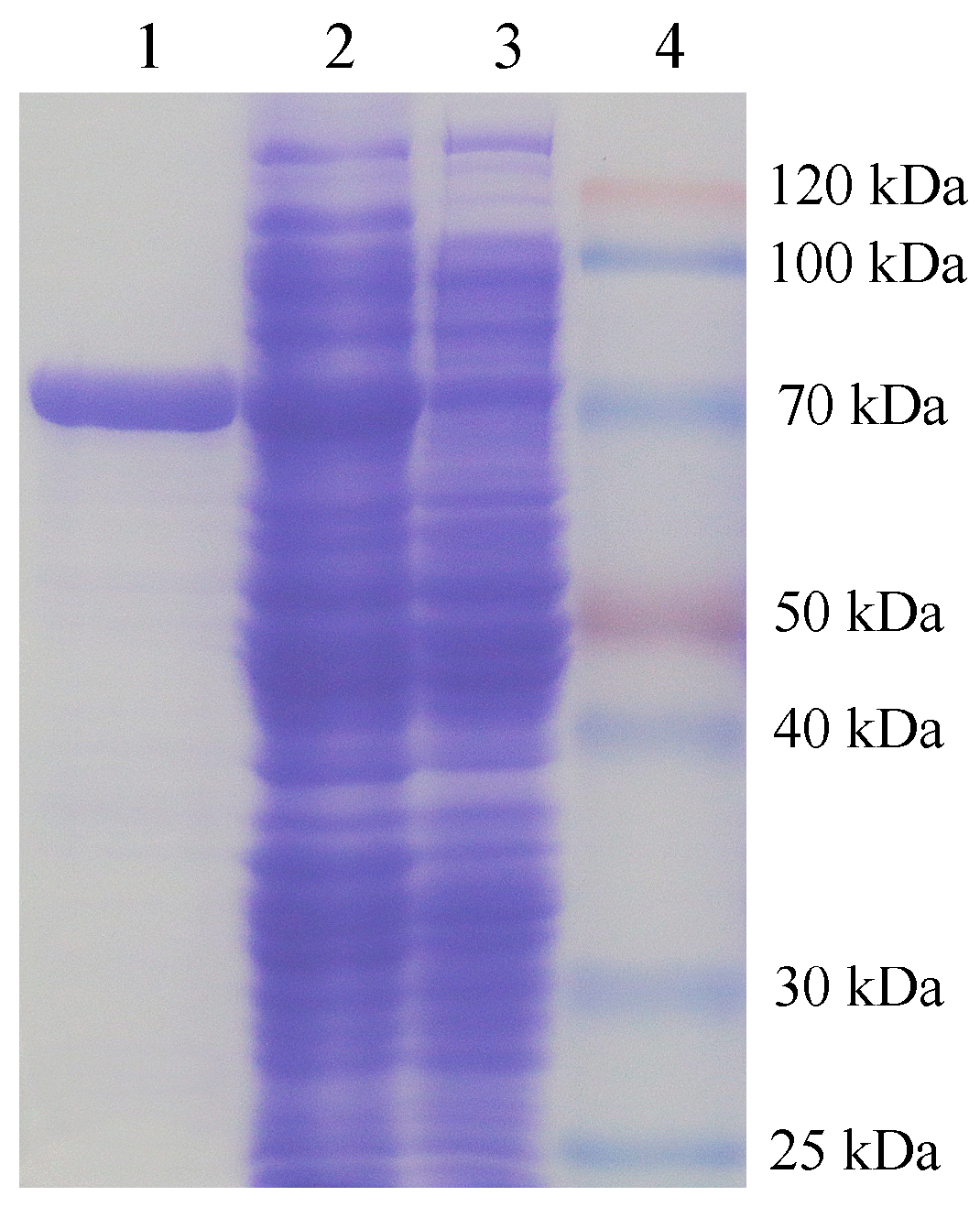
2.3. Expression and Purification of Recombination Agarase
2.4. Effects of pH and Temperature on the Activity of Recombination Agarase
2.5. Effects of Various Metal Ions and Reagents on the Activity of Recombination Agarase
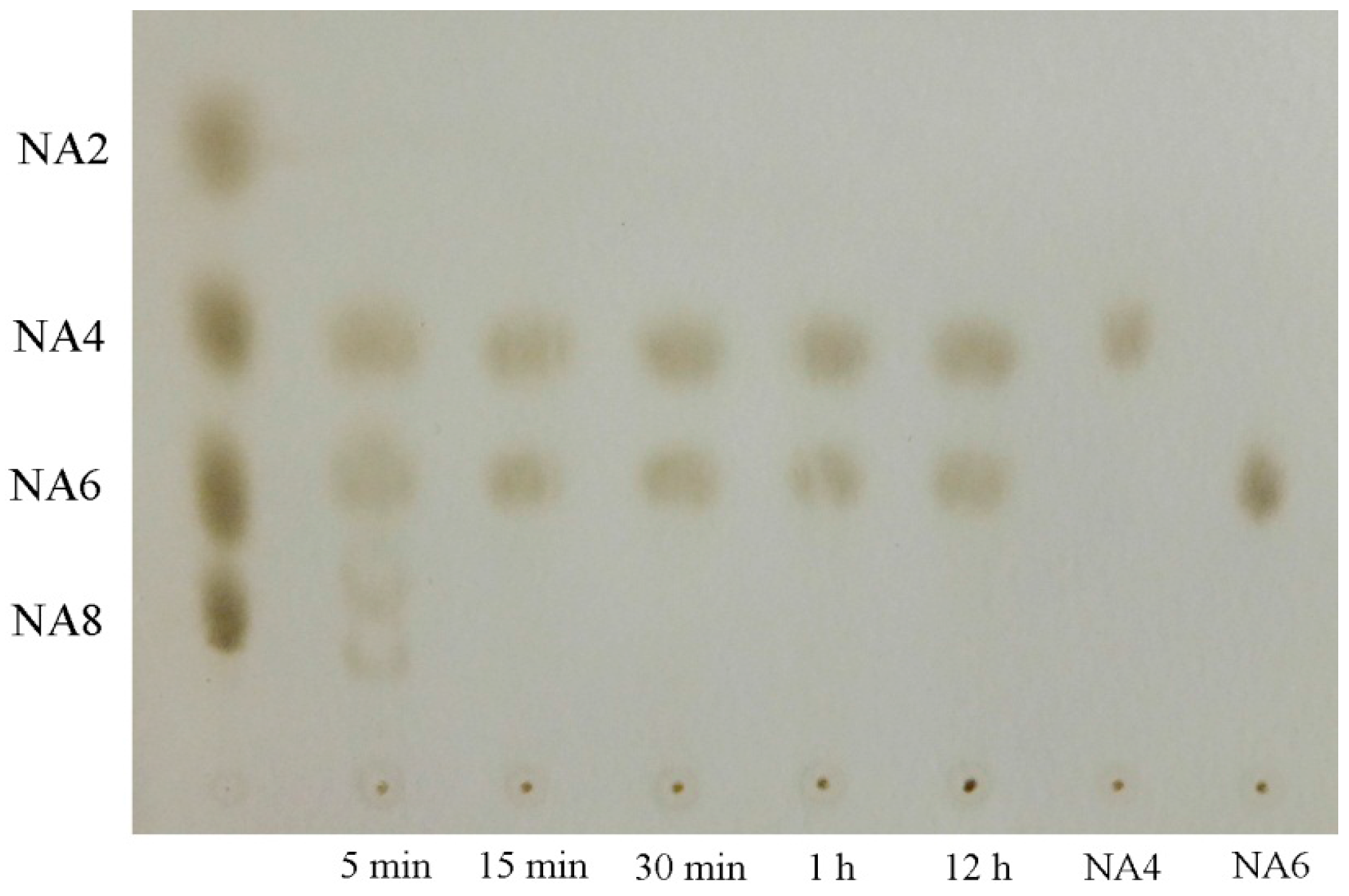
2.6. Analysis of the Hydrolysis Pattern and Products of Recombination Agarase
3. Materials and Methods
3.1. Strains, Plasmids, and Culture Conditions
3.2. Construction of Metagenomic Library
3.3. Library Screening and AgaML Gene Isolation
3.4. Sequence Analysis and Classification of AgaML
3.5. Cloning and Expression of AgaML
3.6. Purification of Recombinant Agarase
3.7. Agarase Activity Assay
3.8. Effects of pH and Temperature on Recombinant Agarase Activity
3.9. Effects of Various Metal Ions and Reagents on Recombinant Agarase Activity
3.10. Hydrolysis Products Analysis of Recombinant Agarase
3.11. Nucleotide Sequence Accession Number
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duckworth, M.; Yaphe, W. The structure of agar: Part I. Fractionation of a complex mixture of polysaccharides. Carbohydr. Res. 1971, 16, 189–197. [Google Scholar] [CrossRef]
- Araki, C. Seaweed polysaccharides. In Carbohydrate Chemistry of Substances of Biological Interests; Pergamon Press: London, UK, 1959; pp. 15–30. [Google Scholar]
- Fernández, L.E.; Valiente, O.G.; Mainardi, V.; Bello, J.L.; Vélez, H.; Rosado, A. Isolation and characterization of an antitumor active agar-type polysaccharide of Gracilaria dominguensis. Carbohydr. Res. 1989, 190, 77–83. [Google Scholar] [CrossRef]
- Takemoto, K. Plaque mutants of animal viruses. Progr. Med. Virol. 1965, 8, 314–348. [Google Scholar]
- Yoshizawa, Y.; Enomoto, A.; Todoh, H.; Ametani, A.; Kaminogawa, S. Activation of murine macrophages by polysaccharide fractions from marine algae (Porphyra yezoensis). Biosci. Biotechnol. Biochem. 1993, 57, 1862–1866. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, N.; Zhou, G.; Lu, X.; Xu, Z.; Li, Z. In vivo antioxidant activity of polysaccharide fraction from Porphyra haitanesis (Rhodephyta) in aging mice. Pharmacol. Res. 2003, 48, 151–155. [Google Scholar] [CrossRef]
- Ohta, Y.; Hatada, Y.; Miyazaki, M.; Nogi, Y.; Ito, S.; Horikoshi, K. Purification and characterization of a novel α-agarase from a Thalassomonas sp. Curr. Microbiol. 2005, 50, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Sawai, Y.; Suzuki, T.; Kawai, K. Purification and characterization of an extracellular β-agarase from Bacillus sp. MK03. J. Biosci. Bioeng. 2003, 95, 328–334. [Google Scholar] [CrossRef]
- Dong, J.; Hashikawa, S.; Konishi, T.; Tamaru, Y.; Araki, T. Cloning of the novel gene encoding β-agarase C from a marine bacterium, Vibrio sp. strain PO-303, and characterization of the gene product. Appl. Environ. Microbiol. 2006, 72, 6399–6401. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.Y.; Choi, Y.L.; Cho, Y.S.; Kim, B.K.; Jeon, B.S.; Cha, J.Y.; Kim, C.H.; Lee, Y.C. Cloning, expression and characterization of a β-agarase gene from a marine bacterium, Pseudomonas sp. SK38. Biotechnol. Lett. 2003, 25, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Park, G.T.; Kim, N.Y.; Lee, E.J.; Jang, M.K.; Shin, Y.G.; Park, G.S.; Kim, T.M.; Lee, J.H.; Lee, J.H.; et al. Cloning, expression, and characterization of a glycoside hydrolase family 50 β-agarase from a marine Agarivorans isolate. Biotechnol. Lett. 2006, 28, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Tamaru, Y.; Araki, T. A unique β-agarase, AgaA, from a marine bacterium, Vibrio sp. strain PO-303. Appl. Microbiol. Biotechnol. 2007, 74, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mou, H.; Jiang, X.; Guan, H. Characterization of a novel β-agarase from marine Alteromonas sp. SY37-12 and its degrading products. Appl. Microbiol. Biotechnol. 2006, 71, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Kim, H.J.; Ha, S.D.; Hwang, S.H.; Byun, D.S.; Lee, T.H.; Kong, J.Y. Purification and characterization of β-agarase from marine bacterium Bacillus cereus ASK202. Biotechnol. Lett. 1999, 21, 1011–1015. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [PubMed]
- Mai, Z.; Su, H.; Yang, J.; Huang, S.; Zhang, S. Cloning and characterization of a novel GH44 family endoglucanase from mangrove soil metagenomic library. Biotechnol. Lett. 2014, 36, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Lee, C.H.; Oh, T.K.; Song, J.K.; Yoon, J.H. Isolation and characterization of a novel lipase from a metagenomic library of tidal flat sediments: Evidence for a new family of bacterial lipases. Appl. Environ. Microbiol. 2006, 72, 7406–7409. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.J.; Hao, Z.Y.; Zeng, R.; Shen, P.H.; Li, J.F.; Wu, B. Characterization of a novel serine protease inhibitor gene from a marine metagenome. Mar. Drugs 2011, 9, 1487–1501. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Kang, S.; Park, S.; Yoon, H.; Kim, M.J.; Heu, S.; Ryu, S. Characterization of a novel amylolytic enzyme encoded by a gene from a soil-derived metagenomic library. Appl. Environ. Microbiol. 2004, 70, 7229–7235. [Google Scholar] [CrossRef] [PubMed]
- Voget, S.; Leggewie, C.; Uesbeck, A.; Raasch, C.; Jaeger, K.E.; Streit, W. Prospecting for novel biocatalysts in a soil metagenome. Appl. Environ. Microbiol. 2003, 69, 6235–6242. [Google Scholar] [CrossRef] [PubMed]
- Mai, Z.; Su, H.; Li, L.; Zhang, S. Construction of a mangrove soil metagenome library and identification of two novel β-glucosidase genes. Biotechnol. Bull. 2014, 6, 168–172. [Google Scholar]
- Daniel, R. The soil metagenome—A rich resource for the discovery of novel natural products. Curr. Opin. Biotechnol. 2004, 15, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Jaitinder, G.; Rixon, J.E.; Bolam, D.N.; Mcqueen-Mason, S.; Simpson, P.J.; Williamson, M.P.; Hazlewood, G.P.; Gilbert, H.J. The type II and X cellulose-binding domains of Pseudomonas xylanase: A potentiate catalytic activity against complex substrates by a common mechanism. Biochem. J. 1999, 342, 473–480. [Google Scholar]
- Strobel, K.L.; Pfeiffer, K.A.; Blanch, H.W.; Clark, D.S. Structural insights into the affinity of Cel7A carbohydrate-binding module for lignin. J. Biol. Chem. 2015, 290, 22818–22826. [Google Scholar] [CrossRef] [PubMed]
- Henshaw, J.L.; Bolam, D.N.; Pires, V.M.; Czjzek, M.; Henrissat, B.; Ferreira, L.M.; Fontes, C.M.; Gilbert, H.J. The family 6 carbohydrate binding module CmCBM6-2 contains two ligand-binding sites with distinct specificities. J. Biol. Chem. 2004, 279, 21552–21559. [Google Scholar] [CrossRef] [PubMed]
- McCartney, L.; Gilbert, H.J.; Bolam, D.N.; Boraston, A.B.; Knox, J.P. Glycoside hydrolase carbohydrate-binding modules as molecular probes for the analysis of plant cell wall polymers. Anal. Biochem. 2004, 326, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Yu, Z.; Xu, X. A novel β-agarase with high pH stability from marine Agarivorans sp. LQ48. Mar. Biotechnol. 2010, 12, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Henshaw, J.; Horne-Bitschy, A.; van Bueren, A.L.; Money, V.A.; Bolam, D.N.; Czjzek, M.; Ekborg, N.A.; Weiner, R.M.; Hutcheson, S.W.; Davies, G.J.; et al. Family 6 carbohydrate binding modules in β-agarases display exquisite selectivity for the non-reducing termini of agarose chains. J. Biol. Chem. 2006, 281, 17099–17107. [Google Scholar] [CrossRef] [PubMed]
- McCarter, J.D.; Withers, G.S. Mechanisms of enzymatic glycoside hydrolysis. Curr. Opin. Struct. Biol. 1994, 4, 885–892. [Google Scholar] [CrossRef]
- Kim, H.T.; Lee, S.; Lee, D.; Kim, H.S.; Bang, W.G.; Kim, K.H.; Choi, I.G. Overexpression and molecular characterization of Aga50D from Saccharophagus degradans 2-40: An exo-type β-agarase producing neoagarobiose. Appl. Microbiol. Biotechnol. 2010, 86, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Lin, H.; Kim, S.M. Purification and characterization of a novel β-agarase, AgaA34, from Agarivorans albus YKW-34. Appl. Microbiol. Biotechnol. 2008, 78, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Temuujin, U.; Chi, W.J.; Lee, S.Y.; Chang, Y.K.; Hong, S.K. Overexpression and biochemical characterization of DagA from Streptomyces coelicolor A3(2): An endo-type β-agarase producing neoagarotetraose and neoagarohexaose. Appl. Microbiol. Biotechnol. 2011, 92, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.H.; Wei, C.H.; Lu, W.J.; Shen, F.; Pan, C.L.; Lin, H.T.V. Extracellular production of a novel endo-β-agarase AgaA from Pseudomonas vesicularis MA103 that cleaves agarose into neoagarotetraose and neoagarohexaose. Int. J. Mol. Sci. 2015, 16, 5590–5603. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Mao, X.; Yang, M.; Mu, B.; Wei, D. Gene cloning, expression and characterisation of a new β-agarase, AgWH50C, producing neoagarobiose from Agarivorans gilvus WH0801. World J. Microbiol. Biotechnol. 2014, 30, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.J.; Seo, Y.B.; Chang, Y.K.; Lee, S.Y.; Hong, S.K. Cloning, expression, and biochemical characterization of a novel GH16 β-agarase AgaG1 from Alteromonas sp. GNUM-1. Appl. Microbiol. Biotechnol. 2014, 98, 4545–4555. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Kim, S.M. Agarase: Review of major sources, categories, purification method, enzyme characteristics and applications. Mar. Drugs 2010, 8, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Chen, X.; Chan, Z.; Zeng, R. Expression and characterization of a thermostable and pH-stable β-agarase encoded by a new gene from Flammeovirga pacifica WPAGA1. Process Biochem. 2015, 50, 1068–1075. [Google Scholar] [CrossRef]
- Lu, X.; Chu, Y.; Wu, Q.; Gu, Y.; Han, F.; Yu, W. Cloning, expression and characterization of a new agarase-encoding gene from marine Pseudoalteromonas sp. Biotechnol. Lett. 2009, 31, 1565–1570. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.B.; Lu, Y.; Chi, W.J.; Park, H.R.; Jeong, K.J.; Hong, S.K.; Chang, Y.K. Heterologous expression of a newly screened β-agarase from Alteromonas sp. GNUM1 in Escherichia coli and its application for agarose degradation. Process Biochem. 2014, 49, 430–436. [Google Scholar] [CrossRef]
- Yang, J.I.; Chen, L.C.; Shih, Y.Y.; Hsieh, C.; Chen, C.Y.; Chen, W.M.; Chen, C.C. Cloning and characterization of β-agarase AgaYT from Flammeovirga yaeyamensis strain YT. J. Biosci. Bioeng. 2011, 112, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Hatada, Y.; Nogi, Y.; Li, Z.; Ito, S.; Horikoshi, K. Cloning, expression, and characterization of a glycoside hydrolase family 86 β-agarase from a deep-sea Microbulbifer-like isolate. Appl. Microbiol. Biotechnol. 2004, 66, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Mao, X.; Du, Z.; Mu, B.; Wei, D. Cloning and characterisation of a novel neoagarotetraose-forming-β-agarase, AgWH50A from Agarivorans gilvus WH0801. Carbohyd. Res. 2014, 388, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Hatada, Y.; Ito, S.; Horikoshi, K. High-level expression of a neoagarobiose-producing β-agarase gene from Agarivorans sp. JAMB-A11 in Bacillus subtilis and enzymic properties of the recombinant enzyme. Biotechnol. Appl. Biochem. 2005, 41, 183–191. [Google Scholar] [PubMed]
- Osamu, A.; Kubo, H.; Nakamura, M.; Moriyama, H.; Nakasaki, K. Cloning of agarase gene from non-marine agarolytic bacterium Cellvibrio sp. J. Microbiol. Biotechnol. 2012, 22, 1237–1244. [Google Scholar]
- Xie, W.; Lin, B.; Zhou, Z.; Lu, G.; Lun, J.; Xia, C.; Li, S.; Hu, Z. Characterization of a novel β-agarase from an agar-degrading bacterium Catenovulum sp. X3. Appl. Microbiol. Biotechnol. 2013, 97, 4907–4915. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Mao, X.; Liu, N.; Qiu, Y.; Xue, C. Purification and characterization of two agarases from Agarivorans albus OAY02. Process Biochem. 2014, 49, 905–912. [Google Scholar] [CrossRef]
- Ma, C.; Lu, X.; Shi, C.; Li, J.; Gu, Y.; Ma, Y.; Chu, Y.; Han, F.; Gong, Q.; Yu, W. Molecular cloning and characterization of a novel β-agarase, AgaB, from marine Pseudoalteromonas sp. CY24. J. Biol. Chem. 2007, 282, 3747–3754. [Google Scholar] [CrossRef] [PubMed]
- Mai, Z.; Su, H.; Zhang, S. Characterization of a metagenome-derived β-glucosidase and its application in conversion of polydatin to resveratrol. Catalysts 2016, 6, 35. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents | Concentration (mM) | Relative Activity (%) a |
|---|---|---|
| None | - | 100.0 ± 4.2 b |
| Na+ | 100 | 103 ± 3.4 |
| K+ | 100 | 104 ± 3.5 |
| NH4+ | 100 | 69 ± 5.6 |
| Mn2+ | 1 | 115 ± 4.3 |
| Mg2+ | 1 | 87 ± 4.6 |
| Fe3+ | 1 | 98 ± 3.2 |
| Zn2+ | 1 | 74 ± 6.2 |
| Ca2+ | 1 | 108 ± 6.4 |
| Cu2+ | 1 | 75 ± 5.1 |
| Ba2+ | 1 | 113 ± 4.2 |
| Co2+ | 1 | 103 ± 3.6 |
| Cd2+ | 1 | 85 ± 4.3 |
| EDTA | 10 | 17 ± 5.7 |
| SDS | 10 | 73 ± 5.3 |
| Family | Protein | Strain | Products | References |
|---|---|---|---|---|
| GH16 | AgaML | Metagenomic library | NA4, NA6 | This study |
| AgaA | Pseudoalteromonas sp. CY24 | NA2, NA4, NA6 | [38] | |
| AgaG1 | Alteromonas sp. GNUM1 | NA2, NA4 | [39] | |
| AgaA | Agarivorans sp. LQ48 | NA4, NA6 | [27] | |
| AgaYT | Flammeovirga yaeyamensis | NA2, NA4 | [40] | |
| GH50 | RagaA11 | Agarivorans sp. JAMB-A11 | NA2 | [41] |
| Unnamed | Agarivorans sp. JA-1 | NA2, NA4 | [11] | |
| AgWH50A | Agarivorans gilus WH0801 | NA4 | [42] | |
| GH86 | AgaP4383 | Flammeovirga pacifica WPAGA1 | NA4, NA6 | [37] |
| AgaO | Microbulbifer sp. JAMB-A94 | NA6 | [43] | |
| AgaA | Cellvibrio sp. OA-2007 | NA2, NA4 | [44] | |
| GH118 | AgaXa | Catenovulum sp. X3 | NA6, NA8, NA10, NA12 | [45] |
| Agarase-a | Agarivorans albus OAY02 | NA2, NA4, NA6 | [46] | |
| AgaB | Pseudoalteromonas sp. CY24 | NA8, NA10 | [47] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, Z.; Su, H.; Zhang, S. Isolation and Characterization of a Glycosyl Hydrolase Family 16 β-Agarase from a Mangrove Soil Metagenomic Library. Int. J. Mol. Sci. 2016, 17, 1360. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081360
Mai Z, Su H, Zhang S. Isolation and Characterization of a Glycosyl Hydrolase Family 16 β-Agarase from a Mangrove Soil Metagenomic Library. International Journal of Molecular Sciences. 2016; 17(8):1360. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081360
Chicago/Turabian StyleMai, Zhimao, Hongfei Su, and Si Zhang. 2016. "Isolation and Characterization of a Glycosyl Hydrolase Family 16 β-Agarase from a Mangrove Soil Metagenomic Library" International Journal of Molecular Sciences 17, no. 8: 1360. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081360