
Bone Niches, Hematopoietic Stem Cells, and Vessel Formation


{kind=link}
{kind=link}
Abstract
:1. Introduction
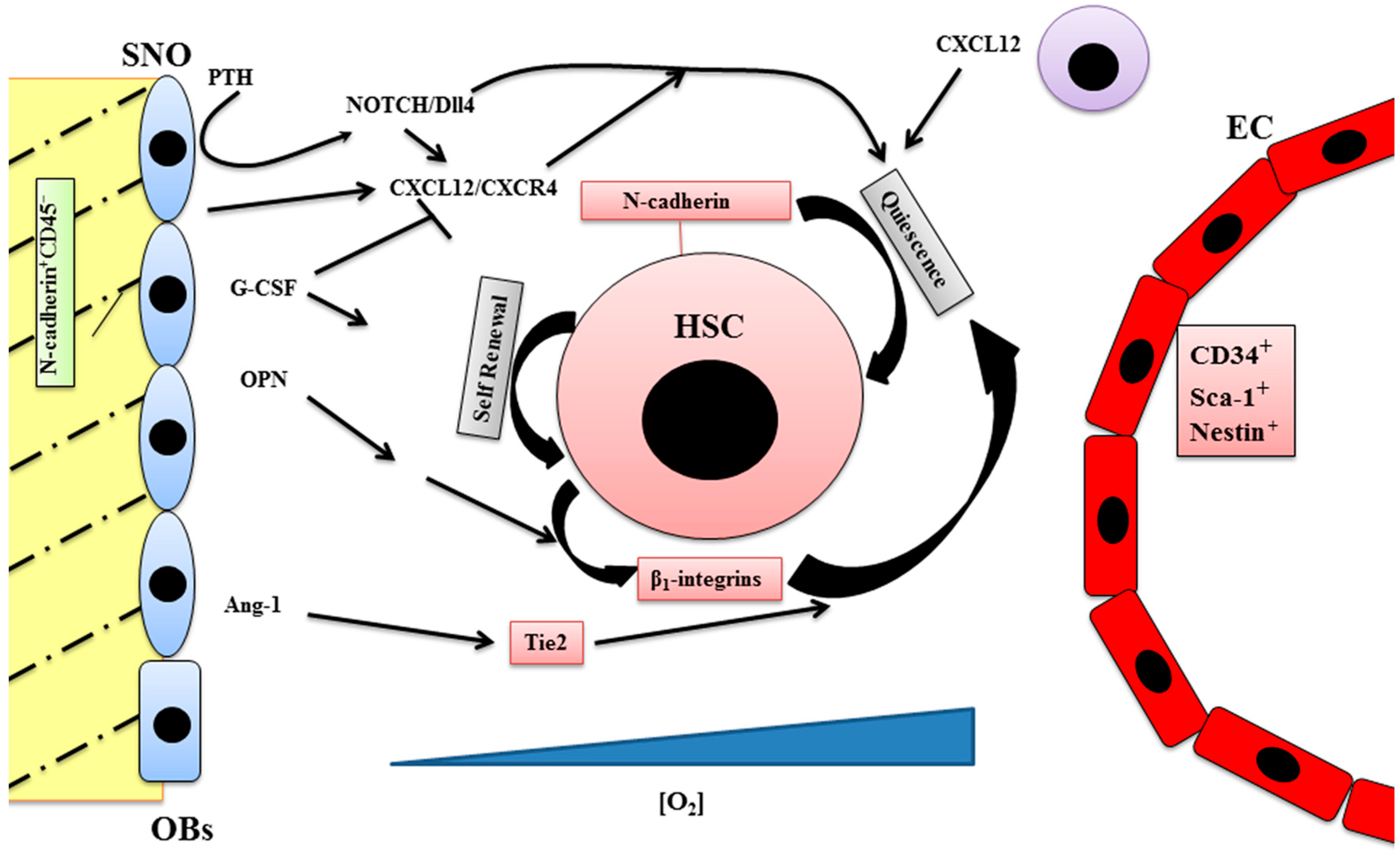
2. Endosteal Niche
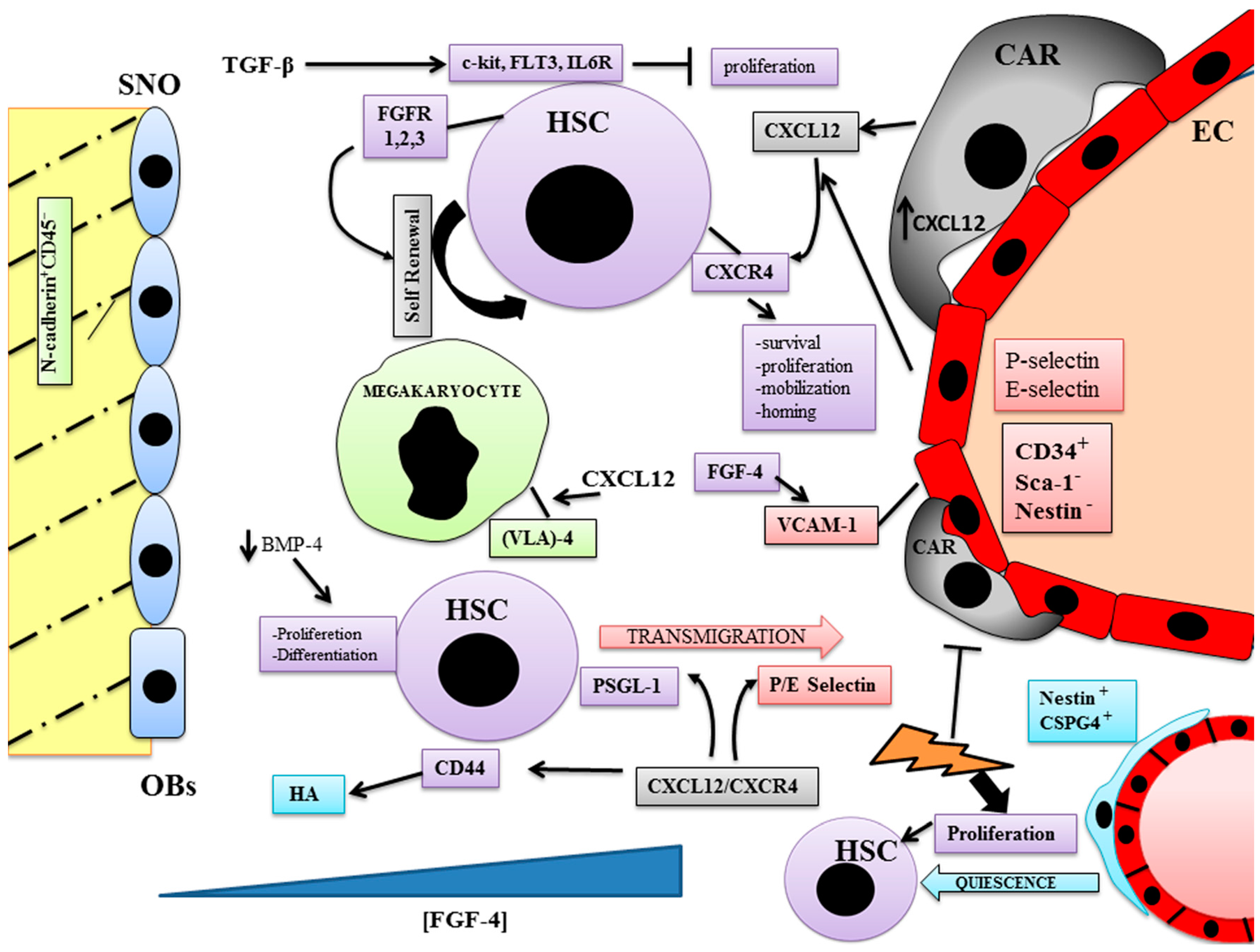
3. Vascular Niche
4. Bone Marrow Niches and Vessel Formation
5. Conclusions
Conflicts of Interest
References
- Birbrair, A.; Frenette, P.S. Niche heterogeneity in the bone marrow. Ann. N. Y. Acad. Sci. 2016, 1370, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.S. Regulation of hematopoietic stem cell fate. Oncogene 2002, 21, 3262–3269. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, L. Dissecting the bone marrow HSC niches. Cell Res. 2016, 26, 975–976. [Google Scholar] [CrossRef] [PubMed]
- Askmyr, M.; Sims, N.A.; Martin, T.J.; Purton, L.E. What is the true nature of the osteoblastic hematopoietic stem cell niche? Trends Endocrinol. Metab. 2009, 20, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Nico, B.; Vacca, A. Multiple myeloma as a model for the role of bone marrow niches in the control of angiogenesis. Int. Rev. Cell Mol. Biol. 2015, 314, 259–282. [Google Scholar] [PubMed]
- Mikkola, H.K.A. The journey of developing hematopoietic stem cells. Development 2006, 133, 3733–3744. [Google Scholar] [CrossRef] [PubMed]
- Lin, H. The stem-cell niche theory: Lessons from flies. Nat. Rev. Genet. 2002, 3, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Spradling, A.C. Stem cells and niches: Mechanisms that promote stem cell maintenance throughout life. Cell 2008, 132, 598–611. [Google Scholar] [CrossRef] [PubMed]
- Lord, B.I.; Testa, N.G.; Hendry, J.H. The relative spatial distributions of CFUs and CFUc in the normal mouse femur. Blood 1975, 46, 65–72. [Google Scholar] [PubMed]
- Calvi, L.M.; Adams, G.B.; Weibrecht, K.W.; Weber, J.M.; Olson, D.P.; Knight, M.C.; Martin, R.P.; Schipani, E.; Divieti, P.; Bringhurst, F.R.; et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 2003, 425, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Hauge, E.M.; Qvesel, D.; Eriksen, E.F.; Mosekilde, L.; Melsen, F. Cancellous bone remodeling occurs in specialized compartments lined by cells expressing osteoblastic markers. J. Bone Miner. Res. 2001, 16, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Arai, F.; Hosokawa, K.; Toyama, H.; Matsumoto, Y.; Suda, T. Role of n-cadherin in the regulation of hematopoietic stem cells in the bone marrow niche. Ann. N. Y. Acad. Sci. 2012, 1266, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, K.; Arai, F.; Yoshihara, H.; Iwasaki, H.; Nakamura, Y.; Gomei, Y.; Suda, T. Knockdown of N-cadherin suppresses the long-term engraftment of hematopoietic stem cells. Blood 2010, 116, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Niu, C.; Ye, L.; Huang, H.; He, X.; Tong, W.G.; Ross, J.; Haug, J.; Johnson, T.; Feng, J.Q.; et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature 2003, 425, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, A.M.; Revollo, L.D.; Woloszynek, J.R.; Civitelli, R.; Link, D.C. N-cadherin in osteolineage cells is not required for maintenance of hematopoietic stem cells. Blood 2012, 120, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Suda, T.; Arai, F.; Shimmura, S. Regulation of stem cells in the niche. Cornea 2005, 24, S12–S17. [Google Scholar] [CrossRef] [PubMed]
- Arai, F.; Hirao, A.; Suda, T. Regulation of hematopoietic stem cells by the niche. Trends Cardiovasc. Med. 2005, 15, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Taichman, R.S. Blood and bone: Two tissues whose fates are intertwined to create the hematopoietic stem-cell niche. Blood 2005, 105, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Askenasy, N.; Zorina, T.; Farkas, D.L.; Shalit, I. Transplanted hematopoietic cells seed in clusters in recipient bone marrow in vivo. Stem Cells 2002, 20, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K. Spatial localization of transplanted hemopoietic stem cells: Inferences for the localization of stem cell niches. Blood 2001, 97, 2293–2299. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.; Mauch, P.; Vergilio, J.A.; Sackstein, R.; Down, J.D. Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia. Proc. Natl. Acad. Sci. USA 2007, 104, 5431–5436. [Google Scholar] [CrossRef] [PubMed]
- Tothova, Z.; Kollipara, R.; Huntly, B.J.; Lee, B.H.; Castrillon, D.H.; Cullen, D.E.; McDowell, E.P.; Lazo-Kallanian, S.; Williams, I.R.; Sears, C.; et al. FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 2007, 128, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yu, S.; Zimmerman, G.; Wang, Y.; Myers, J.; Yu, V.W.C.; Huang, D.; Huang, X.; Shim, J.; Huang, Y.; et al. Notch receptor-ligand engagement maintains hematopoietic stem cell quiescence and niche retention. Stem Cells 2015, 33, 2280–2293. [Google Scholar] [CrossRef] [PubMed]
- Mancini, S.J.C. Jagged1-dependent notch signaling is dispensable for hematopoietic stem cell self-renewal and differentiation. Blood 2005, 105, 2340–2342. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, W.; Si, J.W.; Miao, X.Y.; Li, J.C.; Wang, Y.C.; Wang, Z.R.; Ma, J.; Zhao, X.C.; Li, Z.; et al. Notch signaling regulates CXCR4 expression and the migration of mesenchymal stem cells. Cell. Immunol. 2013, 281, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.K.; Segarra, M.; de la Luz Sierra, M.; Sainson, R.C.A.; Tosato, G.; Harris, A.L. Regulation of CXCR4 by the notch ligand δ-like 4 in endothelial cells. Cancer Res. 2008, 68, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Arai, F.; Iwasaki, H.; Hosokawa, K.; Kobayashi, I.; Gomei, Y.; Matsumoto, Y.; Yoshihara, H.; Suda, T. Isolation and characterization of endosteal niche cell populations that regulate hematopoietic stem cells. Blood 2010, 116, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Pelus, L.M.; Horowitz, D.; Cooper, S.C.; King, A.G. Peripheral blood stem cell mobilization. Crit. Rev. Oncol. Hematol. 2002, 43, 257–275. [Google Scholar] [CrossRef]
- Nervi, B.; Link, D.C.; di Persio, J.F. Cytokines and hematopoietic stem cell mobilization. J. Cell. Biochem. 2006, 99, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M. Differentiation and proliferation of hematopoietic stem cells. Blood 1993, 81, 2844–2853. [Google Scholar] [PubMed]
- Khodadi, E.; Shahrabi, S.; Shahjahani, M.; Azandeh, S.; Saki, N. Role of stem cell factor in the placental niche. Cell Tissue Res. 2016, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Ikuta, K.; Weissman, I.L. Evidence that hematopoietic stem cells express mouse c-kit but do not depend on steel factor for their generation. Proc. Natl. Acad. Sci. USA 1992, 89, 1502–1506. [Google Scholar] [CrossRef] [PubMed]
- Bradford, G.B.; Bertoncello, I. Noninvasive measurement of hematopoietic stem cell cycling and turnover by in vivo bromodeoxyuridine incorporation. In Hematopoietic Stem Cell Protocols; Humana Press: New York, NY, USA, 2002; pp. 79–92. [Google Scholar]
- Subramani, V.; Thiyagarajan, M.; Malathi, N.; Rajan, S.T. OPN—Revisited. J. Clin. Diagn. Res. 2015, 9, ZE10–ZE13. [Google Scholar] [CrossRef] [PubMed]
- Standal, T.; Borset, M.; Sundan, A. Role of osteopontin in adhesion, migration, cell survival and bone remodeling. Exp. Oncol. 2004, 26, 179–184. [Google Scholar] [PubMed]
- Stier, S.; Ko, Y.; Forkert, R.; Lutz, C.; Neuhaus, T.; Grünewald, E.; Cheng, T.; Dombkowski, D.; Calvi, L.M.; Rittling, S.R.; et al. Osteopontin is a hematopoietic stem cell niche component that negatively regulates stem cell pool size. J. Exp. Med. 2005, 201, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Lucas, D.; Frenette, P.S. Osteoblasts: Yes, they can. Blood 2008, 112, 455. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K. Osteopontin, a key component of the hematopoietic stem cell niche and regulator of primitive hematopoietic progenitor cells. Blood 2005, 106, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulou, T.; Craddock, C.; Nakamoto, B.; Priestley, G.V.; Wolf, N.S. The VLA4/VCAM-1 adhesion pathway defines contrasting mechanisms of lodgement of transplanted murine hemopoietic progenitors between bone marrow and spleen. Proc. Natl. Acad. Sci. USA 1995, 92, 9647–9651. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, M.; Le Pesteur, F.; Gagnerault, M.C.; Mary, J.Y.; Sainteny, F.; Lepault, F. Role of adhesion molecules in the homing and mobilization of murine hematopoietic stem and progenitor cells. Blood 1998, 92, 894–900. [Google Scholar] [PubMed]
- Cao, B.; Zhang, Z.; Grassinger, J.; Williams, B.; Heazlewood, C.K.; Churches, Q.I.; James, S.A.; Li, S.; Papayannopoulou, T.; Nilsson, S.K. Therapeutic targeting and rapid mobilization of endosteal HSC using a small molecule integrin antagonist. Nat. Commun. 2016, 7, 11007. [Google Scholar] [CrossRef] [PubMed]
- Alexander, W.S.; Roberts, A.W.; Nicola, N.A.; Li, R.; Metcalf, D. Deficiencies in progenitor cells of multiple hematopoietic lineages and defective megakaryocytopoiesis in mice lacking the thrombopoietic receptor c-Mpl. Blood 1996, 87, 2162–2170. [Google Scholar] [PubMed]
- Fliedner, T.M.; Graessle, D.; Paulsen, C.; Reimers, K. Structure and function of bone marrow hemopoiesis: Mechanisms of response to ionizing radiation exposure. Cancer Biother. Radiopharm. 2002, 17, 405–426. [Google Scholar] [CrossRef] [PubMed]
- Kusumbe, A.P.; Ramasamy, S.K.; Itkin, T.; Mäe, M.A.; Langen, U.H.; Betsholtz, C.; Lapidot, T.; Adams, R.H. Age-dependent modulation of vascular niches for haematopoietic stem cells. Nature 2016, 532, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Kopp, H.-G.; Hooper, A.T.; Avecilla, S.T.; Rafii, S. Functional heterogeneity of the bone marrow vascular niche. Ann. N. Y. Acad. Sci. 2009, 1176, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T. Microenvironmental niches in the bone marrow required for B-cell development. Nat. Rev. Immunol. 2006, 6, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Benayahu, D.; Akavia, U.D.; Shur, I. Differentiation of bone marrow stroma-derived mesenchymal cells. Curr. Med. Chem. 2007, 14, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Avecilla, S.T.; Hattori, K.; Heissig, B.; Tejada, R.; Liao, F.; Shido, K.; Jin, D.K.; Dias, S.; Zhang, F.; Hartman, T.E.; et al. Chemokine-mediated interaction of hematopoietic progenitors with the bone marrow vascular niche is required for thrombopoiesis. Nat. Med. 2003, 10, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.H.; Seo, J.B.; Hur, J.; Choi, J.H.; Kim, J.H.; Park, S.J.; Jo, S.H.; Lee, J.H.; Hwang, K.K.; Kim, S.H.; et al. Characterization of two types of endothelial progenitor cells (EPC). Korean Circ. J. 2004, 34, 304–313. [Google Scholar] [CrossRef]
- Maher, P.A. Modulation of the epidermal growth factor receptor by basic fibroblast growth factor. J. Cell. Physiol. 1993, 154, 350–358. [Google Scholar] [CrossRef] [PubMed]
- De Haan, G.; Weersing, E.; Dontje, B.; van Os, R.; Bystrykh, L.V.; Vellenga, E.; Miller, G. In vitro generation of long-term repopulating hematopoietic stem cells by fibroblast growth factor-1. Dev. Cell 2003, 4, 241–251. [Google Scholar] [CrossRef]
- Wright, D.E. Physiological migration of hematopoietic stem and progenitor cells. Science 2001, 294, 1933–1936. [Google Scholar] [CrossRef] [PubMed]
- Batard, P.; Monier, M.N.; Fortunel, N.; Ducos, K.; Sansilvestri-Morel, P.; Phan, T.; Hatzfeld, A.; Hatzfeld, J.A. TGF-(β)1 maintains hematopoietic immaturity by a reversible negative control of cell cycle and induces CD34 antigen up-modulation. J. Cell Sci. 2000, 113, 383–390. [Google Scholar] [PubMed]
- Scandura, J.M.; Boccuni, P.; Massague, J.; Nimer, S.D. Transforming growth factor-induced cell cycle arrest of human hematopoietic cells requires p57KIP2 up-regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 15231–15236. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.E.; Ruscetti, F.W.; Dubois, C.M.; Lee, J.; Boone, T.C.; Keller, J.R. Transforming growth factor-β trans-modulates the expression of colony stimulating factor receptors on murine hematopoietic progenitor cell lines. Blood 1991, 77, 1706–1716. [Google Scholar] [PubMed]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Bonnet, D.; Wu, D.; Murdoch, B.; Wrana, J.; Gallacher, L.; Dick, J.E. Bone morphogenetic proteins regulate the developmental program of human hematopoietic stem cells. J. Exp. Med. 1999, 189, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Willert, K.; Brown, J.D.; Danenberg, E.; Duncan, A.W.; Weissman, I.L.; Reya, T.; Yates, J.R.; Nusse, R. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature 2003, 423, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Blank, U.; Karlsson, G.; Karlsson, S. Signaling pathways governing stem-cell fate. Blood 2008, 111, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Duncan, A.W.; Rattis, F.M.; DiMascio, L.N.; Congdon, K.L.; Pazianos, G.; Zhao, C.; Yoon, K.; Cook, J.M.; Willert, K.; Gaiano, N.; et al. Integration of notch and Wnt signaling in hematopoietic stem cell maintenance. Nat. Immunol. 2005, 6, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Trowbridge, J.J.; Xenocostas, A.; Moon, R.T.; Bhatia, M. Glycogen synthase kinase-3 is an in vivo regulator of hematopoietic stem cell repopulation. Nat. Med. 2005, 12, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y. PSGL-1 participates in e-selectin-mediated progenitor homing to bone marrow: Evidence for cooperation between E-selectin ligands and 4 integrin. Blood 2003, 102, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Naor, D.; Nedvetzki, S.; Golan, I.; Melnik, L.; Faitelson, Y. CD44 in cancer. Crit. Rev. Clin. Lab. Sci. 2002, 39, 527–579. [Google Scholar] [CrossRef] [PubMed]
- Schmits, R.; Filmus, J.; Gerwin, N.; Senaldi, G.; Kiefer, F.; Kundig, T.; Wakeham, A.; Shahinian, A.; Catzavelos, C.; Rak, J.; et al. CD44 regulates hematopoietic progenitor distribution, granuloma formation, and tumorigenicity. Blood 1997, 90, 2217–2233. [Google Scholar] [PubMed]
- Oostendorp, R.A.J.; Ghaffari, S.; Eaves, C.J. Kinetics of in vivo homing and recruitment into cycle of hematopoietic cells are organ-specific but CD44-independent. Bone Marrow Transplant. 2000, 26, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Gadhoum, Z.; Leibovitch, M.P.; Qi, J.; Dumenil, D.; Durand, L.; Leibovitch, S.; Smadja-Joffe, F. CD44: A new means to inhibit acute myeloid leukemia cell proliferation via p27KIP1. Blood 2004, 103, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Charrad, R.S.; Li, Y.; Delpech, B.; Balitrand, N.; Clay, D.; Jasmin, C.; Chomienne, C.; Smadja-Joffe, F. Ligation of the CD44 adhesion molecule reverses blockage of differentiation in human acute myeloid leukemia. Nat. Med. 1999, 5, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K. Hyaluronan is synthesized by primitive hemopoietic cells, participates in their lodgment at the endosteum following transplantation, and is involved in the regulation of their proliferation and differentiation in vitro. Blood 2002, 101, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Avigdor, A.; Goichberg, P.; Shivtiel, S.; Dar, A.; Peled, A.; Samira, S.; Kollet, O.; Hershkoviz, R.; Alon, R.; Hardan, I.; et al. CD44 and hyaluronic acid cooperate with SDF-1 in the trafficking of human CD34+ stem/progenitor cells to bone marrow. Blood 2004, 103, 2981–2989. [Google Scholar] [CrossRef] [PubMed]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Mechanisms of angiogenesis and arteriogenesis. Nat. Med. 2000, 6, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; DeBusk, L.M.; Fukuda, K.; Fingleton, B.; Green-Jarvis, B.; Shyr, Y.; Matrisian, L.M.; Carbone, D.P.; Lin, P.C. Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell 2004, 6, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Ziegelhoeffer, T. Bone marrow-derived cells do not incorporate into the adult growing vasculature. Circ. Res. 2004, 94, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A. CXCR4: A key receptor in the crosstalk between tumor cells and their microenvironment. Blood 2006, 107, 1761–1767. [Google Scholar] [CrossRef] [PubMed]
- Ruiz de Almodovar, C.; Luttun, A.; Carmeliet, P. An SDF-1 trap for myeloid cells stimulates angiogenesis. Cell 2006, 124, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Hattori, K.; Dias, S.; Heissig, B.; Hackett, N.R.; Lyden, D.; Tateno, M.; Hicklin, D.J.; Zhu, Z.; Witte, L.; Crystal, R.G.; et al. Vascular endothelial growth factor and angiopoietin-1 stimulate postnatal hematopoiesis by recruitment of vasculogenic and hematopoietic stem cells. J. Exp. Med. 2001, 193, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Seandel, M.; Butler, J.; Lyden, D.; Rafii, S. A catalytic role for proangiogenic marrow-derived cells in tumor neovascularization. Cancer Cell 2008, 13, 181–183. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; McCabe, N.P.; Mahabeleshwar, G.H.; Somanath, P.R.; Phillips, D.R.; Byzova, T.V. The angiogenic response is dictated by β 3 integrin on bone marrow-derived cells. J. Cell Biol. 2008, 183, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, D.H.J. The role of endothelial progenitor and cardiac stem cells in the cardiovascular adaptations to age and exercise. Front. Biosci. 2009, 14, 4685–4702. [Google Scholar] [CrossRef]
- Khakoo, A.Y.; Finkel, T. Endothelial progenitor cells. Ann. Rev. Med. 2005, 56, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Rehman, J. Peripheral blood “endothelial progenitor cells” are derived from monocyte/macrophages and secrete angiogenic growth factors. Circulation 2003, 107, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Takahashi, T.; Masuda, H.; Kalka, C.; Chen, D.; Iwaguro, H.; Inai, Y.; Silver, M.; Isner, J.M. VEGF contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells. EMBO J. 1999, 18, 3964–3972. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Kalka, C.; Masuda, H.; Chen, D.; Silver, M.; Kearney, M.; Magner, M.; Isner, J.M.; Asahara, T. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat. Med. 1999, 5, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Akeno, N.; Robins, J.; Zhang, M.; Czyzyk-Krzeska, M.F.; Clemens, T.L. Induction of vascular endothelial growth factor by IGF-I in osteoblast-like cells is mediated by the PI3K signaling pathway through the hypoxia-inducible factor-2α. Endocrinology 2002, 143, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Rankin, E.B.; Wu, C.; Khatri, R.; Wilson, T.L.S.; Andersen, R.; Araldi, E.; Rankin, A.L.; Yuan, J.; et al. The HIF signaling pathway in osteoblasts directly modulates erythropoiesis through the production of EPO. Cell 2012, 149, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Sautina, L.; Sautin, Y.; Beem, E.; Zhou, Z.; Schuler, A.; Brennan, J.; Zharikov, S.I.; Diao, Y.; Bungert, J.; Segal, M.S. Induction of nitric oxide by erythropoietin is mediated by the common receptor and requires interaction with VEGF receptor 2. Blood 2009, 115, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Bahlmann, F.H. Erythropoietin regulates endothelial progenitor cells. Blood 2003, 103, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Aicher, A.; Heeschen, C.; Mildner-Rihm, C.; Urbich, C.; Ihling, C.; Technau-Ihling, K.; Zeiher, A.M.; Dimmeler, S. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat. Med. 2003, 9, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Brandi, M.L.; Collin-Osdoby, P. Vascular biology and the skeleton. J. Bone Miner. Res. 2006, 21, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Manalo, D.J. Transcriptional regulation of vascular endothelial cell responses to hypoxia by HIF-1. Blood 2005, 105, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Sorescu, G.P.; Sykes, M.; Weiss, D.; Platt, M.O.; Saha, A.; Hwang, J.; Boyd, N.; Boo, Y.C.; Vega, J.D.; Taylor, W.R.; et al. Bone morphogenic protein 4 produced in endothelial cells by oscillatory shear stress stimulates an inflammatory response. J. Biol. Chem. 2003, 278, 31128–31135. [Google Scholar] [CrossRef] [PubMed]
- Epstein, F.H.; Levin, E.R. Endothelins. N. Engl. J. Med. 1995, 333, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, K.; Karsdal, M.; Delaisse, J.M.; Engsig, M.T. RANKL and vascular endothelial growth factor (VEGF) induce osteoclast chemotaxis through an ERK1/2-dependent mechanism. J. Biol. Chem. 2003, 278, 48745–48753. [Google Scholar] [CrossRef] [PubMed]
- Trebec-Reynolds, D.P.; Voronov, I.; Heersche, J.N.M.; Manolson, M.F. VEGF—A expression in osteoclasts is regulated by NF-κB induction of HIF-1α. J. Cell. Biochem. 2010, 110, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.-Y.; Liu, C.; Tonelli-Zasarsky, R.M.L.; Simmons, C.A.; You, L. Osteocyte apoptosis is mechanically regulated and induces angiogenesis in vitro. J. Orthop. Res. 2010, 29, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Juffer, P.; Jaspers, R.T.; Lips, P.; Bakker, A.D.; Klein-Nulend, J. Expression of muscle anabolic and metabolic factors in mechanically loaded MLO-Y4 osteocytes. AJP 2011, 302, E389–E395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, S.K.; Kusumbe, A.P.; Wang, L.; Adams, R.H. Endothelial notch activity promotes angiogenesis and osteogenesis in bone. Nature 2014, 507, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Himburg, H.A.; Harris, J.R.; Ito, T.; Daher, P.; Russell, J.L.; Quarmyne, M.; Doan, P.L.; Helms, K.; Nakamura, M.; Fixsen, E.; et al. Pleiotrophin regulates the retention and self-renewal of hematopoietic stem cells in the bone marrow vascular niche. Cell Rep. 2012, 2, 1774. [Google Scholar] [CrossRef]
- Papadimitriou, E.; Mikelis, C.; Lampropoulou, E.; Koutsioumpa, M.; Theochari, K.; Tsirmoula, S.; Theodoropoulou, C.; Lamprou, M.; Sfaelou, E.; Vourtsis, D.; et al. Roles of pleiotrophin in tumor growth and angiogenesis. Eur. Cytokine Netw. 2009, 20, 180–190. [Google Scholar] [PubMed]
- Heiss, C.; Wong, M.L.; Block, V.I.; Lao, D.; Real, W.M.; Yeghiazarians, Y.; Lee, R.J.; Springer, M.L. Pleiotrophin induces nitric oxide dependent migration of endothelial progenitor cells. J. Cell. Physiol. 2008, 215, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Tare, R.S.; Oreffo, R.O.; Clarke, N.M.; Roach, H.I. Pleiotrophin/osteoblast-stimulating factor 1: Dissecting its diverse functions in bone formation. J. Bone Miner. Res. 2002, 17, 2009–2020. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamma, R.; Ribatti, D. Bone Niches, Hematopoietic Stem Cells, and Vessel Formation. Int. J. Mol. Sci. 2017, 18, 151. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010151
Tamma R, Ribatti D. Bone Niches, Hematopoietic Stem Cells, and Vessel Formation. International Journal of Molecular Sciences. 2017; 18(1):151. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010151
Chicago/Turabian StyleTamma, Roberto, and Domenico Ribatti. 2017. "Bone Niches, Hematopoietic Stem Cells, and Vessel Formation" International Journal of Molecular Sciences 18, no. 1: 151. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010151