
Mesenchymal Stem Cells from Adipose Tissue in Clinical Applications for Dermatological Indications and Skin Aging

Abstract
:
1. Introduction
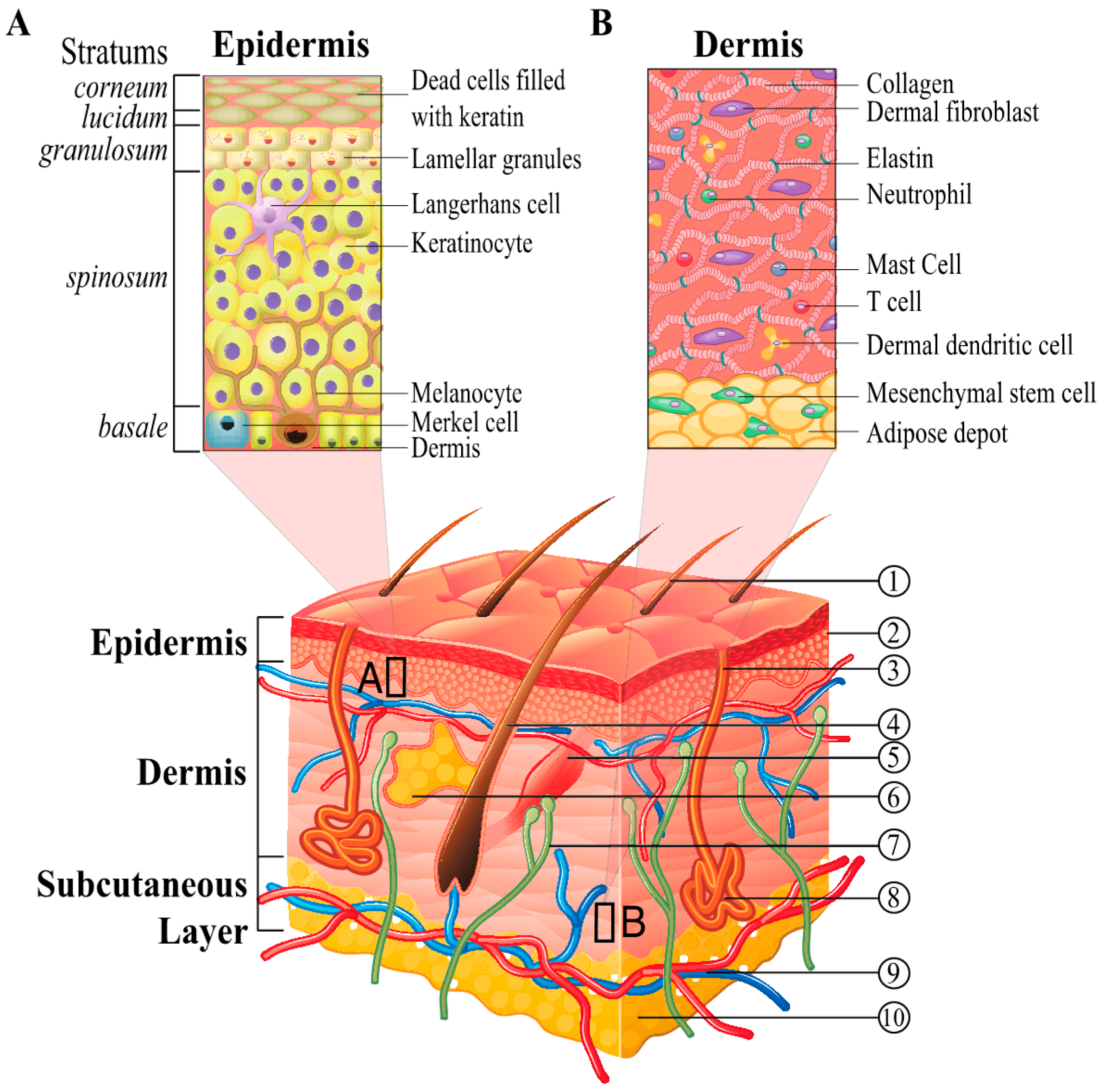
2. Skin Structure and Cellular Composition as It Relates to Regenerative Medicine
3. Dermatological Aging and Practical Use and Significance of Non-Invasive Methods for Its Assessment
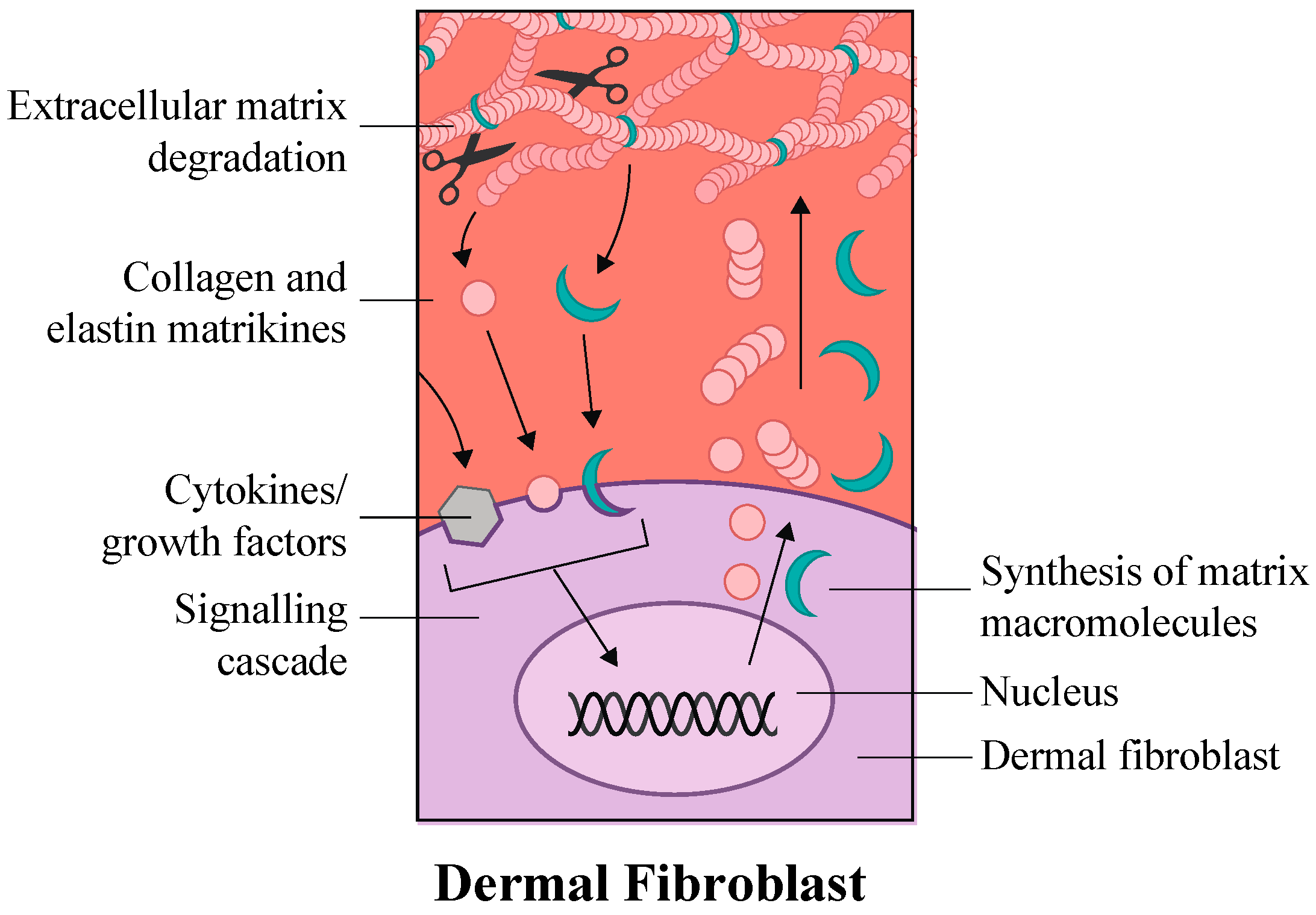
4. Signaling Pathways of Cellular Interactions within Skin
5. Wound Healing Paradigms
6. From Lipoaspirate to Adipose-Derived Stem Cells (ADSCs)
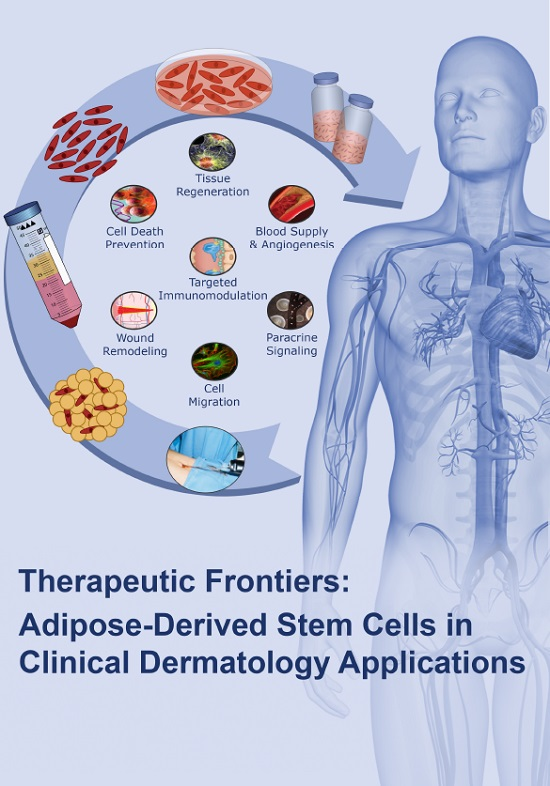
7. Adipose-Derived Stem Cells and Their Applications in Clinical Dermatology
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Makrantonaki, E.; Zouboulis, C.C. Molecular mechanisms of skin aging: State of the art. Ann. N. Y. Acad. Sci. 2007, 1119, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Makrantonaki, E. Clinical aspects and molecular diagnostics of skin aging. Clin. Dermatol. 2011, 29, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Boschnakow, A. Chronological ageing and photoageing of the human sebaceous gland. Clin. Exp. Dermatol. 2001, 26, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Fuchs, E. Epidermal homeostasis: A balancing act of stem cells in the skin. Nat. Rev. Mol. Cell Biol. 2009, 10, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Horsley, V.; Fuchs, E. Epithelial stem cells: Turning over new leaves. Cell 2007, 128, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Fuchs, E. Epidermal stem cells of the skin. Annu. Rev. Cell Dev. Biol. 2006, 22, 339–373. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; O’Carroll, D.; Pasolli, H.A.; Zhang, Z.; Dietrich, F.S.; Tarakhovsky, A.; Fuchs, E. Morphogenesis in skin is governed by discrete sets of differentially expressed microRNAs. Nat. Genet. 2006, 38, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Tollervey, J.R.; Lunyak, V.V. Epigenetics: Judge, jury and executioner of stem cell fate. Epigenetics 2012, 7, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Morgner, J.; Ghatak, S.; Jakobi, T.; Dieterich, C.; Aumailley, M.; Wickstrom, S.A. Integrin-linked kinase regulates the niche of quiescent epidermal stem cells. Nat. Commun. 2015, 6, 8198. [Google Scholar] [CrossRef] [PubMed]
- Odorisio, T. Epigenetic Control of Skin Re-Epithelialization: The NF-κB/JMJD3 Connection. J. Investig. Dermatol. 2016, 136, 738–740. [Google Scholar] [CrossRef] [PubMed]
- Kilroy, G.E.; Foster, S.J.; Wu, X.; Ruiz, J.; Sherwood, S.; Heifetz, A.; Ludlow, J.W.; Stricker, D.M.; Potiny, S.; Green, P.; et al. Cytokine profile of human adipose-derived stem cells: Expression of angiogenic, hematopoietic, and pro-inflammatory factors. J. Cell. Physiol. 2007, 212, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Cappuzzello, C.; Doni, A.; Dander, E.; Pasqualini, F.; Nebuloni, M.; Bottazzi, B.; Mantovani, A.; Biondi, A.; Garlanda, C.; D’Amico, G. Mesenchymal Stromal Cell-Derived PTX3 Promotes Wound Healing via Fibrin Remodeling. J. Investig. Dermatol. 2016, 136, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Ozpur, M.A.; Guneren, E.; Canter, H.I.; Karaaltin, M.V.; Ovali, E.; Yogun, F.N.; Baygol, E.G.; Kaplan, S. Generation of Skin Tissue Using Adipose Tissue-Derived Stem Cells. Plast. Reconstr. Surg. 2016, 137, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, E.T.; Gustafson, M.P.; Dudakovic, A.; Riester, S.M.; Garces, C.G.; Paradise, C.R.; Takai, H.; Karperien, M.; Cool, S.; Sampen, H.J.; et al. Identification and validation of multiple cell surface markers of clinical-grade adipose-derived mesenchymal stromal cells as novel release criteria for good manufacturing practice-compliant production. Stem Cell Res. Ther. 2016, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Menon, G.K. New insights into skin structure: Scratching the surface. Adv. Drug Deliv. Rev. 2002, 54 (Suppl. S1), S3–S17. [Google Scholar] [CrossRef]
- Elias, P.M.; Friend, D.S. The permeability barrier in mammalian epidermis. J. Cell Biol. 1975, 65, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Nemes, Z.; Steinert, P.M. Bricks and mortar of the epidermal barrier. Exp. Mol. Med. 1999, 31, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Barry, B.W. Novel mechanisms and devices to enable successful transdermal drug delivery. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2001, 14, 101–114. [Google Scholar] [CrossRef]
- Foldvari, M. Non-invasive administration of drugs through the skin: Challenges in delivery system design. Pharm. Sci. Technol. Today 2000, 3, 417–425. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Ponec, M. The skin barrier in healthy and diseased state. Biochim. Biophys. Acta 2006, 1758, 2080–2095. [Google Scholar] [CrossRef] [PubMed]
- Mesa, K.R.; Rompolas, P.; Zito, G.; Myung, P.; Sun, T.Y.; Brown, S.; Gonzalez, D.G.; Blagoev, K.B.; Haberman, A.M.; Greco, V. Niche-induced cell death and epithelial phagocytosis regulate hair follicle stem cell pool. Nature 2015, 522, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Green, H. Changes in keratin gene expression during terminal differentiation of the keratinocyte. Cell 1980, 19, 1033–1042. [Google Scholar] [CrossRef]
- Niu, P.; Smagul, A.; Wang, L.; Sadvakas, A.; Sha, Y.; Perez, L.M.; Nussupbekova, A.; Amirbekov, A.; Akanov, A.A.; Galvez, B.G.; et al. Transcriptional profiling of interleukin-2-primed human adipose derived mesenchymal stem cells revealed dramatic changes in stem cells response imposed by replicative senescence. Oncotarget 2015, 6, 17938–17957. [Google Scholar] [CrossRef] [PubMed]
- Astori, G.; Vignati, F.; Bardelli, S.; Tubio, M.; Gola, M.; Albertini, V.; Bambi, F.; Scali, G.; Castelli, D.; Rasini, V.; et al. “In vitro” and multicolor phenotypic characterization of cell subpopulations identified in fresh human adipose tissue stromal vascular fraction and in the derived mesenchymal stem cells. J. Transl. Med. 2007, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Varma, M.J.; Breuls, R.G.; Schouten, T.E.; Jurgens, W.J.; Bontkes, H.J.; Schuurhuis, G.J.; van Ham, S.M.; van Milligen, F.J. Phenotypical and functional characterization of freshly isolated adipose tissue-derived stem cells. Stem Cells Dev. 2007, 16, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Roh, N.K.; Kim, M.J.; Lee, Y.W.; Choe, Y.B.; Ahn, K.J. A Split-Face Study of the Effects of a Stabilized Hyaluronic Acid-Based Gel of Nonanimal Origin for Facial Skin Rejuvenation Using a Stamp-Type Multineedle Injector: A Randomized Clinical Trial. Plast. Reconstr. Surg. 2016, 137, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.M.; Gibran, N.S. Mesenchymal stem cells: Paracrine signaling and differentiation during cutaneous wound repair. Exp. Cell Res. 2010, 316, 2213–2219. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Permana, P.A.; Owecki, M.; Chaldakov, G.N.; Bohm, M.; Hausman, G.; Lapiere, C.M.; Atanassova, P.; Sowinski, J.; Fasshauer, M.; et al. What are subcutaneous adipocytes really good for? Exp. Dermatol. 2007, 16, 45–70. [Google Scholar] [PubMed]
- Jackson, W.M.; Nesti, L.J.; Tuan, R.S. Concise review: Clinical translation of wound healing therapies based on mesenchymal stem cells. Stem Cells Transl. Med. 2012, 1, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Jackson, W.M.; Nesti, L.J.; Tuan, R.S. Mesenchymal stem cell therapy for attenuation of scar formation during wound healing. Stem Cell Res. Ther. 2012, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Naderi, N.; Combellack, E.J.; Griffin, M.; Sedaghati, T.; Javed, M.; Findlay, M.W.; Wallace, C.G.; Mosahebi, A.; Butler, P.E.; Seifalian, A.M.; et al. The regenerative role of adipose-derived stem cells (adsc) in plastic and reconstructive surgery. Int. Wound J. 2017, 14, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.A.; Horsley, V. Intradermal adipocytes mediate fibroblast recruitment during skin wound healing. Development 2013, 140, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Kato, J.; Kamiya, H.; Himeno, T.; Shibata, T.; Kondo, M.; Okawa, T.; Fujiya, A.; Fukami, A.; Uenishi, E.; Seino, Y.; et al. Mesenchymal stem cells ameliorate impaired wound healing through enhancing keratinocyte functions in diabetic foot ulcerations on the plantar skin of rats. J. Diabetes Complicat. 2014, 28, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Correa, D. PDGF in bone formation and regeneration: New insights into a novel mechanism involving MSCs. J. Orthop. Res. 2011, 29, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Park, B.S.; Sung, J.H.; Yang, J.M.; Park, S.B.; Kwak, S.J.; Park, J.S. Wound healing effect of adipose-derived stem cells: A critical role of secretory factors on human dermal fibroblasts. J. Dermatol. Sci. 2007, 48, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.F.; Underhill, C.B.; Hahn, P.J.; Brown, D.B.; Uitto, J. Chronic sun exposure alters both the content and distribution of dermal glycosaminoglycans. Br. J. Dermatol. 1996, 135, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Waller, J.M.; Maibach, H.I. Age and skin structure and function, a quantitative approach (II): Protein, glycosaminoglycan, water, and lipid content and structure. Skin Res. Technol. 2006, 12, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Functional and physiological characteristics of the aging skin. Aging Clin. Exp. Res. 2008, 20, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Suter-Widmer, J.; Elsner, P. The Irritant Contact Dermatitis Syndrome; CRC Press: Boca Raton, FL, USA, 1996; pp. 257–265. [Google Scholar]
- Waller, J.M.; Maibach, H.I. Age and skin structure and function, a quantitative approach (I): Blood flow, pH, thickness, and ultrasound echogenicity. Skin Res. Technol. 2005, 11, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Oriba, H.A.; Bucks, D.A.; Maibach, H.I. Percutaneous absorption of hydrocortisone and testosterone on the vulva and forearm: Effect of the menopause and site. Br. J. Dermatol. 1996, 134, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.O.; Leffell, D.J. Preoperative assessment of the elderly patient. Dermatol. Clin. 1997, 15, 583–593. [Google Scholar] [CrossRef]
- Kaya, G.; Saurat, J.H. Dermatoporosis: A chronic cutaneous insufficiency/fragility syndrome. Clinicopathological features, mechanisms, prevention and potential treatments. Dermatology 2007, 215, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Kaya, G.; Tran, C.; Sorg, O.; Hotz, R.; Grand, D.; Carraux, P.; Didierjean, L.; Stamenkovic, I.; Saurat, J.H. Hyaluronate fragments reverse skin atrophy by a CD44-dependent mechanism. PLoS Med. 2006, 3, e493. [Google Scholar] [CrossRef] [PubMed]
- Nagai, R.; Mori, T.; Yamamoto, Y.; Kaji, Y.; Yonei, Y. Significance of advanced glycation end products in ageing-related disease. Anti Ageing Med. 2010, 7, 112–119. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Winsz-Szczotka, K.; Klimek, K.; Olczyk, K. Plasma biomarkers of oxidative and AGE-mediated damage of proteins and glycosaminoglycans during healthy ageing: A possible association with ECM metabolism. Mech. Ageing Dev. 2012, 133, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Nedic, O.; Rattan, S.I.; Grune, T.; Trougakos, I.P. Molecular effects of advanced glycation end products on cell signalling pathways, ageing and pathophysiology. Free Radic. Res. 2013, 47 (Suppl. S1), 28–38. [Google Scholar] [CrossRef] [PubMed]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Characteristics of the Aging Skin. Adv. Wound Care 2013, 2, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Alkilani, A.Z.; McCrudden, M.T.; Donnelly, R.F. Transdermal Drug Delivery: Innovative Pharmaceutical Developments Based on Disruption of the Barrier Properties of the stratum corneum. Pharmaceutics 2015, 7, 438–470. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.J.; Jung, H.J.; Schrammek-Drusios, M.C.; Lee, S.N.; Kim, J.H.; Kwon, S.B.; An, I.S.; An, S.; Ahn, K.J. Instrumental evaluation of anti-aging effects of cosmetic formulations containing palmitoyl peptides, Silybum marianum seed oil, vitamin E and other functional ingredients on aged human skin. Exp. Ther. Med. 2016, 12, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Sotoodian, B.; Maibach, H.I. Noninvasive test methods for epidermal barrier function. Clin. Dermatol. 2012, 30, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.; Hong, J.Y.; Choi, S.Y.; Park, K.Y.; Kim, B.J. A potential relationship between skin hydration and stamp-type microneedle intradermal hyaluronic acid injection in middle-aged male face. J. Cosmet. Dermatol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Eklouh-Molinier, C.; Gaydou, V.; Froigneux, E.; Barlier, P.; Couturaud, V.; Manfait, M.; Piot, O. In vivo confocal Raman microspectroscopy of the human skin: Highlighting of spectral markers associated to aging via a research of correlation between Raman and biometric mechanical measurements. Anal. Bioanal. Chem. 2015, 407, 8363–8372. [Google Scholar] [CrossRef] [PubMed]
- Franzen, L.; Windbergs, M. Applications of Raman spectroscopy in skin research—From skin physiology and diagnosis up to risk assessment and dermal drug delivery. Adv. Drug Deliv. Rev. 2015, 89, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, T.M.; Wilhelm, K.P. A review of ageing and an examination of clinical methods in the assessment of ageing skin. Part I: Cellular and molecular perspectives of skin ageing. Int. J. Cosmet. Sci. 2008, 30, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Kottner, J.; Schario, M.; Garcia Bartels, N.; Pantchechnikova, E.; Hillmann, K.; Blume-Peytavi, U. Comparison of two in vivo measurements for skin surface topography. Skin Res. Technol. 2013, 19, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Trojahn, C.; Schario, M.; Dobos, G.; Blume-Peytavi, U.; Kottner, J. Reliability and validity of two in vivo measurements for skin surface topography in aged adults. Skin Res. Technol. 2015, 21, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Myer, K.; Maibach, H. Stratum corneum evaluation methods: Overview. Skin Res. Technol. 2013, 19, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Savory, S.A.; Agim, N.G.; Mao, R.; Peter, S.; Wang, C.; Maldonado, G.; Bearden Dietert, J.; Lieu, T.J.; Pretzlaff, K.; Das, S.; et al. Reliability assessment and validation of the postacne hyperpigmentation index (PAHPI), a new instrument to measure postinflammatory hyperpigmentation from acne vulgaris. J. Am. Acad. Dermatol. 2014, 70, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Gniadecka, M.; Jemec, G.B. Quantitative evaluation of chronological ageing and photoageing in vivo: Studies on skin echogenicity and thickness. Br. J. Dermatol. 1998, 139, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Irfan-Maqsood, M.; Matin, M.M.; Heirani-Tabasi, A.; Bahrami, M.; Naderi-Meshkin, H.; Mirahmadi, M.; Hassanzadeh, H.; Sanjar Moussavi, N.; Raza-Shah, H.; Raeesolmohaddeseen, M.; et al. Adipose derived mesenchymal stem cells express keratinocyte lineage markers in a co-culture model. Cell Mol. Biol. 2016, 62, 44–54. [Google Scholar] [PubMed]
- Hanel, K.H.; Cornelissen, C.; Luscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulukian, A.; Holland, A.J.; Vitre, B.; Naik, S.; Cleveland, D.W.; Fuchs, E. Epidermal development, growth control, and homeostasis in the face of centrosome amplification. Proc. Natl. Acad. Sci. USA 2015, 112, E6311–E6320. [Google Scholar] [CrossRef] [PubMed]
- Pesenacker, A.M.; Broady, R.; Levings, M.K. Control of tissue-localized immune responses by human regulatory T cells. Eur. J. Immunol. 2015, 45, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Mesa, K.R.; Kawaguchi, K.; Park, S.; Gonzalez, D.; Brown, S.; Boucher, J.; Klein, A.M.; Greco, V. Spatiotemporal coordination of stem cell commitment during epidermal homeostasis. Science 2016, 352, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Sayedyahossein, S.; Rudkouskaya, A.; Leclerc, V.; Dagnino, L. Integrin-linked kinase is indispensable for keratinocyte differentiation and epidermal barrier function. J. Investig. Dermatol. 2016, 136, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, R.E.; Rostan, E.F. Reversal of photodamage with topical growth factors: A pilot study. J. Cosmet. Laser Ther. 2003, 5, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Gold, M.H.; Goldman, M.P.; Biron, J. Efficacy of novel skin cream containing mixture of human growth factors and cytokines for skin rejuvenation. J. Drugs Dermatol. 2007, 6, 197–201. [Google Scholar] [PubMed]
- Ehrlich, M.; Rao, J.; Pabby, A.; Goldman, M.P. Improvement in the appearance of wrinkles with topical transforming growth factor β(1) and l-ascorbic acid. Dermatol. Surg. 2006, 32, 618–625. [Google Scholar] [PubMed]
- Lee, H.J.; Lee, E.G.; Kang, S.; Sung, J.H.; Chung, H.M.; Kim, D.H. Efficacy of microneedling plus human stem cell conditioned medium for skin rejuvenation: A randomized, controlled, blinded split-face study. Ann. Dermatol. 2014, 26, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Park, S.H.; Ahn, S.J.; Kim, H.K.; Park, J.S.; Lee, G.Y.; Kim, K.J.; Whang, K.K.; Kang, S.H.; Park, B.S.; et al. Whitening effect of adipose-derived stem cells: A critical role of TGF-β 1. Biol. Pharm. Bull. 2008, 31, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Park, B.S.; Park, S.H.; Kim, H.K.; Sung, J.H. Antiwrinkle effect of adipose-derived stem cell: Activation of dermal fibroblast by secretory factors. J. Dermatol. Sci. 2009, 53, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.Y.; Kim, D.H.; Lee, S.E.; Yoon, M.S.; Lee, H.J. Skin rejuvenation by microneedle fractional radiofrequency and a human stem cell conditioned medium in Asian skin: A randomized controlled investigator blinded split-face study. J. Cosmet. Laser Ther. 2013, 15, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Kim, J.; Lee, K.I.; Shin, J.M.; Chae, J.I.; Chung, H.M. Enhancement of wound healing by secretory factors of endothelial precursor cells derived from human embryonic stem cells. Cytotherapy 2011, 13, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Marikovsky, M.; Breuing, K.; Liu, P.Y.; Eriksson, E.; Higashiyama, S.; Farber, P.; Abraham, J.; Klagsbrun, M. Appearance of heparin-binding EGF-like growth factor in wound fluid as a response to injury. Proc. Natl. Acad. Sci. USA 1993, 90, 3889–3893. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Duan, D.S.; de Vries, C.; Peters, K.G.; Johnson, D.E.; Williams, L.T. Differential splicing in the extracellular region of fibroblast growth factor receptor 1 generates receptor variants with different ligand-binding specificities. Mol. Cell. Biol. 1992, 12, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.A.; Whang, J.L.; Tumolo, A.; Mergia, A.; Friedman, J.; Gospodarowicz, D.; Fiddes, J.C. Human basic fibroblast growth factor: Nucleotide sequence and genomic organization. EMBO J. 1986, 5, 2523–2528. [Google Scholar] [PubMed]
- Eriksson, A.; Siegbahn, A.; Westermark, B.; Heldin, C.H.; Claesson-Welsh, L. PDGF α- and β-receptors activate unique and common signal transduction pathways. EMBO J. 1992, 11, 543–550. [Google Scholar] [PubMed]
- Rappolee, D.A.; Mark, D.; Banda, M.J.; Werb, Z. Wound macrophages express TGF-α and other growth factors in vivo: Analysis by mRNA phenotyping. Science 1988, 241, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.R.; Platz, K.P.; Nalesnik, M.A.; Langrehr, J.M.; Neuhaus, P.; Schraut, W.H. Small bowel preservation causes basement membrane and endothelial cell injury. Transp. Proc. 1994, 26, 1498–1500. [Google Scholar]
- Madlener, M.; Mauch, C.; Conca, W.; Brauchle, M.; Parks, W.C.; Werner, S. Regulation of the expression of stromelysin-2 by growth factors in keratinocytes: Implications for normal and impaired wound healing. Biochem. J. 1996, 320, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Madlener, M.; Werner, S. Transforming growth factors β1, β2, and β3 and their receptors are differentially regulated during normal and impaired wound healing. J. Biol. Chem. 1996, 271, 10188–10193. [Google Scholar] [PubMed]
- Shah, M.; Foreman, D.M.; Ferguson, M.W. Neutralisation of TGF-β1 and TGF-β2 or exogenous addition of TGF-β3 to cutaneous rat wounds reduces scarring. J. Cell Sci. 1995, 108, 985–1002. [Google Scholar] [PubMed]
- Hubner, G.; Brauchle, M.; Smola, H.; Madlener, M.; Fassler, R.; Werner, S. Differential regulation of pro-inflammatory cytokines during wound healing in normal and glucocorticoid-treated mice. Cytokine 1996, 8, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Hubner, G.; Werner, S. Serum growth factors and proinflammatory cytokines are potent inducers of activin expression in cultured fibroblasts and keratinocytes. Exp. Cell Res. 1996, 228, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Leung, G.; Wang, A.; Fernando, M.; Phan, V.C.; McKay, D.M. Bone marrow-derived alternatively activated macrophages reduce colitis without promoting fibrosis: Participation of IL-10. American journal of physiology. Gastrointest. Liver Physiol. 2013, 304, G781–G792. [Google Scholar] [CrossRef] [PubMed]
- Peranteau, W.H.; Zhang, L.; Muvarak, N.; Badillo, A.T.; Radu, A.; Zoltick, P.W.; Liechty, K.W. IL-10 overexpression decreases inflammatory mediators and promotes regenerative healing in an adult model of scar formation. J. Investig. Dermatol. 2008, 128, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Thal, M.; Verma, S.; Hoxha, E.; Lambers, E.; Ramirez, V.; Qin, G.; Losordo, D.; Kishore, R. Interleukin-10 deficiency impairs bone marrow-derived endothelial progenitor cell survival and function in ischemic myocardium. Circ. Res. 2011, 109, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Balaji, S.; Le, L.D.; Crombleholme, T.M.; Keswani, S.G. Regenerative Wound Healing: The Role of Interleukin-10. Adv. Wound Care 2014, 3, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Mattey, D.L. Interleukin-4 induces myofibroblast differentiation is synovial fibroblasts. Biochem. Soc. Trans. 1997, 25, 290S. [Google Scholar] [CrossRef] [PubMed]
- Salmon-Ehr, V.; Serpier, H.; Nawrocki, B.; Gillery, P.; Clavel, C.; Kalis, B.; Birembaut, P.; Maquart, F.X. Expression of interleukin-4 in scleroderma skin specimens and scleroderma fibroblast cultures. Potential role in fibrosis. Arch. Dermatol. 1996, 132, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Lacraz, S.; Nicod, L.; Galve-de Rochemonteix, B.; Baumberger, C.; Dayer, J.M.; Welgus, H.G. Suppression of metalloproteinase biosynthesis in human alveolar macrophages by interleukin-4. J. Clin. Investig. 1992, 90, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Serpier, H.; Gillery, P.; Salmon-Ehr, V.; Garnotel, R.; Georges, N.; Kalis, B.; Maquart, F.X. Antagonistic effects of interferon-gamma and interleukin-4 on fibroblast cultures. J. Investig. Dermatol. 1997, 109, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Salmon-Ehr, V.; Ramont, L.; Godeau, G.; Birembaut, P.; Guenounou, M.; Bernard, P.; Maquart, F.X. Implication of interleukin-4 in wound healing. Lab. Investig. J. Tech. Methods Pathol. 2000, 80, 1337–1343. [Google Scholar] [CrossRef]
- Matias, M.A.; Saunus, J.M.; Ivanovski, S.; Walsh, L.J.; Farah, C.S. Accelerated wound healing phenotype in Interleukin 12/23 deficient mice. J. Inflamm. 2011, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, M.; Nakano, M.; Terabe, F.; Kawahata, H.; Ohkawara, T.; Han, Y.; Ripley, B.; Serada, S.; Nishikawa, T.; Kimura, A.; et al. The influence of excessive IL-6 production in vivo on the development and function of FOXP3+ regulatory T cells. J. Immunol. 2011, 186, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Jeong, H.J.; Kim, M.K.; Wee, W.R. Bone marrow-derived mesenchymal stem cells affect immunologic profiling of interleukin-17-secreting cells in a chemical burn mouse model. Korean J. Ophthalmol. 2014, 28, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Stallmeyer, B.; Kampfer, H.; Kolb, N.; Pfeilschifter, J. Leptin enhances wound re-epithelialization and constitutes a direct function of leptin in skin repair. J. Clin. Investig. 2000, 106, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Torsney, E.; Charlton, R.; Parums, D.; Collis, M.; Arthur, H.M. Inducible expression of human endoglin during inflammation and wound healing in vivo. Inflamm. Res. 2002, 51, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Tada, Y.; Hau, C.S.; Mitsui, A.; Kamata, M.; Asano, Y.; Sugaya, M.; Kadono, T.; Masamoto, Y.; Kurokawa, M.; et al. Adiponectin regulates psoriasiform skin inflammation by suppressing IL-17 production from gammadelta-T cells. Nat. Commun. 2015, 6, 7687. [Google Scholar] [CrossRef] [PubMed]
- Qa’aty, N.; Vincent, M.; Wang, Y.; Wang, A.; Mitts, T.F.; Hinek, A. Synthetic ligands of the elastin receptor induce elastogenesis in human dermal fibroblasts via activation of their IGF-1 receptors. J. Dermatol. Sci. 2015, 80, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Prokop, I.; Kononczuk, J.; Surazynski, A.; Palka, J. Cross-talk between integrin receptor and insulin-like growth factor receptor in regulation of collagen biosynthesis in cultured fibroblasts. Adv. Med. Sci. 2013, 58, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Noordam, R.; Gunn, D.A.; Tomlin, C.C.; Maier, A.B.; Griffiths, T.; Catt, S.D.; Ogden, S.; Slagboom, P.E.; Westendorp, R.G.; Griffiths, C.E.; et al. Serum insulin-like growth factor 1 and facial ageing: High levels associate with reduced skin wrinkling in a cross-sectional study. Br. J. Dermatol. 2013, 168, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Seamark, R.F. Granulocyte macrophage colony stimulating factor (GM-CSF) in the murine reproductive tract: Stimulation by seminal factors. Reprod. Fertil. Dev. 1990, 2, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Haisa, M.; Igarashi, A.; Pencev, D.; Grotendorst, G.R. Leukocyte-derived growth factor links the PDGF and CXC chemokine families of peptides. FASEB J. 1996, 10, 1336–1345. [Google Scholar] [PubMed]
- Schmitt, S.; Safferling, K.; Westphal, K.; Hrabowski, M.; Muller, U.; Angel, P.; Wiechert, L.; Ehemann, V.; Muller, B.; Holland-Cunz, S.; et al. Stathmin regulates keratinocyte proliferation and migration during cutaneous regeneration. PLoS ONE 2013, 8, e75075. [Google Scholar] [CrossRef]
- Bevan, D.; Gherardi, E.; Fan, T.P.; Edwards, D.; Warn, R. Diverse and potent activities of HGF/SF in skin wound repair. J. Pathol. 2004, 203, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.; Liotiri, S.; Bodo, E.; Hagen, E.; Biro, T.; Arck, P.C.; Paus, R. Probing the effects of stress mediators on the human hair follicle: Substance P holds central position. Am. J. Pathol. 2007, 171, 1872–1886. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Tang, Y.; Fang, H.; Su, Z.; Xu, B.; Lin, Y.; Zhang, P.; Wei, X. Decellularized scaffolds containing hyaluronic acid and EGF for promoting the recovery of skin wounds. J. Mater. Sci. Mater. Med. 2015, 26, 5322. [Google Scholar] [CrossRef] [PubMed]
- Gainza, G.; Bonafonte, D.C.; Moreno, B.; Aguirre, J.J.; Gutierrez, F.B.; Villullas, S.; Pedraz, J.L.; Igartua, M.; Hernandez, R.M. The topical administration of rhEGF-loaded nanostructured lipid carriers (rhEGF-NLC) improves healing in a porcine full-thickness excisional wound model. J. Control. Release 2015, 197, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, M.; Shabani, I.; Ahvaz, H.H.; Soleimani, M. PLGA/gelatin hybrid nanofibrous scaffolds encapsulating EGF for skin regeneration. J. Biomed. Mater. Res. Part A 2015, 103, 2225–2235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, L.; Wang, X.; Chen, S.; Jia, Q.; Liu, N.; Chen, Y.; Kong, Y.; Zhang, L.; Zhang, A.L. Activin B promotes BM-MSC-mediated cutaneous wound healing by regulating cell migration via the JNK-ERK signaling pathway. Cell Transp. 2013. [Google Scholar] [CrossRef]
- Zaragosi, L.E.; Wdziekonski, B.; Villageois, P.; Keophiphath, M.; Maumus, M.; Tchkonia, T.; Bourlier, V.; Mohsen-Kanson, T.; Ladoux, A.; Elabd, C.; et al. Activin A plays a critical role in proliferation and differentiation of human adipose progenitors. Diabetes 2010, 59, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Moura, J.; da Silva, L.; Cruz, M.T.; Carvalho, E. Molecular and cellular mechanisms of bone morphogenetic proteins and activins in the skin: Potential benefits for wound healing. Arch. Dermatol. Res. 2013, 305, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Dani, C. Activins in adipogenesis and obesity. Int. J. Obes. 2013, 37, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Gragnani, A.; Cezillo, M.V.; da Silva, I.D.; de Noronha, S.M.; Correa-Noronha, S.A.; Ferreira, L.M. Gene expression profile of cytokines and receptors of inflammation from cultured keratinocytes of burned patients. Burns J. Int. Soc. Burn Inj. 2014, 40, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Kobayashi, T.; Sugihara, E.; Yamada, T.; Ikuta, K.; Pittaluga, S.; Saya, H.; Amagai, M.; Nagao, K. Hair follicle-derived IL-7 and IL-15 mediate skin-resident memory T cell homeostasis and lymphoma. Nat. Med. 2015, 21, 1272–1279. [Google Scholar] [CrossRef] [PubMed]
- Knipper, J.A.; Willenborg, S.; Brinckmann, J.; Bloch, W.; Maass, T.; Wagener, R.; Krieg, T.; Sutherland, T.; Munitz, A.; Rothenberg, M.E.; et al. Interleukin-4 Receptor α Signaling in Myeloid Cells Controls Collagen Fibril Assembly in Skin Repair. Immunity 2015, 43, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.D.; MacNeil, L.G.; Lally, J.S.; Ford, R.J.; Bujak, A.L.; Brar, I.K.; Kemp, B.E.; Raha, S.; Steinberg, G.R.; Tarnopolsky, M.A. Exercise-stimulated interleukin-15 is controlled by AMPK and regulates skin metabolism and aging. Aging Cell 2015, 14, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Jeon, B.J.; Hwang, N.H.; Kim, M.S.; Park, S.H.; Dhong, E.S.; Yoon, E.S.; Lee, B.I. Adipose-derived stem cells inhibit epidermal melanocytes through an interleukin-6-mediated mechanism. Plast. Reconstr. Surg. 2014, 134, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Grunler, J.; Brismar, K.; Wang, J. Effects of IGFBP-1 and IGFBP-2 and their fragments on migration and IGF-induced proliferation of human dermal fibroblasts. Growth Horm. IGF Res. 2015, 25, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Song, H.J.; Lee, S.H.; Lee, C.K. Comparative study of various growth factors and cytokines on type I collagen and hyaluronan production in human dermal fibroblasts. J. Cosmet. Dermatol. 2014, 13, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Tzellos, T.G.; Klagas, I.; Vahtsevanos, K.; Triaridis, S.; Printza, A.; Kyrgidis, A.; Karakiulakis, G.; Zouboulis, C.C.; Papakonstantinou, E. Extrinsic ageing in the human skin is associated with alterations in the expression of hyaluronic acid and its metabolizing enzymes. Exp. Dermatol. 2009, 18, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, A.; Yoshida, H.; Nakamura, S.; Morikawa, T.; Kawabata, K.; Kobayashi, M.; Sakai, S.; Takahashi, Y.; Okada, Y.; Inoue, S. Regulation of Hyaluronan (HA) Metabolism Mediated by HYBID (Hyaluronan-binding Protein Involved in HA Depolymerization, KIAA1199) and HA Synthases in Growth Factor-stimulated Fibroblasts. J. Biol. Chem. 2015, 290, 30910–30923. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Sanchez, M.; Zalduendo, M.M.; de la Fuente, M.; Prado, R.; Orive, G.; Andia, I. Fibroblastic response to treatment with different preparations rich in growth factors. Cell Prolif. 2009, 42, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Poimenidi, E.; Theodoropoulou, C.; Koutsioumpa, M.; Skondra, L.; Droggiti, E.; van den Broek, M.; Koolwijk, P.; Papadimitriou, E. Vascular endothelial growth factor A (VEGF-A) decreases expression and secretion of pleiotrophin in a VEGF receptor-independent manner. Vasc. Pharmacol. 2016, 80, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Rezza, A.; Sennett, R.; Tanguy, M.; Clavel, C.; Rendl, M. PDGF signalling in the dermis and in dermal condensates is dispensable for hair follicle induction and formation. Exp. Dermatol. 2015, 24, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Ezure, T.; Amano, S. Adiponectin and leptin up-regulate extracellular matrix production by dermal fibroblasts. BioFactors 2007, 31, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, S.; Ide, S.; Tokuyama, R.; Umeki, H.; Tatehara, S.; Kataoka, S.; Satomura, K. Leptin promotes wound healing in the skin. PLoS ONE 2015, 10, e0121242. [Google Scholar] [CrossRef] [PubMed]
- Kypriotou, M.; Rivero, D.; Haller, S.; Mariotto, A.; Huber, M.; Acha-Orbea, H.; Werner, S.; Hohl, D. Activin A inhibits antigen-induced allergy in murine epicutaneous sensitization. Front. Immunol. 2013, 4, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugrillon, A.; Eichler, H.; Kern, S.; Kluter, H. Autologous concentrated platelet-rich plasma (cPRP) for local application in bone regeneration. Int. J. Oral Maxillofac. Surg. 2002, 31, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Rendu, F.; Brohard-Bohn, B. The platelet release reaction: Granules’ constituents, secretion and functions. Platelets 2001, 12, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Hassan, W.U.; Greiser, U.; Wang, W. Role of adipose-derived stem cells in wound healing. Wound Repair Regen. 2014, 22, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Ojeh, N.; Pastar, I.; Tomic-Canic, M.; Stojadinovic, O. Stem Cells in Skin Regeneration, Wound Healing, and Their Clinical Applications. Int. J. Mol. Sci. 2015, 16, 25476–25501. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.F. Neutrophil activation on biological surfaces. Massive secretion of hydrogen peroxide in response to products of macrophages and lymphocytes. J. Clin. Investig. 1987, 80, 1550–1560. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [PubMed]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.E.; Alonso, D.F.; Yoshiji, H.; Thorgeirsson, U.P. Tissue inhibitors of metalloproteinases: Structure, regulation and biological functions. Eur. J. Cell Biol. 1997, 74, 111–122. [Google Scholar] [PubMed]
- Fisher, G.J.; Varani, J.; Voorhees, J.J. Looking older: Fibroblast collapse and therapeutic implications. Arch. Dermatol. 2008, 144, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Rittie, L.; Berton, A.; Monboisse, J.C.; Hornebeck, W.; Gillery, P. Decreased contraction of glycated collagen lattices coincides with impaired matrix metalloproteinase production. Biochem. Biophys. Res. Commun. 1999, 264, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Werner, S.; Bugnon, P.; Wickenhauser, C.; Siewe, L.; Utermohlen, O.; Davidson, J.M.; Krieg, T.; Roers, A. Accelerated wound closure in mice deficient for interleukin-10. Am. J. Pathol. 2007, 170, 188–202. [Google Scholar] [CrossRef] [PubMed]
- McCartney-Francis, N.; Mizel, D.; Wong, H.; Wahl, L.; Wahl, S. TGF-β regulates production of growth factors and TGF-β by human peripheral blood monocytes. Growth Factors 1990, 4, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Kiritsy, C.P.; Lynch, A.B.; Lynch, S.E. Role of growth factors in cutaneous wound healing: A review. Crit. Rev. Oral Biol. Med. 1993, 4, 729–760. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.B.; Reichner, J.S.; Albina, J.E. Interleukin-6 activity in wounds. Am. J. Physiol. 1994, 266, R1840–R1844. [Google Scholar] [PubMed]
- Moulin, V. Growth factors in skin wound healing. Eur. J. Cell Biol. 1995, 68, 1–7. [Google Scholar] [PubMed]
- Nishikawa, K.; Asai, T.; Shigematsu, H.; Shimizu, K.; Kato, H.; Asano, Y.; Takashima, S.; Mekada, E.; Oku, N.; Minamino, T. Development of anti-HB-EGF immunoliposomes for the treatment of breast cancer. J. Control. Release 2012, 160, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Yun, M.; Paik, J.S.; Lee, S.B.; Yang, S.W. PDGF-BB Enhances the Proliferation of Cells in Human Orbital Fibroblasts by Suppressing PDCD4 Expression via Up-Regulation of microRNA-21. Investig. Ophthalmol. Vis. Sci. 2016, 57, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Scholz, F.; Schulte, A.; Adamski, F.; Hundhausen, C.; Mittag, J.; Schwarz, A.; Kruse, M.L.; Proksch, E.; Ludwig, A. Constitutive expression and regulated release of the transmembrane chemokine CXCL16 in human and murine skin. J. Investig. Dermatol. 2007, 127, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.J.; Jiang, D.G.; Jiang, W.G.; Harding, K.G.; Patel, G.K. Activated leukocyte cell adhesion molecule impacts on clinical wound healing and inhibits HaCaT migration. Int. Wound J. 2011, 8, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ren, G.S.; Li, F.; Sun, S.Q. Expression of livin and vascular endothelial growth factor in different clinical stages of human esophageal carcinoma. World J. Gastroenterol. 2008, 14, 5749–5754. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, A.; Lampugnani, M.G.; Balconi, G.; Breviario, F.; Corada, M.; Lanfrancone, L.; Dejana, E. Vascular endothelial growth factor induces SHC association with vascular endothelial cadherin: A potential feedback mechanism to control vascular endothelial growth factor receptor-2 signaling. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Chung, P.S.; Ahn, J.C. Adipose-derived stromal cell cluster with light therapy enhance angiogenesis and skin wound healing in mice. Biochem. Biophys. Res. Commun. 2015, 462, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, H.; Pfeilschifter, J.; Frank, S. Expressional regulation of angiopoietin-1 and -2 and the tie-1 and -2 receptor tyrosine kinases during cutaneous wound healing: A comparative study of normal and impaired repair. Lab. Investig. J. Tech. Methods Pathol. 2001, 81, 361–373. [Google Scholar] [CrossRef]
- Staton, C.A.; Valluru, M.; Hoh, L.; Reed, M.W.; Brown, N.J. Angiopoietin-1, angiopoietin-2 and Tie-2 receptor expression in human dermal wound repair and scarring. Br. J. Dermatol. 2010, 163, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Joao de Masi, E.C.; Campos, A.C.; Joao de Masi, F.D.; Ratti, M.A.; Ike, I.S.; Joao de Mais, R.D. The influence of growth factors on skin wound healing in rats. Braz. J. Otorhinolaryngol. 2016, 82, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Peplow, P.V.; Chatterjee, M.P. A review of the influence of growth factors and cytokines in in vitro human keratinocyte migration. Cytokine 2013, 62, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Krieg, T.; Smola, H. Keratinocyte-fibroblast interactions in wound healing. J. Investig. Dermatol. 2007, 127, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Ansel, J.; Perry, P.; Brown, J.; Damm, D.; Phan, T.; Hart, C.; Luger, T.; Hefeneider, S. Cytokine modulation of keratinocyte cytokines. J. Investig. Dermatol. 1990, 94 (Suppl. S6), 101S–107S. [Google Scholar] [CrossRef] [PubMed]
- Finch, P.W.; Rubin, J.S.; Miki, T.; Ron, D.; Aaronson, S.A. Human KGF is FGF-related with properties of a paracrine effector of epithelial cell growth. Science 1989, 245, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Grossman, R.M.; Krueger, J.; Yourish, D.; Granelli-Piperno, A.; Murphy, D.P.; May, L.T.; Kupper, T.S.; Sehgal, P.B.; Gottlieb, A.B. Interleukin 6 is expressed in high levels in psoriatic skin and stimulates proliferation of cultured human keratinocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 6367–6371. [Google Scholar] [CrossRef] [PubMed]
- Freedberg, I.M.; Tomic-Canic, M.; Komine, M.; Blumenberg, M. Keratins and the keratinocyte activation cycle. J. Investig. Dermatol. 2001, 116, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Babu, P.S.; Danilovich, N.; Sairam, M.R. Hormone-induced receptor gene splicing: Enhanced expression of the growth factor type I follicle-stimulating hormone receptor motif in the developing mouse ovary as a new paradigm in growth regulation. Endocrinology 2001, 142, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Florin, L.; Maas-Szabowski, N.; Werner, S.; Szabowski, A.; Angel, P. Increased keratinocyte proliferation by JUN-dependent expression of PTN and SDF-1 in fibroblasts. J. Cell Sci. 2005, 118, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Kapur, S.K.; Dos-Anjos Vilaboa, S.; Llull, R.; Katz, A.J. Adipose tissue and stem/progenitor cells: Discovery and development. Clin. Plast. Surg. 2015, 42, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Conde-Green, A.; Marano, A.A.; Lee, E.S.; Reisler, T.; Price, L.A.; Milner, S.M.; Granick, M.S. Fat Grafting and Adipose-Derived Regenerative Cells in Burn Wound Healing and Scarring: A Systematic Review of the Literature. Plast. Reconstr. Surg. 2016, 137, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Negenborn, V.L.; Groen, J.W.; Smit, J.M.; Niessen, F.B.; Mullender, M.G. The Use of Autologous Fat Grafting for Treatment of Scar Tissue and Scar-Related Conditions: A Systematic Review. Plast. Surg. Nurs. 2016, 36, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Shridharani, S.M.; Broyles, J.M.; Matarasso, A. Liposuction devices: Technology update. Med. Devices 2014, 7, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar] [PubMed]
- Banyard, D.A.; Salibian, A.A.; Widgerow, A.D.; Evans, G.R. Implications for human adipose-derived stem cells in plastic surgery. J. Cell. Mol. Med. 2015, 19, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Aronowitz, J.A.; Lockhart, R.A.; Hakakian, C.S. Mechanical versus enzymatic isolation of stromal vascular fraction cells from adipose tissue. SpringerPlus 2015, 4, 713. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, E.C.; Albano, N.J.; Hazen, A. Roll, Spin, Wash, or Filter? Processing of Lipoaspirate for Autologous Fat Grafting: An Updated, Evidence-Based Review of the Literature. Plast. Reconstr. Surg. 2015, 136, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Conde-Green, A.; Kotamarti, V.S.; Sherman, L.S.; Keith, J.D.; Lee, E.S.; Granick, M.S.; Rameshwar, P. Shift toward Mechanical Isolation of Adipose-derived Stromal Vascular Fraction: Review of Upcoming Techniques. Plastic and reconstructive surgery. Glob. Open 2016, 4, e1017. [Google Scholar] [CrossRef] [PubMed]
- Hanke, A.; Prantl, L.; Wenzel, C.; Nerlich, M.; Brockhoff, G.; Loibl, M.; Gehmert, S. Semi-automated extraction and characterization of Stromal Vascular Fraction using a new medical device. Clin. Hemorheol. Microcirc. 2016. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.A.; Stevens, H.P.; Parvizi, M.; van der Lei, B.; Harmsen, M.C. The fractionation of adipose tissue procedure to obtain stromal vascular fractions for regenerative purposes. Wound Repair Regen. 2016, 24, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Grasys, J.; Kim, B.S.; Pallua, N. Content of Soluble Factors and Characteristics of Stromal Vascular Fraction Cells in Lipoaspirates from Different Subcutaneous Adipose Tissue Depots. Aesthet. Surg. J. 2016, 36, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Kakagia, D.; Pallua, N. Autologous fat grafting: In search of the optimal technique. Surg. Innov. 2014, 21, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Strong, A.L.; Cederna, P.S.; Rubin, J.P.; Coleman, S.R.; Levi, B. The Current State of Fat Grafting: A Review of Harvesting, Processing, and Injection Techniques. Plast. Reconstr. Surg. 2015, 136, 897–912. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Wang, J.R.; Liao, X.; Li, S.H.; Xiao, L.L.; Cheng, B.; Xie, G.H.; Song, J.X.; Liu, H.W. Effect of suction pressures on cell yield and functionality of the adipose-derived stromal vascular fraction. J. Plast. Reconstr. Aesthet. Surg. 2016. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Nguyen, A.; Banyard, D.A.; Fadavi, D.; Toranto, J.D.; Wirth, G.A.; Paydar, K.Z.; Evans, G.R.; Widgerow, A.D. Stromal vascular fraction: A regenerative reality? Part 2: Mechanisms of regenerative action. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Streit, L.; Jaros, J.; Sedlakova, V.; Sedlackova, M.; Drazan, L.; Svoboda, M.; Pospisil, J.; Vyska, T.; Vesely, J.; Hampl, A. A Comprehensive In Vitro Comparison of Preparation Techniques for Fat Grafting. Plast. Reconstr. Surg. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Berry, D.C.; Tang, W.; Graff, J.M. Independent stem cell lineages regulate adipose organogenesis and adipose homeostasis. Cell Rep. 2014, 9, 1007–1022. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Guilak, F. Differentiation potential of adipose derived adult stem (ADAS) cells. Curr. Top. Dev. Biol. 2003, 58, 137–160. [Google Scholar] [PubMed]
- Katz, A.J.; Tholpady, A.; Tholpady, S.S.; Shang, H.; Ogle, R.C. Cell surface and transcriptional characterization of human adipose-derived adherent stromal (hADAS) cells. Stem Cells 2005, 23, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; de Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Strioga, M.; Viswanathan, S.; Darinskas, A.; Slaby, O.; Michalek, J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012, 21, 2724–2752. [Google Scholar] [CrossRef] [PubMed]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Wu, X.Y.; Tong, J.B.; Yang, X.X.; Zhao, J.L.; Zheng, Q.F.; Zhao, G.B.; Ma, Z.J. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Res. Ther. 2015, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Geesman, G.J.; Hostikka, S.L.; Atallah, M.; Blackwell, B.; Lee, E.; Cook, P.J.; Pasaniuc, B.; Shariat, G.; Halperin, E.; et al. Inhibition of activated pericentromeric SINE/Alu repeat transcription in senescent human adult stem cells reinstates self-renewal. Cell Cycle 2011, 10, 3016–3030. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Niu, P.; Wang, L.; Vogelsang, M.; Gaur, M.; Krastins, B.; Zhao, Y.; Smagul, A.; Nussupbekova, A.; Akanov, A.; et al. Opposing activities of oncogenic MIR17HG and tumor suppressive MIR100HG clusters and their gene targets regulate replicative senescence in human adult stem cells. Aging Mech. Dis. 2017, in press. [Google Scholar]
- Gruber, H.E.; Somayaji, S.; Riley, F.; Hoelscher, G.L.; Norton, H.J.; Ingram, J.; Hanley, E.N., Jr. Human adipose-derived mesenchymal stem cells: Serial passaging, doubling time and cell senescence. Biotech. Histochem. 2012, 87, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Jun, H.S.; Dao, L.T.; Pyun, J.C.; Cho, S. Effect of cell senescence on the impedance measurement of adipose tissue-derived stem cells. Enzym. Microb. Technol. 2013, 53, 302–306. [Google Scholar] [CrossRef] [PubMed]
- von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef]
- Jeyapalan, J.C.; Sedivy, J.M. Cellular senescence and organismal aging. Mech. Ageing Dev. 2008, 129, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J. Cellular senescence as a tumor-suppressor mechanism. Trends Cell Biol. 2001, 11, S27–S31. [Google Scholar] [CrossRef]
- Lin, A.W.; Barradas, M.; Stone, J.C.; van Aelst, L.; Serrano, M.; Lowe, S.W. Premature senescence involving p53 and p16 is activated in response to constitutive MEK/MAPK mitogenic signaling. Genes Dev. 1998, 12, 3008–3019. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Bayliss, P.E.; Hanai, J. A prospective epigenetic paradigm between cellular senescence and epithelial-mesenchymal transition in organismal development and aging. Transl. Res. J. Lab. Clin. Med. 2015, 165, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Agostini, T.; Lazzeri, D.; Pini, A.; Marino, G.; Li Quattrini, A.; Bani, D.; Dini, M. Wet and dry techniques for structural fat graft harvesting: Histomorphometric and cell viability assessments of lipoaspirated samples. Plast. Reconstr. Surg. 2012, 130, 331e–339e. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.R.; Katzel, E.B. Fat Grafting for Facial Filling and Regeneration. Clin. Plast. Surg. 2015, 42, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Marten, T.J.; Elyassnia, D. Fat grafting in facial rejuvenation. Clin. Plast. Surg. 2015, 42, 219–252. [Google Scholar] [CrossRef] [PubMed]
- Sinno, S.; Mehta, K.; Simmons, C.; Stuzin, J.M. Current Trends in Facial Rejuvenation: An Assessment of ASPS Members Utilization of Fat Grafting During Facelifting. Plast. Reconstr. Surg. 2015, 136 (Suppl. S4), 141. [Google Scholar] [CrossRef] [PubMed]
- Maione, L.; Memeo, A.; Pedretti, L.; Verdoni, F.; Lisa, A.; Bandi, V.; Giannasi, S.; Vinci, V.; Mambretti, A.; Klinger, M. Autologous fat graft as treatment of post short stature surgical correction scars. Injury 2014, 45 (Suppl. S6), S126–S132. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, F.P.; Gennai, A.; Izzo, L.; Zambelli, A.; Repaci, E.; Baldelli, I.; Fraternali-Orcioni, G.; Hartstein, M.E.; Santi, P.L.; Quarto, R. Superficial Enhanced Fluid Fat Injection (SEFFI) to Correct Volume Defects and Skin Aging of the Face and Periocular Region. Aesthet. Surg. J. 2015, 35, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Giunta, R.E.; Eder, M.; Machens, H.G.; Muller, D.F.; Kovacs, L. Structural fat grafting for rejuvenation of the dorsum of the hand. Handchir. Mikrochir. Plast. Chir. 2010, 42, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Luu, C.A.; Larson, E.; Rankin, T.M.; Pappalardo, J.L.; Slepian, M.J.; Armstrong, D.G. Plantar Fat Grafting and Tendon Balancing for the Diabetic Foot Ulcer in Remission. Plast. Reconstr. Surg. Glob. Open 2016, 4, e810. [Google Scholar] [CrossRef] [PubMed]
- Zvonic, S.; Lefevre, M.; Kilroy, G.; Floyd, Z.E.; DeLany, J.P.; Kheterpal, I.; Gravois, A.; Dow, R.; White, A.; Wu, X.; et al. Secretome of primary cultures of human adipose-derived stem cells: Modulation of serpins by adipogenesis. Mol. Cell. Proteom. 2007, 6, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Jansma, J.; Schepers, R.H.; Vissink, A. Microfat transfer in cosmetic facial procedures. Ned. Tijdschr. Tandheelk. 2014, 121, 330–335. [Google Scholar] [CrossRef]
- Kemaloglu, C.A. Nanofat grafting under a split-thickness skin graft for problematic wound management. SpringerPlus 2016, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Modarressi, A. Platlet rich plasma (prp) improves fat grafting outcomes. World J Plast Surg 2013, 2, 6–13. [Google Scholar] [PubMed]
- Koh, K.S.; Oh, T.S.; Kim, H.; Chung, I.W.; Lee, K.W.; Lee, H.B.; Park, E.J.; Jung, J.S.; Shin, I.S.; Ra, J.C.; et al. Clinical application of human adipose tissue-derived mesenchymal stem cells in progressive hemifacial atrophy (Parry-Romberg disease) with microfat grafting techniques using 3-dimensional computed tomography and 3-dimensional camera. Ann. Plast. Surg. 2012, 69, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Karagianni, M.; Kraneburg, U.; Kluter, H.; Machens, H.G.; Bieback, K.; Schantz, J.T.; Papadopulos, N. Autologous fat grafts and supportive enrichment with adipose tissue stromal cells. Handchir. Mikrochir. Plast. Chir. 2013, 45, 93–98. [Google Scholar] [PubMed]
- Han, S.; Sun, H.M.; Hwang, K.C.; Kim, S.W. Adipose-derived stromal vascular fraction cells: Update on clinical utility and efficacy. Crit. Rev. Eukaryot. Gene Expr. 2015, 25, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.; Guo, J.; Banyard, D.A.; Fadavi, D.; Toranto, J.D.; Wirth, G.A.; Paydar, K.Z.; Evans, G.R.; Widgerow, A.D. Stromal vascular fraction: A regenerative reality? Part 1: Current concepts and review of the literature. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, G.; Marchi, A.; Galie, M.; Baroni, G.; Benati, D.; Krampera, M.; Pasini, A.; Sbarbati, A. Clinical treatment of radiotherapy tissue damage by lipoaspirate transplant: A healing process mediated by adipose-derived adult stem cells. Plast. Reconstr. Surg. 2007, 119, 1409–1422. [Google Scholar] [CrossRef] [PubMed]
- Toyserkani, N.M.; Christensen, M.L.; Sheikh, S.P.; Sorensen, J.A. Adipose-Derived Stem Cells: New Treatment for Wound Healing? Ann. Plast. Surg. 2015, 75, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Marques, L.F.; Stessuk, T.; Camargo, I.C.; Sabeh Junior, N.; dos Santos, L.; Ribeiro-Paes, J.T. Platelet-rich plasma (PRP): Methodological aspects and clinical applications. Platelets 2015, 26, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Stessuk, T.; Puzzi, M.B.; Chaim, E.A.; Alves, P.C.; de Paula, E.V.; Forte, A.; Izumizawa, J.M.; Oliveira, C.C.; Frei, F.; Ribeiro-Paes, J.T. Platelet-rich plasma (PRP) and adipose-derived mesenchymal stem cells: Stimulatory effects on proliferation and migration of fibroblasts and keratinocytes in vitro. Arch. Dermatol. Res. 2016, 308, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Son, W.C.; Yun, J.W.; Kim, B.H. Adipose-derived mesenchymal stem cells reduce MMP-1 expression in UV-irradiated human dermal fibroblasts: Therapeutic potential in skin wrinkling. Biosci. Biotechnol. Biochem. 2015, 79, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Dos Anjos, S.; Matas-Palau, A.; Mercader, J.; Katz, A.J.; Llull, R. Reproducible Volume Restoration and Efficient Long-term Volume Retention after Point-of-care Standardized Cell-enhanced Fat Grafting in Breast Surgery. Plast. Reconstr. Surg. Glob. Open 2015, 3, e547. [Google Scholar] [CrossRef] [PubMed]
- Wankhade, U.D.; Shen, M.; Kolhe, R.; Fulzele, S. Advances in Adipose-Derived Stem Cells Isolation, Characterization, and Application in Regenerative Tissue Engineering. Stem Cells Int. 2016, 2016, 3206807. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; de Angelis, B.; Pasin, M.; Cervelli, G.; Curcio, C.B.; Floris, M.; Di Pasquali, C.; Bocchini, I.; Balzani, A.; Nicoli, F.; et al. Adipose-derived stromal vascular fraction cells and platelet-rich plasma: Basic and clinical evaluation for cell-based therapies in patients with scars on the face. J. Craniofac. Surg. 2014, 25, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Trojahn Kolle, S.F.; Oliveri, R.S.; Glovinski, P.V.; Elberg, J.J.; Fischer-Nielsen, A.; Drzewiecki, K.T. Importance of mesenchymal stem cells in autologous fat grafting: A systematic review of existing studies. J. Plast. Surg. Hand Surg. 2012, 46, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Tonnard, P.; Verpaele, A.; Peeters, G.; Hamdi, M.; Cornelissen, M.; Declercq, H. Nanofat grafting: Basic research and clinical applications. Plast. Reconstr. Surg. 2013, 132, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Lee, H.J.; Lee, J.S.; Yoon, H.; Chun, W.; Kim, G.H. A novel cell-printing method and its application to hepatogenic differentiation of human adipose stem cell-embedded mesh structures. Sci. Rep. 2015, 5, 13427. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Coruh, A.; Deniz, K. Stromal vascular fraction improves deep partial thickness burn wound healing. Burns J. Int. Soc. Burn Inj. 2014, 40, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Foubert, P.; Barillas, S.; Gonzalez, A.D.; Alfonso, Z.; Zhao, S.; Hakim, I.; Meschter, C.; Tenenhaus, M.; Fraser, J.K. Uncultured adipose-derived regenerative cells (ADRCs) seeded in collagen scaffold improves dermal regeneration, enhancing early vascularization and structural organization following thermal burns. Burns J. Int. Soc. Burn Inj. 2015, 41, 1504–1516. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Sheng, L.; Li, H.; Mao, X.; Zhu, M.; Gao, B.; Li, Q. Cell-Assisted Skin Grafting: Improving Texture and Elasticity of Skin Grafts through Autologous Cell Transplantation. Plast. Reconstr. Surg. 2016, 137, 58e–66e. [Google Scholar] [CrossRef] [PubMed]
- Klar, A.S.; Guven, S.; Zimoch, J.; Zapiorkowska, N.A.; Biedermann, T.; Bottcher-Haberzeth, S.; Meuli-Simmen, C.; Martin, I.; Scherberich, A.; Reichmann, E.; et al. Characterization of vasculogenic potential of human adipose-derived endothelial cells in a three-dimensional vascularized skin substitute. Pediatr. Surg. Int. 2016, 32, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Brohem, C.A.; de Carvalho, C.M.; Radoski, C.L.; Santi, F.C.; Baptista, M.C.; Swinka, B.B.; de, A.U.C.; de Araujo, L.R.; Graf, R.M.; Feferman, I.H.; et al. Comparison between fibroblasts and mesenchymal stem cells derived from dermal and adipose tissue. Int. J. Cosmet. Sci. 2013, 35, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Tissiani, L.A.; Alonso, N. A Prospective and Controlled Clinical Trial on Stromal Vascular Fraction Enriched Fat Grafts in Secondary Breast Reconstruction. Stem Cells Int. 2016, 2016, 2636454. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Laliberte, M.; Blelloch, S.; Ratanshi, I.; Safneck, J.; Buchel, E.; Raouf, A. Adipose-Derived Stromal Vascular Fraction Differentially Expands Breast Progenitors in Tissue Adjacent to Tumors Compared to Healthy Breast Tissue. Plast. Reconstr. Surg. 2015, 136, 414e–425e. [Google Scholar] [CrossRef] [PubMed]
- Toyserkani, N.M.; Quaade, M.L.; Sorensen, J.A. Cell-Assisted Lipotransfer: A Systematic Review of Its Efficacy. Aesthet. Plast. Surg. 2016, 40, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Luo, X.; Lu, Y.; Fan, Z.H.; Hu, X. Is the Resorption of Grafted Fat Reduced in Cell-Assisted Lipotransfer for Breast Augmentation? Ann. Plast. Surg. 2015, 75, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Kozlik, M.; Wojcicki, P. The use of stem cells in plastic and reconstructive surgery. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2014, 23, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Minteer, D.; Marra, K.G.; Rubin, J.P. Adipose-derived mesenchymal stem cells: Biology and potential applications. Adv. Biochem. Eng. Biotechnol. 2013, 129, 59–71. [Google Scholar] [PubMed]
- Mestak, O.; Sukop, A.; Hsueh, Y.S.; Molitor, M.; Mestak, J.; Matejovska, J.; Zarubova, L. Centrifugation versus PureGraft for fatgrafting to the breast after breast-conserving therapy. World J. Surg. Oncol. 2014, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Kahn, C.R. Transplantation of adipose tissue and stem cells: Role in metabolism and disease. Nat. Rev. Endocrinol. 2010, 6, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, G.; Charles-de-Sa, L.; Gontijo-de-Amorim, N.F.; Takiya, C.M.; Amable, P.R.; Borojevic, R.; Benati, D.; Bernardi, P.; Sbarbati, A. Expanded Stem Cells, Stromal-Vascular Fraction, and Platelet-Rich Plasma Enriched Fat: Comparing Results of Different Facial Rejuvenation Approaches in a Clinical Trial. Aesthet. Surg. J. Am. Soc. Aesthet. Plast. Surg. 2016, 36, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Charles-de-Sa, L.; Gontijo-de-Amorim, N.F.; Maeda Takiya, C.; Borojevic, R.; Benati, D.; Bernardi, P.; Sbarbati, A.; Rigotti, G. Antiaging treatment of the facial skin by fat graft and adipose-derived stem cells. Plast. Reconstr. Surg. 2015, 135, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, H.; Suga, H. Hair Regeneration Treatment Using Adipose-Derived Stem Cell Conditioned Medium: Follow-up With Trichograms. Eplasty 2015, 15, e10. [Google Scholar] [PubMed]
- Asero, R.; Tedeschi, A.; Lorini, M.; Gerosa, M.; Meroni, P.; Riboldi, P. Circulating stem cell factor in patients with chronic idiopathic urticaria. Ann. Allergy Asthma Immunol. 2003, 91, 79–81. [Google Scholar] [CrossRef]
- Kesten, S.; Fraser, J.K. Autologous Adipose Derived Regenerative Cells: A Platform for Therapeutic Applications. Surg. Technol. Int. 2016, XXIX, 38–44. [Google Scholar]
- Granel, B.; Daumas, A.; Jouve, E.; Harle, J.R.; Nguyen, P.S.; Chabannon, C.; Colavolpe, N.; Reynier, J.C.; Truillet, R.; Mallet, S.; et al. Safety, tolerability and potential efficacy of injection of autologous adipose-derived stromal vascular fraction in the fingers of patients with systemic sclerosis: An open-label phase I trial. Ann. Rheum. Dis. 2015, 74, 2175–2182. [Google Scholar] [CrossRef] [PubMed]
- Liew, A.; O’Brien, T. Therapeutic potential for mesenchymal stem cell transplantation in critical limb ischemia. Stem Cell Res. Ther. 2012, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Lee, S.K.; Yoon, J.I.; Kim, D.E.; Kim, M.; Ha, H. Fibrin glue improves the therapeutic effect of MSCs by sustaining survival and paracrine function. Tissue Eng. Part A 2013, 19, 2373–2381. [Google Scholar] [CrossRef] [PubMed]
- Marston, W.A.; Hanft, J.; Norwood, P.; Pollak, R. The efficacy and safety of Dermagraft in improving the healing of chronic diabetic foot ulcers: Results of a prospective randomized trial. Diabetes Care 2003, 26, 1701–1705. [Google Scholar] [CrossRef] [PubMed]
- Claudio-da-Silva, C.; Baptista, L.S.; Carias, R.B.; Menezes Neto Hda, C.; Borojevic, R. Autologous mesenchymal stem cells culture from adipose tissue for treatment of facial rhytids. Rev. Col. Bras. Cir. 2009, 36, 288–291. [Google Scholar] [PubMed]
- Tollervey, J.R.; Lunyak, V.V. Adult stem cells: Simply a tool for regenerative medicine or an additional piece in the puzzle of human aging? Cell Cycle 2011, 10, 4173–4176. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Factor | Primary Target Cells and Effect | References |
|---|---|---|
| HB-EGF | Keratinocyte and fibroblast mitogen | [75] |
| FGFs 1, 2, and 4 | Angiogenic and fibroblast mitogen | [76,77] |
| PDGF | Chemotactic for macrophages, fibroblasts; macrophage activation; fibroblast mitogen, and matrix production | [78] |
| IGF-1 | Endothelial cell fibroblast mitogen | [79,80] |
| TGF-β1 and β2 | Keratinocyte migration; chemotactic for macrophages and fibroblasts | [81] |
| TGF-β3 | Wounded mouse skin, cutaneous rat wounds; Antiscarring | [82,83] |
| IL-1α and –β | Early activators of growth factor expression in macrophages, keratinocytes and fibroblasts; function in wound healing | [84,85] |
| IL-10 | Regenerative wound repair, Optimization of fibroblast and endothelial progenitor cells (EPC) function in differentiation, Attenuation of inflammatory response | [86,87,88,89] |
| IL-4 | Induction of fibroblast migration and differentiation, ECM production, control of MMP, Lower dermis maintenance, Wound healing | [90,91,92,93,94] |
| IL-12 | Epithelial cells under mouse tongue; Early inflammatory response and vascularization in wound healing, Regulation of Growth factors synthesis | [95] |
| IL-6, IL-8, IL-17a | Promote connection between immune cells and keratinocytes, fibroblasts Pro-inflammatory response, All the stages of wound healing, | [96,97] |
| LEP and LEPR | Keratinocytes proliferation, fibroblasts migration, Hyaluronic acid and collagen synthesis, Re-epithelialization processes | [32,98] |
| Endoglin | Human skin, lung and liver; Co-receptor for TGF-β, Promotes wound healing | [99] |
| Adipoq | Recruitment of fibroblasts and endothelial cells; Dermis regeneration, Wound healing, Suppression of inflammation | [32,100] |
| IGFBP1, IGFBP3 | Human dermal fibroblasts, human skin Collagen synthesis, Elastogenesis in dermal fibroblasts, Angiogenesis, Proliferation and differentiation | [101,102,103] |
| CSF1, CSF3 and receptor CSFR1 | Rat dermis Stimulation of TGFβ, Anti-inflammatory, Wound healing | Patent US8273712 B2 [104] |
| PPBP/NAP-2 | Fibroblast; mitogenic activity, Modulation of immune cells | [103,105] |
| HGF | Keratinocytes migration and differentiation; Scarless wound healing | [30,106,107] |
| NGFR | Human hair follicle cells, ECM remodeling, Wound healing, Modulation of immune cells | [108] |
| EGF | Rabbit skin cells, White pig skin, human fibroblasts, rat skin, Activation of dormant stem cells, Wound healing, Skin rejuvenation, Production of collagen, elastin, ECM remodeling | [33,109,110,111] |
| TNF-α | Mice epithelial cells, Similar to the IL-1 | [84] |
| Activin A and Activin B | Rat; Cutaneous wound healing, Human adipose progenitors proliferation and differentiation | [112,113,114,115] |
| Condition | Title | Trial Number | Status | References |
|---|---|---|---|---|
| Scars | A Phase I/II Study of Autologous Fat Transfer for Scar Prevention and Remodeling. | NCT01119326 | REGISTERED | [219,220] |
| A Phase II/III Clinical Study of Autologous Cultured Adipocytes (ANTG-adip) for the Treatment of Depressed Scar to Evaluate Safety and Efficacy. | NCT00992147 | COMPLETED | ||
| A Phase I Open-labeled, Single-arm, Single-centred Study to Test the Safety of ADSC-SVF-002 in Subjects With Soft Tissue Defects or Abnormal Wound Healing. | NCT02590042 | RECRUITING | ||
| Facial Atrophy | A Phase II Double-blind, Randomized, Study to Assess the Efficacy of Facial Fat Grafts Supplemented With Autologous, Adipose Derived Stromal Vascular Fraction (SVF). | NCT02526576 | RECRUITING | [210,221,222,223] |
| A Phase One, Open Label, Single Arm Study to Demonstrate the Safety of Antria Cell Preparation Process During Facial Fat Grafting Assisted With Autologous, Adipose Derived Stromal Vascular Fraction (SVF). | NCT01828723 | COMPLETED | ||
| Preliminary Investigation of the Effect of Human Adipose Tissue-derived Mesenchymal Stem Cell (MSC) in Progressive Hemifacial Atrophy (Romberg’s Disease). | NCT01309061 | COMPLETED | ||
| Potential of Mesenchymal Stem Cell Enriched Adipose Tissue Grafting for Contour Deformities of Face. | NCT02494752 | REGISTERED | ||
| Burn and wound healing | A Phase 1 Clinical Study to Evaluate the Safety of Allogeneic Adipose-derived Stem Cells in the Subjects With Deep Second-degree Burn Wound. | NCT02394873 | COMPLETED | [208,224,225,226] |
| Evaluation of Tissue Regeneration Potential (in the Skin) of Child’s Adipose Cells During the Development. | NCT02779205 | COMPLETED | ||
| Feasibility of Obtaining Adipose Derived Regenerative Cells (ADRCs) From Discarded Thermal Burn Eschar Tissue Using Investigational Celution® System for Autologous Treatment of Thermal Burn Injury (The FAST Trial). | NCT02362386 | COMPLETED | ||
| Skin graft | Phase 1/2 Study of Autologous Stromal Vascular Fraction in Adipose Tissue Transplantation in Improving Skin Grafting. | NCT02546882 | RECRUITING | [227,228,229] |
| A Phase I, Two Armed, Open, Prospective and Multicentre Study to Evaluate the Safety of Autologous Tissue-engineered Dermal Substitutes and Dermo-epidermal Skin Substitutes for the Treatment of Large Deep Partial and Full Thickness Skin Defects in Children and Adults. | NCT02145130 | RECRUITING | ||
| Reconstructive surgery | Immunophenotyping of Fresh Stromal Vascular Fraction From Adipose Derived Stem Cells (ADSC) Enriched Fat Grafts for Refinements of Reconstructed Breasts. | NCT01771913 | COMPLETED | [230,231,232,233,234,235,236,237,238,239] |
| Skin improvement in Breast. | NCT01801878 | REGISTERED | ||
| Adipose Stromal Cell Enriched Autologous Fat Grafting for Treating Pain at Amputation Sites: A Single Center Site, Prospective, Randomized, Pilot Outcomes Trial. | NCT02076022 | RECRUITING | ||
| Enriched Autologous Fat Grafting for Treating Pain at Amputation Sites. | NCT01645722 | REGISTERED | ||
| Structural Fat Grafting for Craniofacial Trauma: Effect of Concentrating Endogenous Stromal Cells in the Fat Graft. | NCT01564524 | REGISTERED | ||
| Outcomes After Centrifugation Versus PureGraft for Fatgrafting to the Breast After Breast-conserving Therapy. | NCT01979757 | COMPLETED | ||
| A Clinical Evaluation Of Adipose Derived Regenerative Cells In The Treatment Of Patients With Breast Deformities Post Segmental Breast Resection (Lumpectomy) With Or Without Radiation Therapy. A Phase IV Post Market Study. | NCT00616135 | COMPLETED | ||
| Alopecia | Subcutaneous Transplantation of Autologous Cell Enriched Adipose Tissue For Follicular Niche Stimulation in Early Stage Alopecia Androgenetica (STYLE): a Randomized, Blinded, Controlled Trial. | NCT02503852 | RECRUITING | [137,240] |
| Adipose-derived Stromal Vascular Fraction (SVF) Injections to Stimulate Hair Regrowth for Androgenetic Alopecia. | NCT02626780 | REGISTERED | ||
| Biocellular Regenerative Therapy in Hair Loss: Use of High Density Platelet-Rich Plasma Concentrates and Cell-Enriched Emulsified Adipose-Derived Tissue Stromal Vascular Fraction. | NCT02849470 | RECRUITING | ||
| The Effect of Allogeneic Human Adipose Derived Stem Cell Component Extract on Androgenic Alopecia. | NCT02594046 | REGISTERED | ||
| Point-of-Care Adipose-derived Cells for Hair Growth. | NCT02729415 | REGISTERED | ||
| Adipose Tissue Derived Stem Cell Based Hair Restoration Therapy for Alopecia. | NCT02865421 | REGISTERED | ||
| Urticaria or Scleroderma | Experimental Autologous Mesenchymal Stem Cell Therapy in Treatment of Chronic Autoimmune Urticaria. | NCT02824393 | REGISTERED | [241,242,243] |
| Scleroderma Treatment With Celution Processed Adipose Derived Regenerative Cells (ADRCs) Registry. | NCT02328625 | WITHDRAWN | ||
| Scleroderma Treatment With Celution Processed Adipose Derived Regenerative Cells [134]: A Randomized, Double-Blind, Placebo-Controlled Trial With Incomplete Crossover. | NCT02396238 | REGISTERED | ||
| Subcutaneous Injection of Autologous Adipose Tissue-derived SVF Into the Fingers of Patients With Systemic Sclerosis: Controlled Clinical Trial With Efficacy Assessment. | NCT02558543 | RECRUITING | ||
| Pressure Ulcer | Treatment of Hypertensive Leg Ulcer by Adipose Tissue Grafting. | NCT01932021 | REGISTERED | [133,244,245,246] |
| A Pilot Study: Evaluating the Safety/Feasibility of ADSC on Adults With Stage III or IV Pressure Ulcers. | NCT02375802 | RECRUITING | ||
| Adipose Derived Regenerative Cellular Therapy of Chronic Wounds. | NCT02092870 | RECRUITING | ||
| Lipodystrophies | Phase I Study of a Filler Agent Composed of Mesenchymal Stem Cells Obtained From Autologous Adipose Tissue Associated With Hyaluronic Acid. | NCT02034786 | REGISTERED | [247] |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaur, M.; Dobke, M.; Lunyak, V.V. Mesenchymal Stem Cells from Adipose Tissue in Clinical Applications for Dermatological Indications and Skin Aging. Int. J. Mol. Sci. 2017, 18, 208. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010208
Gaur M, Dobke M, Lunyak VV. Mesenchymal Stem Cells from Adipose Tissue in Clinical Applications for Dermatological Indications and Skin Aging. International Journal of Molecular Sciences. 2017; 18(1):208. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010208
Chicago/Turabian StyleGaur, Meenakshi, Marek Dobke, and Victoria V. Lunyak. 2017. "Mesenchymal Stem Cells from Adipose Tissue in Clinical Applications for Dermatological Indications and Skin Aging" International Journal of Molecular Sciences 18, no. 1: 208. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010208