
Evolutionary Conservation of pou5f3 Genomic Organization and Its Dynamic Distribution during Embryogenesis and in Adult Gonads in Japanese Flounder Paralichthys olivaceus

Abstract
:1. Introduction
2. Results
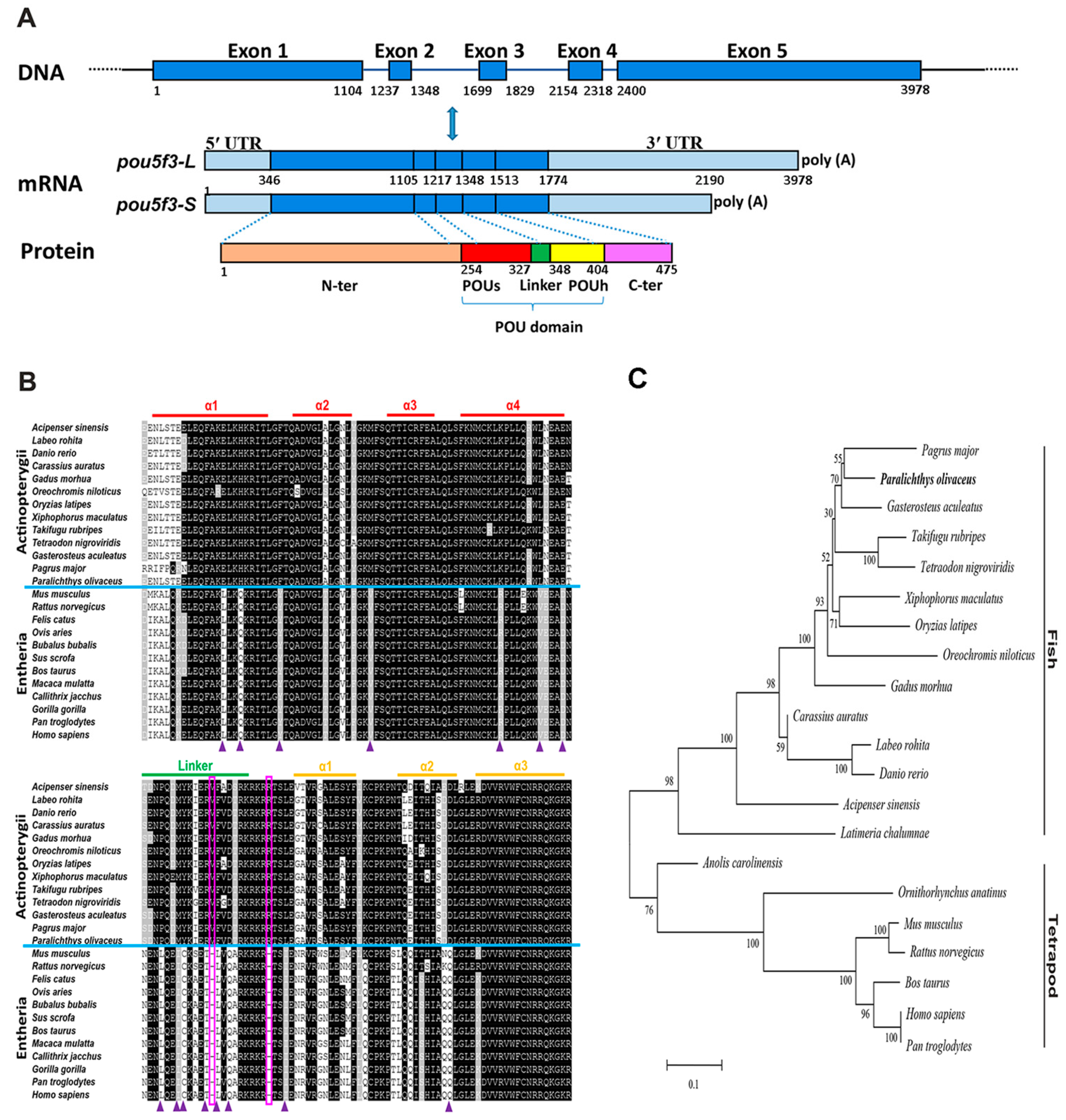
2.1. Molecular Characterization and Homological Analysis of P. olivaceus pou5f3
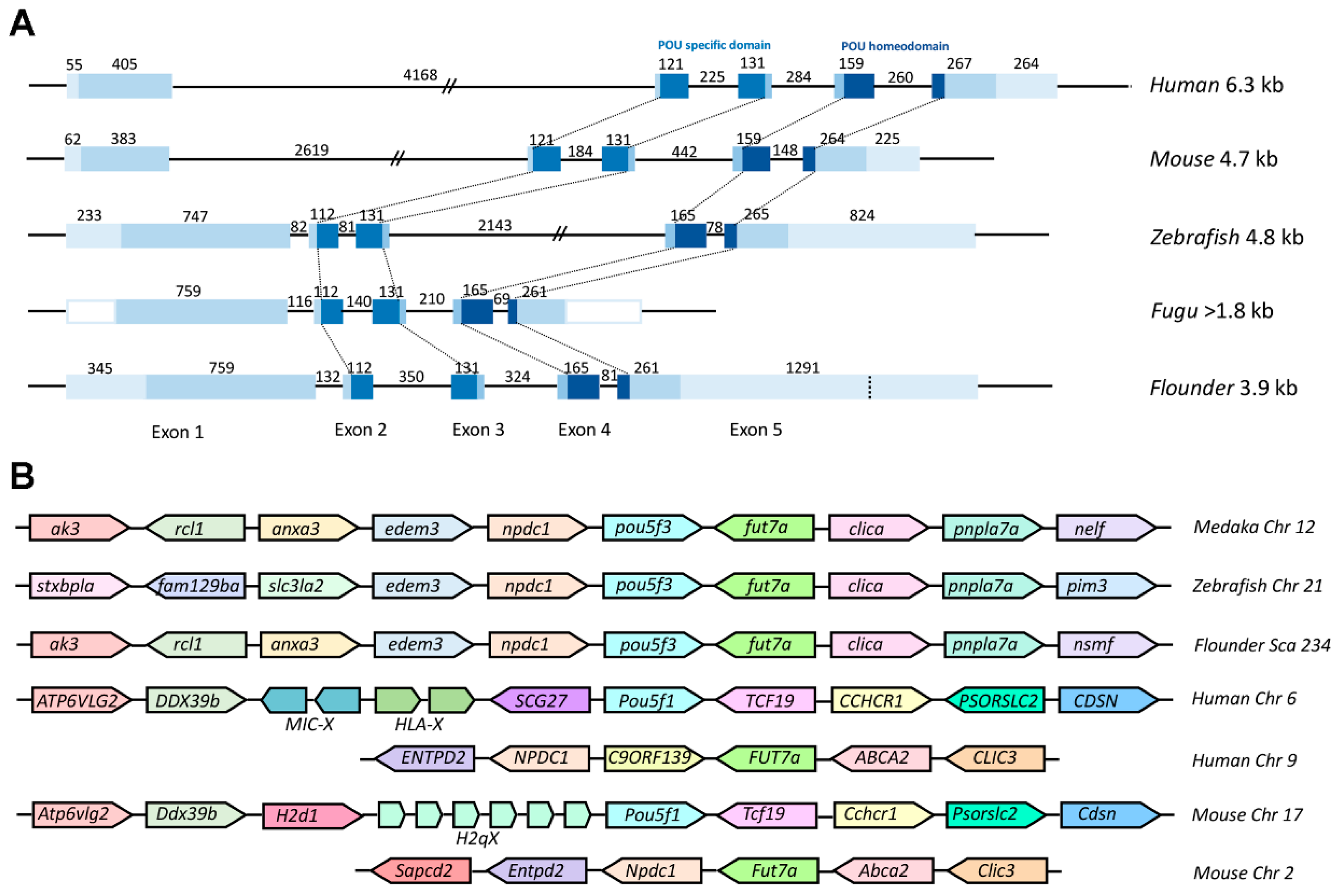
2.2. Genomic Organization and Conserved Synteny of Class V POU Family Members
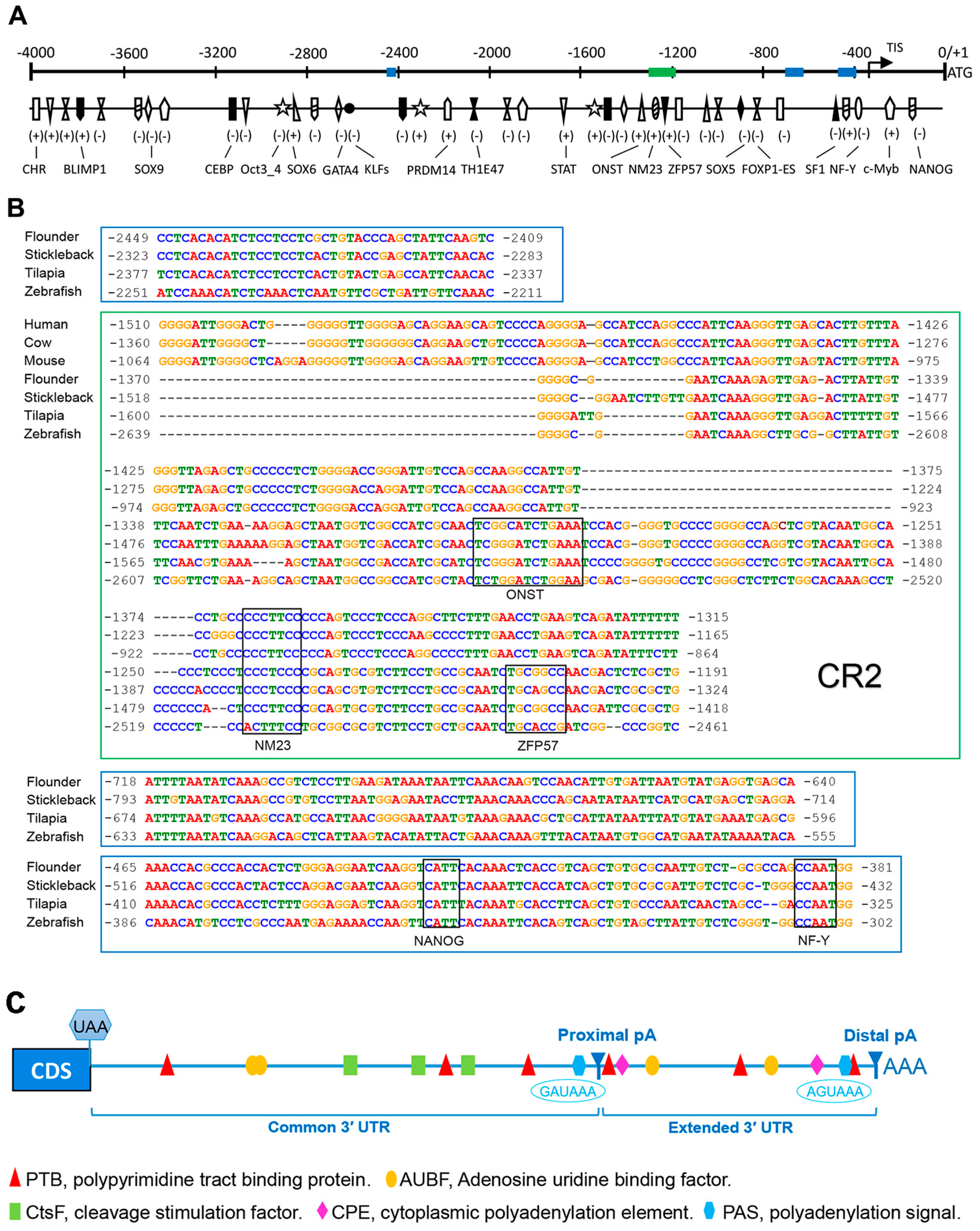
2.3. Bioinformatics Analysis of Popou5f3 5′ and 3′ Regulatory Sequences
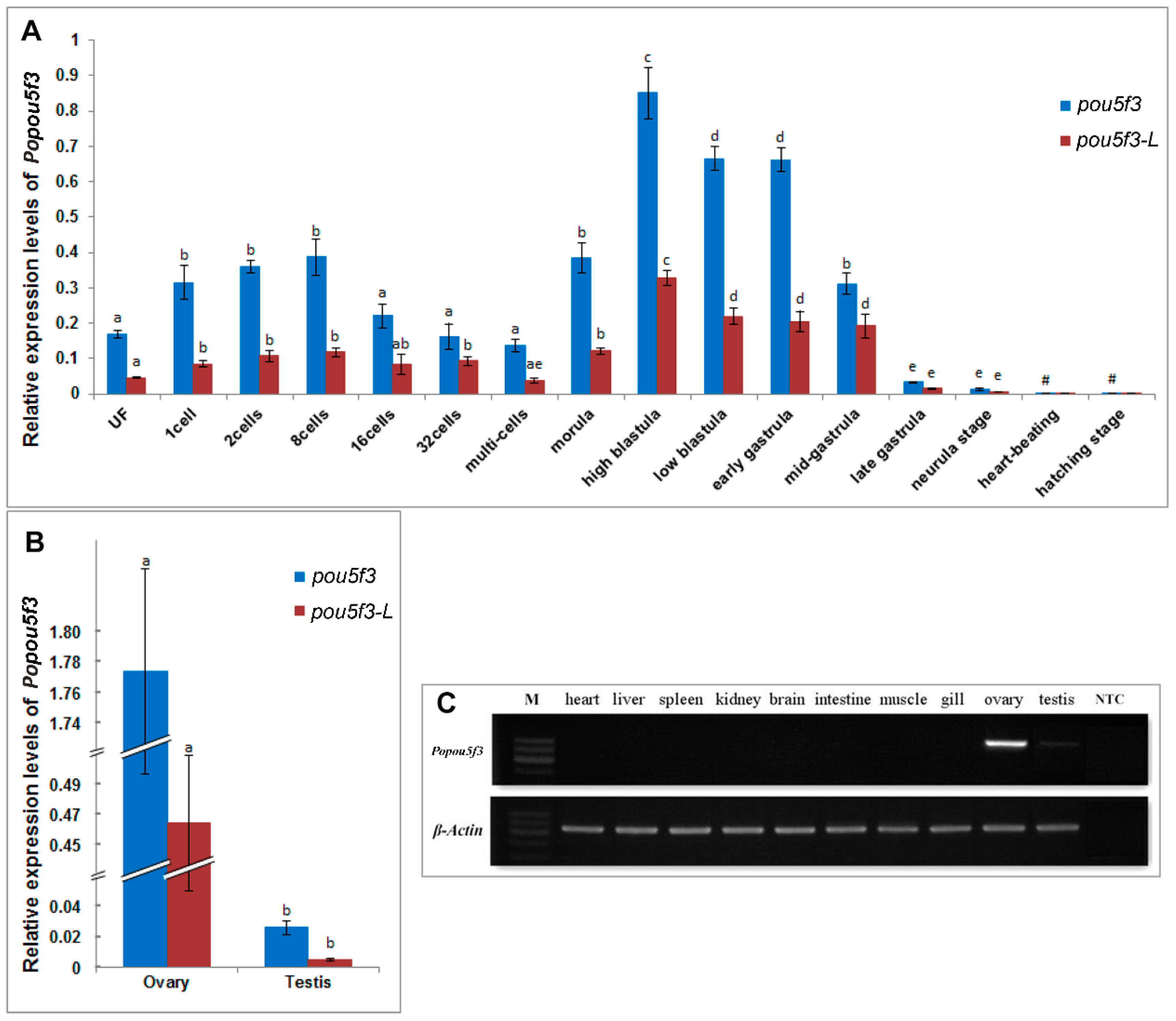
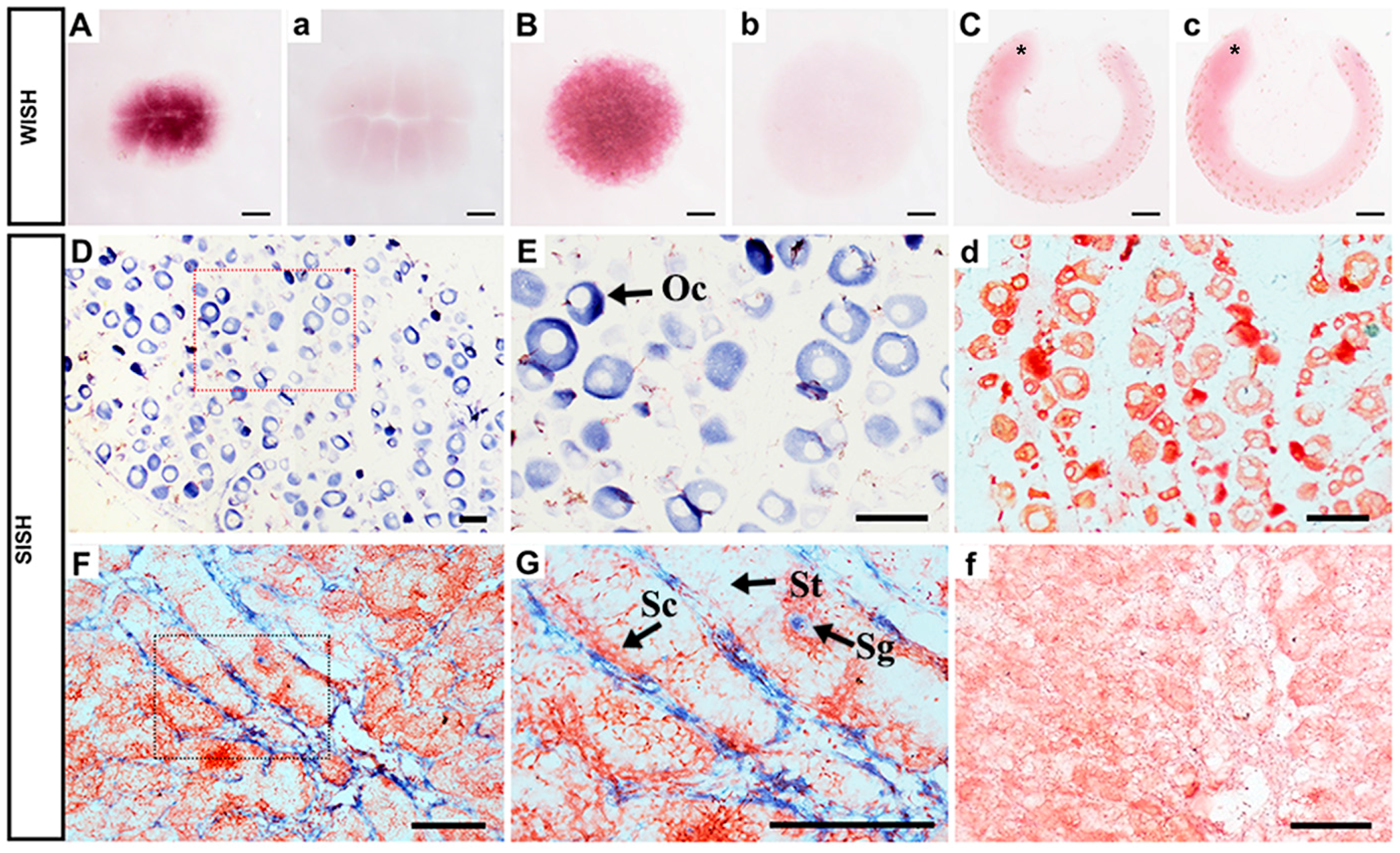
2.4. Spatiotemporal Expression of Popou5f3 during Early Embryogensis
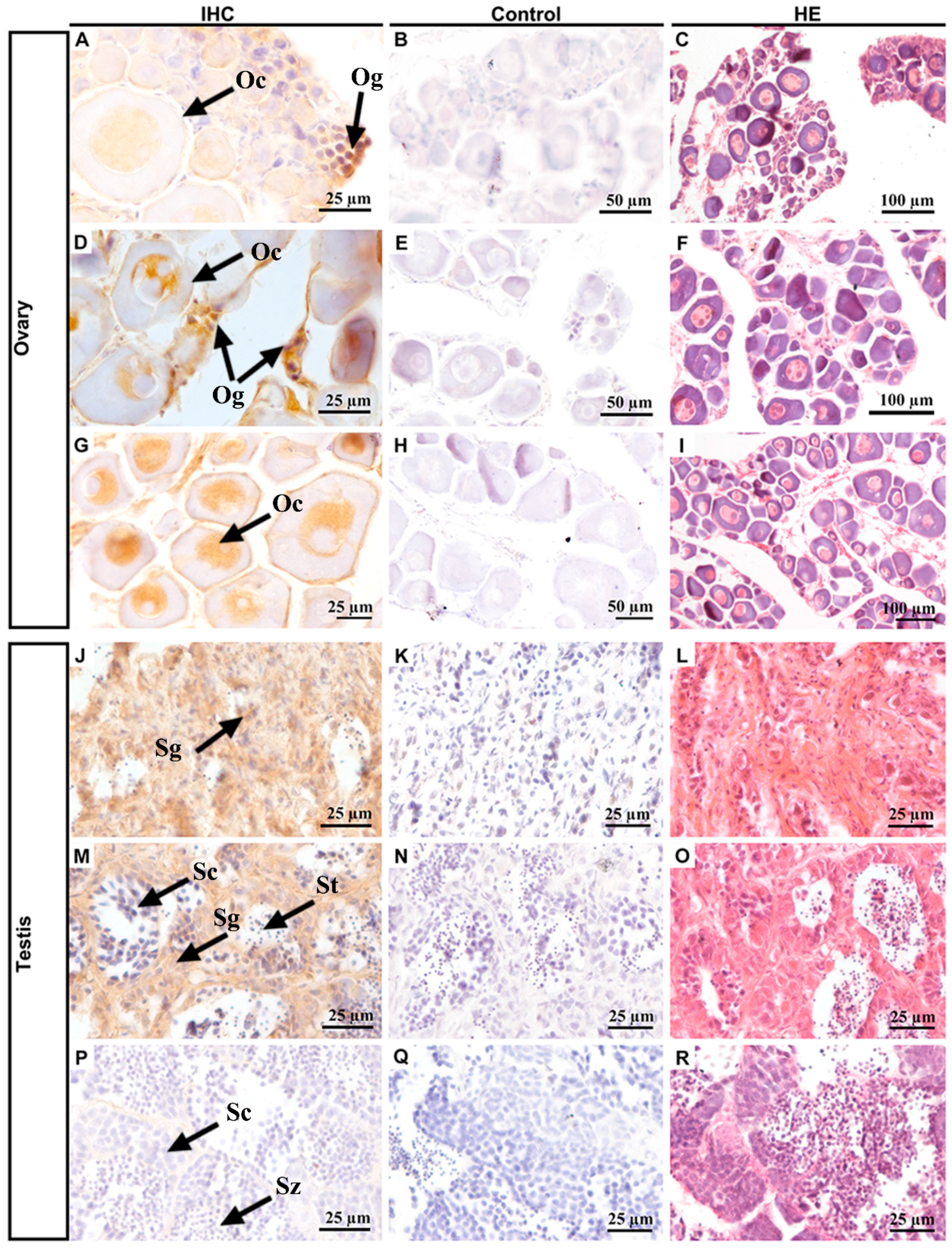
2.5. Gonad-Specific Expression of Popou5f3 in Adult Tissues
3. Discussion
3.1. Popou5f3 Is the Homologue of Mammalian POU5F1
3.2. The Potential Cis-Regulatory Elements (CREs) in Regulating Popou5f3 Expression
3.3. Popou5f3 Is Abundant at Blastula-Stage Embryos and Restricted in Adult Gonads
4. Experimental Section
4.1. Animals and Handling
4.2. Genomic DNA, Total RNA Isolation and cDNA Synthesis
4.3. Cloning and Sequencing of Popou5f3
4.4. Sequence Analysis
4.5. Quantitative Expression Analysis of Popou5f3
4.6. In Situ Hybridization Analysis
4.7. Immunohistochemistry and Histological Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Herr, W.; Cleary, M.A. The POU domain: Versatility in transcriptional regulation by a flexible two-in-one DNA-binding domain. Genes Dev. 1995, 9, 1679–1693. [Google Scholar] [CrossRef] [PubMed]
- Jerabek, S.; Merino, F.; Scholer, H.R.; Cojocaru, V. OCT4: Dynamic DNA binding pioneers stem cell pluripotency. Biochim. Biophys. Acta 2014, 1839, 138–154. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Corbi, N.; Basilico, C.; Dailey, L. Developmental-specific activity of the FGF-4 enhancer requires the synergistic action of Sox2 and Oct3. Genes Dev. 1995, 9, 2635–2645. [Google Scholar] [CrossRef] [PubMed]
- Chew, J.L.; Loh, Y.H.; Zhang, W.; Chen, X.; Tam, W.L.; Yeap, L.S.; Li, P.; Ang, Y.S.; Lim, B.; Robson, P.; et al. Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the Oct4/Sox2 complex in embryonic stem cells. Mol. Cell. Biol. 2005, 25, 6031–6046. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Tomlinson, S.R. The transcriptional foundation of pluripotency. Development 2009, 136, 2311–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, H. How is pluripotency determined and maintained? Development 2007, 134, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Loh, Y.-H.; Wu, Q.; Chew, J.-L.; Vega, V.B.; Zhang, W.; Chen, X.; Bourque, G.; George, J.; Leong, B.; Liu, J. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat. Genet. 2006, 38, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, S.L.; Peter, W.; Hess, H.; Scholer, H.R. Oct4 transcription factor is differentially expressed in the mouse embryo during establishment of the first two extraembryonic cell lineages involved in implantation. Dev. Biol. 1994, 166, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Kehler, J.; Tolkunova, E.; Koschorz, B.; Pesce, M.; Gentile, L.; Boiani, M.; Lomeli, H.; Nagy, A.; McLaughlin, K.J.; Scholer, H.R.; et al. Oct4 is required for primordial germ cell survival. EMBO Rep. 2004, 5, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Boiani, M.; Eckardt, S.; Scholer, H.R.; McLaughlin, K.J. Oct4 distribution and level in mouse clones: Consequences for pluripotency. Genes Dev. 2002, 16, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Scholer, H.; Smith, A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 1998, 95, 379–391. [Google Scholar] [CrossRef]
- Rosner, M.H.; Vigano, M.A.; Ozato, K.; Timmons, P.M.; Poirier, F.; Rigby, P.W.; Staudt, L.M. A POU-domain transcription factor in early stem cells and germ cells of the mammalian embryo. Nature 1990, 345, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Scholer, H.R.; Dressler, G.R.; Balling, R.; Rohdewohld, H.; Gruss, P. Oct4: A germline-specific transcription factor mapping to the mouse t-complex. EMBO J. 1990, 9, 2185–2195. [Google Scholar] [PubMed]
- Yeom, Y.I.; Ha, H.S.; Balling, R.; Scholer, H.R.; Artzt, K. Structure, expression and chromosomal location of the Oct4 gene. Mech. Dev. 1991, 35, 171–179. [Google Scholar] [PubMed]
- Belting, H.G.; Hauptmann, G.; Meyer, D.; Abdelilah-Seyfried, S.; Chitnis, A.; Eschbach, C.; Soll, I.; Thisse, C.; Thisse, B.; Artinger, K.B.; et al. Spiel ohne grenzen/pou2 is required during establishment of the zebrafish midbrain-hindbrain boundary organizer. Development 2001, 128, 4165–4176. [Google Scholar] [PubMed]
- Sanchez-Sanchez, A.V.; Camp, E.; Garcia-Espana, A.; Leal-Tassias, A.; Mullor, J.L. Medaka Oct4 is expressed during early embryo development, and in primordial germ cells and adult gonads. Dev. Dyn. 2010, 239, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Marandel, L.; Labbe, C.; Bobe, J.; Jammes, H.; Lareyre, J.J.; Le Bail, P.Y. Do not put all teleosts in one net: Focus on the sox2 and pou2 genes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2013, 164, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, Z.; Linyan, L.; Zhenhua, F.; Linyan, Z.; Zhijian, W.; Ling, W.; Deshou, W.; Jing, W. Characterization of the POU5F1 homologue in nile tilapia: From expression pattern to biological activity. Stem Cells Dev. 2016, 25, 1386–1395. [Google Scholar]
- Lunde, K.; Belting, H.-G.; Driever, W. Zebrafish pou5f1/pou2, homolog of mammalian Oct4, functions in the endoderm specification cascade. Curr. Biol. 2004, 14, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Matsuzaki, T.; Oki, T.; Miyagawa, T.; Amanuma, H. A novel POU domain gene, zebrafish pou2: Expression and roles of two alternatively spliced twin products in early development. Genes Dev. 1994, 8, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Manali, D.; Wang, T.; Bhat, N.; Hong, N.; Li, Z.; Wang, L.; Yan, Y.; Liu, R.; Hong, Y. Identification of pluripotency genes in the fish medaka. Int. J. Biol. Sci. 2011, 7, 440. [Google Scholar] [CrossRef] [PubMed]
- Tapia, N.; Reinhardt, P.; Duemmler, A.; Wu, G.; Arauzo-Bravo, M.J.; Esch, D.; Greber, B.; Cojocaru, V.; Rascon, C.A.; Tazaki, A.; et al. Reprogramming to pluripotency is an ancient trait of vertebrate Oct4 and Pou2 proteins. Nat. Commun. 2012, 3, 1279. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Gui, J.; Chen, S.; Deng, J.; Manfred, S. Embryonic stem cells in fish. Acta Zoolog. Sin. 2002, 49, 281–294. [Google Scholar]
- Hong, Y.; Schartl, M. Establishment and growth responses of early medakafish (Oryzias latipes) embryonic cells in feeder layer-free cultures. Mol. Mar. Biol. Biotechnol. 1996, 5, 93–104. [Google Scholar]
- Béjar, J.; Hong, Y.; Alvarez, M.C. An ES-like cell line from the marine fish Sparus aurata: Characterization and chimaera production. Transgenic Res. 2002, 11, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-L.; Sha, Z.-X.; Ye, H.-Q. Establishment of a pluripotent embryonic cell line from sea perch (Lateolabrax japonicus) embryos. Aquaculture 2003, 218, 141–151. [Google Scholar] [CrossRef]
- Parameswaran, V.; Shukla, R.; Bhonde, R.; Hameed, A.S. Development of a pluripotent ES-like cell line from asian sea bass (Lates calcarifer)—An oviparous stem cell line mimicking viviparous ES cells. Mar. Biotechnol. 2007, 9, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Dash, C.; Routray, P.; Tripathy, S.; Verma, D.K.; Guru, B.C.; Meher, P.K.; Nandi, S.; Eknath, A.E. Derivation and characterization of embryonic stem-like cells of Indian major carp Catla catla. J. Fish Biol. 2010, 77, 1096–1113. [Google Scholar] [CrossRef] [PubMed]
- Holen, E.; Kausland, A.; Skjaerven, K. Embryonic stem cells isolated from atlantic cod (Gadus morhua) and the developmental expression of a stage-specific transcription factor ac-pou2. Fish Physiol. Biochem. 2010, 36, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, J.; Jiang, J.; Fan, L.; Wang, W.; Liu, J.; Zhang, Q.; Wang, X. Identification and characterization of a nanog homolog in Japanese flounder (Paralichthys olivaceus). Gene 2013, 531, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Okazawa, H.; Okamoto, K.; Ishino, F.; Ishino-Kaneko, T.; Takeda, S.; Toyoda, Y.; Muramatsu, M.; Hamada, H. The Oct3 gene, a gene for an embryonic transcription factor, is controlled by a retinoic acid repressible enhancer. EMBO J. 1991, 10, 2997–3005. [Google Scholar] [PubMed]
- Howley, C.; Ho, R.K. mRNA localization patterns in zebrafish oocytes. Mech. Dev. 2000, 92, 305–309. [Google Scholar] [CrossRef]
- Takeda, J.; Seino, S.; Bell, G.I. Human Oct3 gene family: cDNA sequences, alternative splicing, gene organization, chromosomal location, and expression at low levels in adult tissues. Nucleic Acids Res. 1992, 20, 4613–4620. [Google Scholar] [CrossRef] [PubMed]
- Van Eijk, M.; Van Rooijen, M.; Modina, S.; Scesi, L.; Folkers, G.; Van Tol, H.; Bevers, M.; Fisher, S.; Lewin, H.; Rakacolli, D. Molecular cloning, genetic mapping, and developmental expression of bovine POU5F1. Biol. Reprod. 1999, 60, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, S.; Tisdall, D.; Selwood, L. Identification of a homologue of POU5F1 (OCT3/4) in a marsupial, the brushtail possum. Mol. Reprod. Dev. 2001, 58, 255–261. [Google Scholar] [CrossRef]
- Frankenberg, S.; Pask, A.; Renfree, M.B. The evolution of class V POU domain transcription factors in vertebrates and their characterisation in a marsupial. Dev. Biol. 2010, 337, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Mitalipov, S.M.; Kuo, H.C.; Hennebold, J.D.; Wolf, D.P. Oct4 expression in pluripotent cells of the rhesus monkey. Biol. Reprod. 2003, 69, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Niwa, H.; Sekita, Y.; Tsend-Ayush, E.; Grützner, F. Platypus Pou5f1 reveals the first steps in the evolution of trophectoderm differentiation and pluripotency in mammals. Evol. Dev. 2008, 10, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, S.P.; Shevchenko, A.I.; Elisaphenko, E.A.; Nesterova, T.B.; Brockdorff, N.; Zakian, S.M. Structure and expression pattern of Oct4 gene are conserved in vole Microtus rossiaemeridionalis. BMC Genom. 2008, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Lavial, F.; Acloque, H.; Bertocchini, F.; Macleod, D.J.; Boast, S.; Bachelard, E.; Montillet, G.; Thenot, S.; Sang, H.M.; Stern, C.D.; et al. The Oct4 homologue PouV and nanog regulate pluripotency in chicken embryonic stem cells. Development 2007, 134, 3549–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, G.M.; Brickman, J.M. Conserved roles for oct4 homologues in maintaining multipotency during early vertebrate development. Development 2006, 133, 2011–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Nakamoto, A.; Tai, M.; Saito, S.; Nakayama, Y.; Kawamura, A.; Takeda, H.; Yamasu, K. Mesendoderm specification depends on the function of Pou2, the class V POU-type transcription factor, during zebrafish embryogenesis. Dev. Growth Differ. 2012, 54, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, M.; Li, Z.; Hong, N.; Xu, H.; Hong, Y. Medaka Oct4 is essential for pluripotency in blastula formation and ES cell derivation. Stem Cell Rev. 2015, 11, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Du, H.; Chen, X.H.; Cao, H.; Liu, T.; Li, C.J. Identification of a pou2 ortholog in chinese sturgeon, Acipenser sinensis and its expression patterns in tissues, immature individuals and during embryogenesis. Fish Physiol. Biochem. 2012, 38, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Reim, G.; Chen, W.; Hopkins, N.; Brand, M. The zebrafish spiel-ohne-grenzen (spg) gene encodes the POU domain protein Pou2 related to mammalian Oct4 and is essential for formation of the midbrain and hindbrain, and for pre-gastrula morphogenesis. Development 2002, 129, 905–916. [Google Scholar] [PubMed]
- Wagner, J.T.; Podrabsky, J.E. Gene expression patterns that support novel developmental stress buffering in embryos of the annual killifish Austrofundulus limnaeus. EvoDevo 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, S.; Renfree, M.B. On the origin of POU5F1. BMC Biol. 2013, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, S.R.; Frank, D.; Harland, R.; Johnson, A.D.; Nichols, J.; Niwa, H.; Scholer, H.R.; Tanaka, E.; Wylie, C.; Brickman, J.M. The POU-er of gene nomenclature. Development 2014, 141, 2921–2923. [Google Scholar] [CrossRef] [PubMed]
- Onichtchouk, D. Evolution and functions of Oct4 homologs in non-mammalian vertebrates. Biochim. Biophys. Acta 2016, 1859, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, S.; Enczmann, J.; Waclawczyk, S.; Wernet, P.; Kogler, G. Oct4 and its pseudogenes confuse stem cell research. Cell Stem Cell 2007, 1, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Parvin, M.S.; Okuyama, N.; Inoue, F.; Islam, M.E.; Kawakami, A.; Takeda, H.; Yamasu, K. Autoregulatory loop and retinoic acid repression regulate pou2/pou5f1 gene expression in the zebrafish embryonic brain. Dev. Dyn. 2008, 237, 1373–1388. [Google Scholar] [CrossRef] [PubMed]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of nanog by Oct4 and Sox2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [PubMed]
- McGregor, A.P.; Orgogozo, V.; Delon, I.; Zanet, J.; Srinivasan, D.G.; Payre, F.; Stern, D.L. Morphological evolution through multiple cis-regulatory mutations at a single gene. Nature 2007, 448, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.L.; Frankel, N. The structure and evolution of cis-regulatory regions: The shavenbaby story. Philos. Trans. R. Soc. Lond. 2013, 368, 20130028. [Google Scholar] [CrossRef] [PubMed]
- Nordhoff, V.; Hubner, K.; Bauer, A.; Orlova, I.; Malapetsa, A.; Scholer, H.R. Comparative analysis of human, bovine, and murine Oct4 upstream promoter sequences. Mamm. Genome 2001, 12, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Do, H.J.; Oh, J.H.; Kim, J.H.; Choi, S.Y.; Cha, K.Y.; Chung, H.M.; Kim, J.H. Characterization of putative cis-regulatory elements that control the transcriptional activity of the human Oct4 promoter. J. Cell. Biochem. 2005, 96, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Minucci, S.; Botquin, V.; Yeom, Y.I.; Dey, A.; Sylvester, I.; Zand, D.J.; Ohbo, K.; Ozato, K.; Scholer, H.R. Retinoic acid-mediated down-regulation of Oct3/4 coincides with the loss of promoter occupancy in vivo. EMBO J. 1996, 15, 888–899. [Google Scholar] [PubMed]
- Gu, P.; LeMenuet, D.; Chung, A.C.; Mancini, M.; Wheeler, D.A.; Cooney, A.J. Orphan nuclear receptor GCNF is required for the repression of pluripotency genes during retinoic acid-induced embryonic stem cell differentiation. Mol. Cell. Biol. 2005, 25, 8507–8519. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Usuda, M.; Matsuda, T.; Ko, M.S.; Niwa, H.; Asano, M.; Koide, H.; Yokota, T. Identification of Zfp-57 as a downstream molecule of STAT3 and Oct3/4 in embryonic stem cells. Biochem. Biophys. Res. Commun. 2005, 331, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Gabut, M.; Samavarchi-Tehrani, P.; Wang, X.; Slobodeniuc, V.; O’Hanlon, D.; Sung, H.K.; Alvarez, M.; Talukder, S.; Pan, Q.; Mazzoni, E.O.; et al. An alternative splicing switch regulates embryonic stem cell pluripotency and reprogramming. Cell 2011, 147, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.G.; Chung, I.Y.; Lim, Y.; Lee, Y.H.; Shin, S.Y. A Tcf/Lef element within the enhancer region of the human NANOG gene plays a role in promoter activation. Biochem. Biophys. Res. Commun. 2011, 410, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Tsuneyoshi, N.; Sumi, T.; Onda, H.; Nojima, H.; Nakatsuji, N.; Suemori, H. PRDM14 suppresses expression of differentiation marker genes in human embryonic stem cells. Biochem. Biophys. Res. Commun. 2008, 367, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.H.; Sampsell-Barron, T.L.; Gu, F.; Root, S.; Peck, R.M.; Pan, G.; Yu, J.; Antosiewicz-Bourget, J.; Tian, S.; Stewart, R.; et al. NANOG is a direct target of TGFβ/activin-mediated SMAD signaling in human ESCs. Cell Stem Cell 2008, 3, 196–206. [Google Scholar] [CrossRef] [PubMed]
- He, H.; McHaney, M.; Hong, J.; Weiss, M.L. Cloning and characterization of 3.1 kb promoter region of the Oct4 gene from the Fischer 344 rat. Open Stem Cell J. 2009, 1, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Kobolak, J.; Kiss, K.; Polgar, Z.; Mamo, S.; Rogel-Gaillard, C.; Tancos, Z.; Bock, I.; Baji, A.G.; Tar, K.; Pirity, M.K.; et al. Promoter analysis of the rabbit POU5F1 gene and its expression in preimplantation stage embryos. BMC Mol. Biol. 2009, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Winkler, C.; Liu, T.; Chai, G.; Schartl, M. Activation of the mouse Oct4 promoter in medaka embryonic stem cells and its use for ablation of spontaneous differentiation. Mech. Dev. 2004, 121, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dai, J. Concise review: Isoforms of Oct4 contribute to the confusing diversity in stem cell biology. Stem Cells 2010, 28, 885–893. [Google Scholar] [PubMed]
- Hwang, J.Y.; Oh, J.N.; Lee, D.K.; Choi, K.H.; Park, C.H.; Lee, C.K. Identification and differential expression patterns of porcine Oct4 variants. Reproduction 2015, 149, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Nimura, K.; Yamamoto, M.; Takeichi, M.; Saga, K.; Takaoka, K.; Kawamura, N.; Nitta, H.; Nagano, H.; Ishino, S.; Tanaka, T.; et al. Regulation of alternative polyadenylation by Nkx2–5 and Xrn2 during mouse heart development. eLife 2016, 5, e16030. [Google Scholar] [CrossRef] [PubMed]
- Miles, W.O.; Lembo, A.; Volorio, A.; Brachtel, E.; Tian, B.; Sgroi, D.; Provero, P.; Dyson, N. Alternative polyadenylation in triple-negative breast tumors allows NRAS and c-JUN to bypass PUMILIO post-transcriptional regulation. Cancer Res. 2016, 76. [Google Scholar] [CrossRef] [PubMed]
- Boutet, S.C.; Cheung, T.H.; Quach, N.L.; Liu, L.; Prescott, S.L.; Edalati, A.; Iori, K.; Rando, T.A. Alternative polyadenylation mediates microRNA regulation of muscle stem cell function. Cell Stem Cell 2012, 10, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Lackford, B.; Yao, C.; Charles, G.M.; Weng, L.; Zheng, X.; Choi, E.A.; Xie, X.; Wan, J.; Xing, Y.; Freudenberg, J.M.; et al. Fip1 regulates mRNA alternative polyadenylation to promote stem cell self-renewal. EMBO J. 2014, 33, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Jin, Y. Role of Oct4 in maintaining and regaining stem cell pluripotency. Stem Cell Res. Ther. 2010, 1, 39. [Google Scholar] [CrossRef] [PubMed]
- Reim, G.; Brand, M. Spiel-ohne-grenzen/pou2 mediates regional competence to respond to Fgf8 during zebrafish early neural development. Development 2002, 129, 917–933. [Google Scholar] [PubMed]
- Gao, J.; Wang, Z.; Shao, K.; Fan, L.; Yang, L.; Song, H.; Liu, M.; Wang, Z.; Wang, X.; Zhang, Q. Identification and characterization of a Sox2 homolog in the Japanese flounder Paralichthys olivaceus. Gene 2014, 544, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.; Liu, S.J.; Zou, L.N.; Smith, Z.; Meissner, A.; Ramanathan, S. Pluripotency factors in embryonic stem cells regulate differentiation into germ layers. Cell 2011, 145, 875–889. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Chen, S.; Yan, A.; Ji, X. Study on the embryonic development of Paralichthys olivaceus. J. Fish. China 2004, 28, 609–615. [Google Scholar]
- Gao, J.; Li, P.; Zhang, W.; Wang, Z.; Wang, X.; Zhang, Q. Molecular cloning, promoter analysis and expression profiles of the Sox3 gene in Japanese flounder, Paralichthys olivaceus. Int. J. Mol. Sci. 2015, 16, 27931–27944. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PoPou5f3 Vs * | Full-Length (%) | N-Terminal (%) | POU Domain (%) | POU Domain | C-Terminal (%) | ||
|---|---|---|---|---|---|---|---|
| POUs (%) | Linker (%) | POUh (%) | |||||
| H.s POU5F1 | 37.9 | 23.4 | 68.5 | 75.7 | 36.8 | 69.6 | 14.3 |
| B.t Pou5f1 | 39.3 | 20.4 | 68.5 | 74.3 | 36.8 | 71.4 | 15.9 |
| M.m Pou5f1 | 39.4 | 20.0 | 68.5 | 73.0 | 36.8 | 73.2 | 14.5 |
| T.g Pou5f3 | 49.5 | 10.8 | 74.2 | 87.8 | 40.0 | 68.4 | 32.3 |
| A.c Pou5f3 | 37.2 | 15.7 | 69.6 | 81.1 | 22.2 | 69.6 | 14.5 |
| A.m Pou5f3 | 43.4 | 19.1 | 73.5 | 89.2 | 25.0 | 70.2 | 41.2 |
| T.r Pou5f3 | 81.5 | 75.1 | 94.7 | 94.6 | 85.0 | 98.2 | 76.1 |
| O.l Pou5f3 | 80.8 | 72.5 | 97.4 | 98.6 | 90.0 | 98.2 | 74.3 |
| D.r Pou5f3 | 70.4 | 58.4 | 94.7 | 95.9 | 90.0 | 94.7 | 60.0 |
| Experiment | Primer Name | Primer Sequences (5′–3′) |
|---|---|---|
| Core fragment | pou5f3-core-Fw | TGTTCAGYCAGACVACMATY |
| pou5f3-core-Rv-1 | CDCGBACYACRTCTCTCTC | |
| pou5f3-core-Rv-2 | AGCAGYGGGTCVTCCAG | |
| RACE PCR | 5′-RACE-pou5f3 | GTAATCTCCTGGGTGTTGGGTTTG |
| 3′-RACE-pou5f3 | GCAGAGACCTCAGACAATCCCCAG | |
| Full-length cDNA | pou5f3-full-length-Fw | CTATCTCGGACTTGTTCTTG |
| pou5f3-full-length-Rv | CATAACTTGCCTTTGCTG | |
| Genome walking(1st) | pou5f3-Pro-SP1-1st | CTTCCAAACGCTGTGGGAGATT |
| pou5f3-Pro-SP2-1st | CAAAGAGGAACGAGCGGAGGAC | |
| pou5f3-Pro-SP3-1st | TGCCACATTACCACCAACACGC | |
| Genome walking | pou5f3-Pro-SP1-2nd | TATCGTGCCCTTATGGTGTCCTT |
| pou5f3-Pro-SP2-2nd | TAAAGCACCCATGTAGGTCAA | |
| pou5f3-Pro-SP3-2nd | CTCAGCCATGCAAGCATTGTCC | |
| In Situ hybridization | pou5f3-ISH-Fw | ACAGGTCGACTGAAACGGATGAGCACG |
| pou5f3-ISH-Rv | AGCGGAATTCGTGATGCCACCATACGC | |
| Prokaryotic expression | pou5f3-PE-Fw | CGAATTCATGTCTGAAAGATCTCAGAGTC |
| pou5f3-PE-Rv | TGCGGCCGCTTATCCAGTCATGTGACCAAT | |
| SemiRT-PCR | pou5f3-SRT-Fw | AACACTGGTATCCCTTCGCT |
| pou5f3-SRT-Rv | CATCTTACCATAGAGGTTACCC | |
| Reference gene | β-actin-Fw | GAGATGAAGCCCAGAGCAAGAG |
| β-actin-Rv | CAGCTGTGGTGGTGAAGGAGTAG | |
| qRT-PCR | pou5f3-RT-Fw | GAGGCGTGTTGGTGGTAATGTG |
| pou5f3-RT-Rv | AAGAGGAACGAGCGGAGGAC | |
| qRT-PCR | pou5f3-L-RT-Fw | TGACTGTAACCACCTGAGACCTAA |
| pou5f3-L-RT-Rv | AAGATTTACTGGGAAAAGGAACAA | |
| qRT-PCR | 18S-RT-Fw | GGTAACGGGGAATCAGGGT |
| 18S-RT-Rv | TGCCTTCCTTGGATGTGGT | |
| qRT-PCR | UbcE-RT-Fw | TTACTGTCCATTTCCCCACTGAC |
| UbcE-RT-Rv | GACCACTGCGACCTCAAGATG |
| Website Name | Link |
|---|---|
| NCBI′s online ORF Finder | http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gorf/gorf.html |
| SMART | http://smart.embl-heidelberg.de/ |
| InterProScan | http://www.ebi.ac.uk/Tools/pfa/iprscan/ |
| MatInspector | http://www.genomatix.de/matinspector.html |
| MEME | http://meme.nbcr.net/meme/cgi-bin/meme.cgi |
| Genomarix suite | http://www.genomatix.de/cgi-bin/dialign/dialign.pl |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Wang, X.; Zhang, Q. Evolutionary Conservation of pou5f3 Genomic Organization and Its Dynamic Distribution during Embryogenesis and in Adult Gonads in Japanese Flounder Paralichthys olivaceus. Int. J. Mol. Sci. 2017, 18, 231. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010231
Gao J, Wang X, Zhang Q. Evolutionary Conservation of pou5f3 Genomic Organization and Its Dynamic Distribution during Embryogenesis and in Adult Gonads in Japanese Flounder Paralichthys olivaceus. International Journal of Molecular Sciences. 2017; 18(1):231. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010231
Chicago/Turabian StyleGao, Jinning, Xubo Wang, and Quanqi Zhang. 2017. "Evolutionary Conservation of pou5f3 Genomic Organization and Its Dynamic Distribution during Embryogenesis and in Adult Gonads in Japanese Flounder Paralichthys olivaceus" International Journal of Molecular Sciences 18, no. 1: 231. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18010231