Hydrogen Gas Is Involved in Auxin-Induced Lateral Root Formation by Modulating Nitric Oxide Synthesis


Abstract
:
1. Introduction
2. Results
2.1. Exogenous Hydrogen Gas (H2)-Induced Lateral Root Formation in A Dose-Dependent Manner
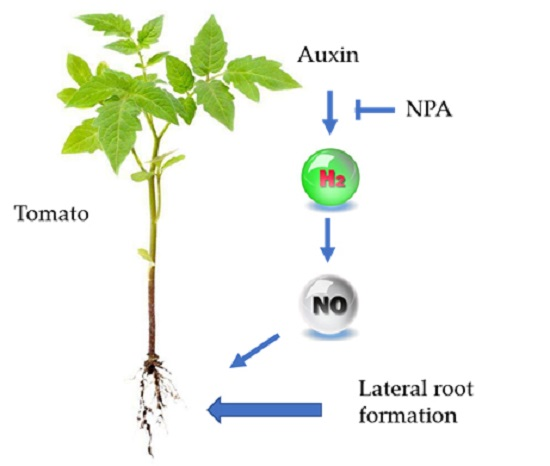
2.2. A Possible Link between Auxin and H2 in the Induction of Lateral Root Formation
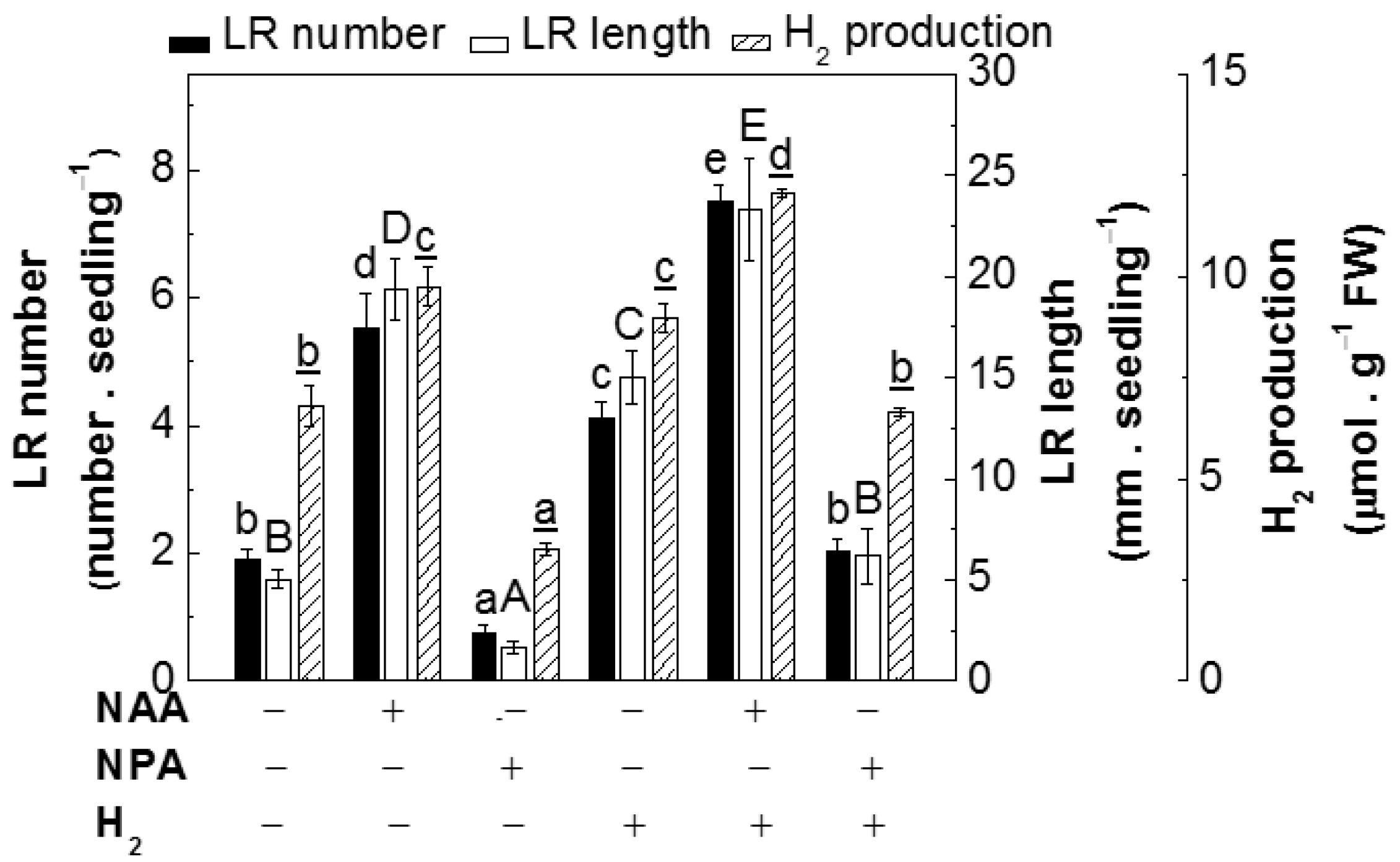
2.3. H2 Was Partly Involved in Auxin-Induced Lateral Root Formation
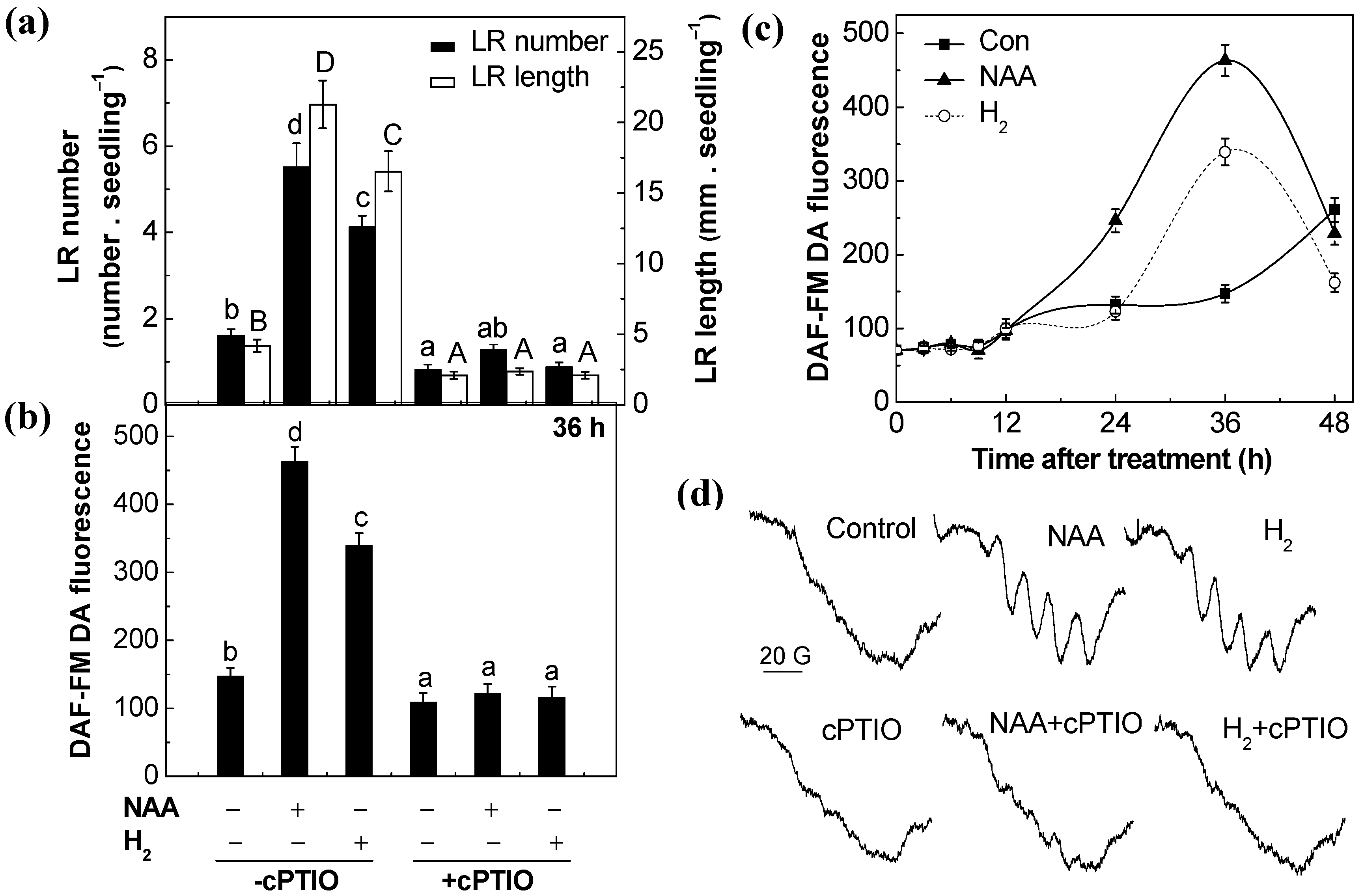
2.4. Nitric Oxide (NO) Was Involved in H2-Promoted Lateral Root Formation
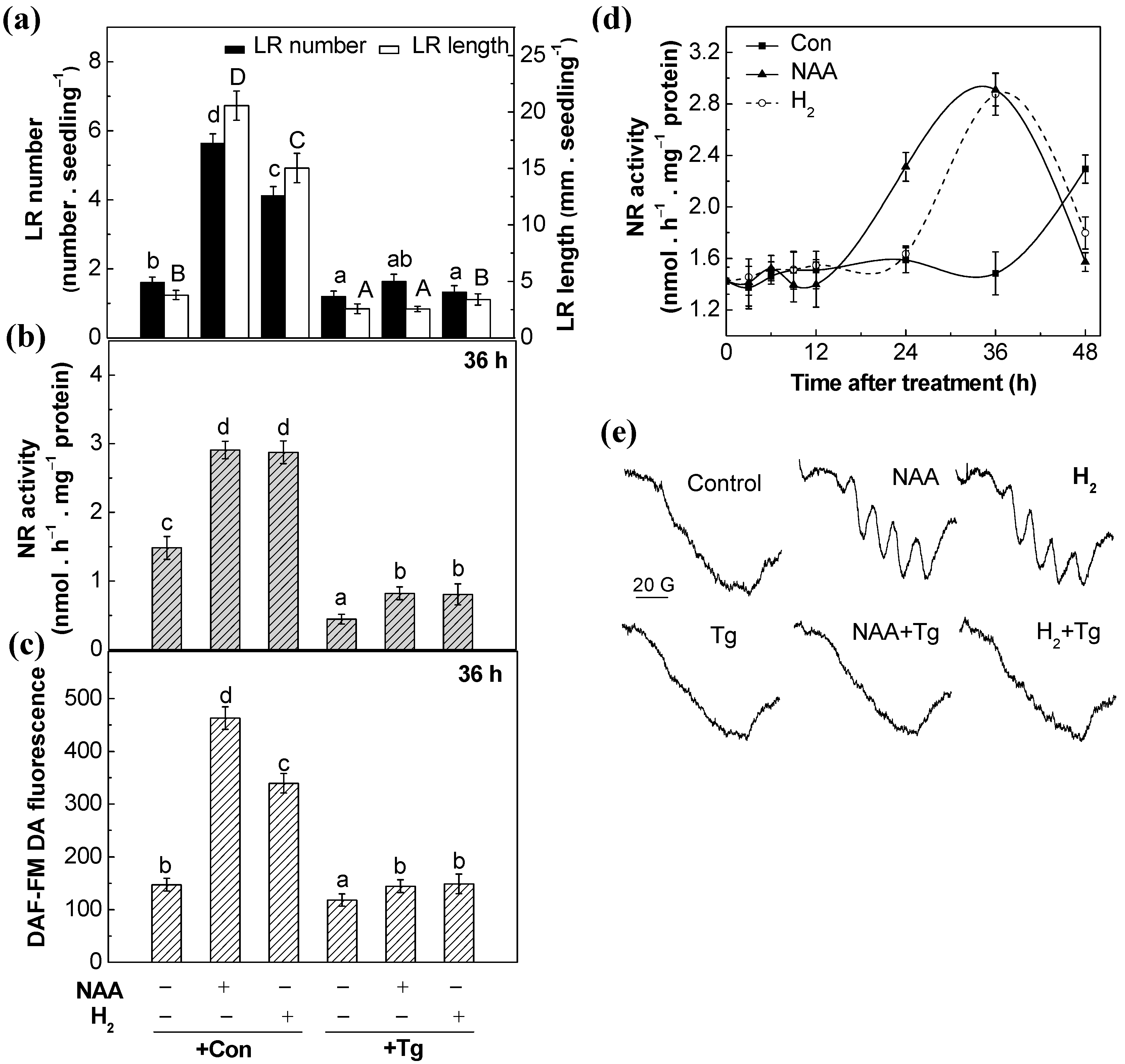
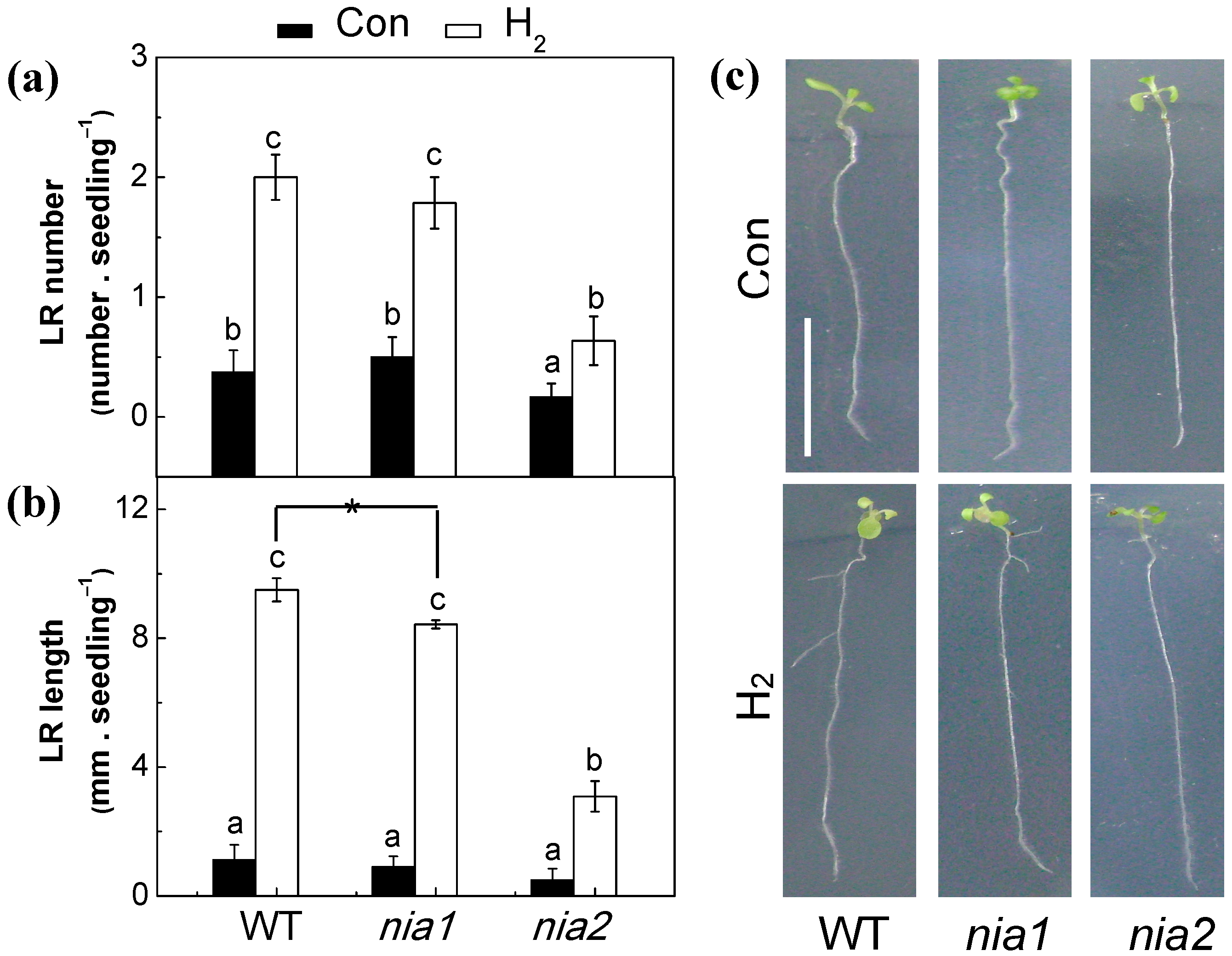
2.5. Nitrate Reductase Might Be the NO Enzymatic Source
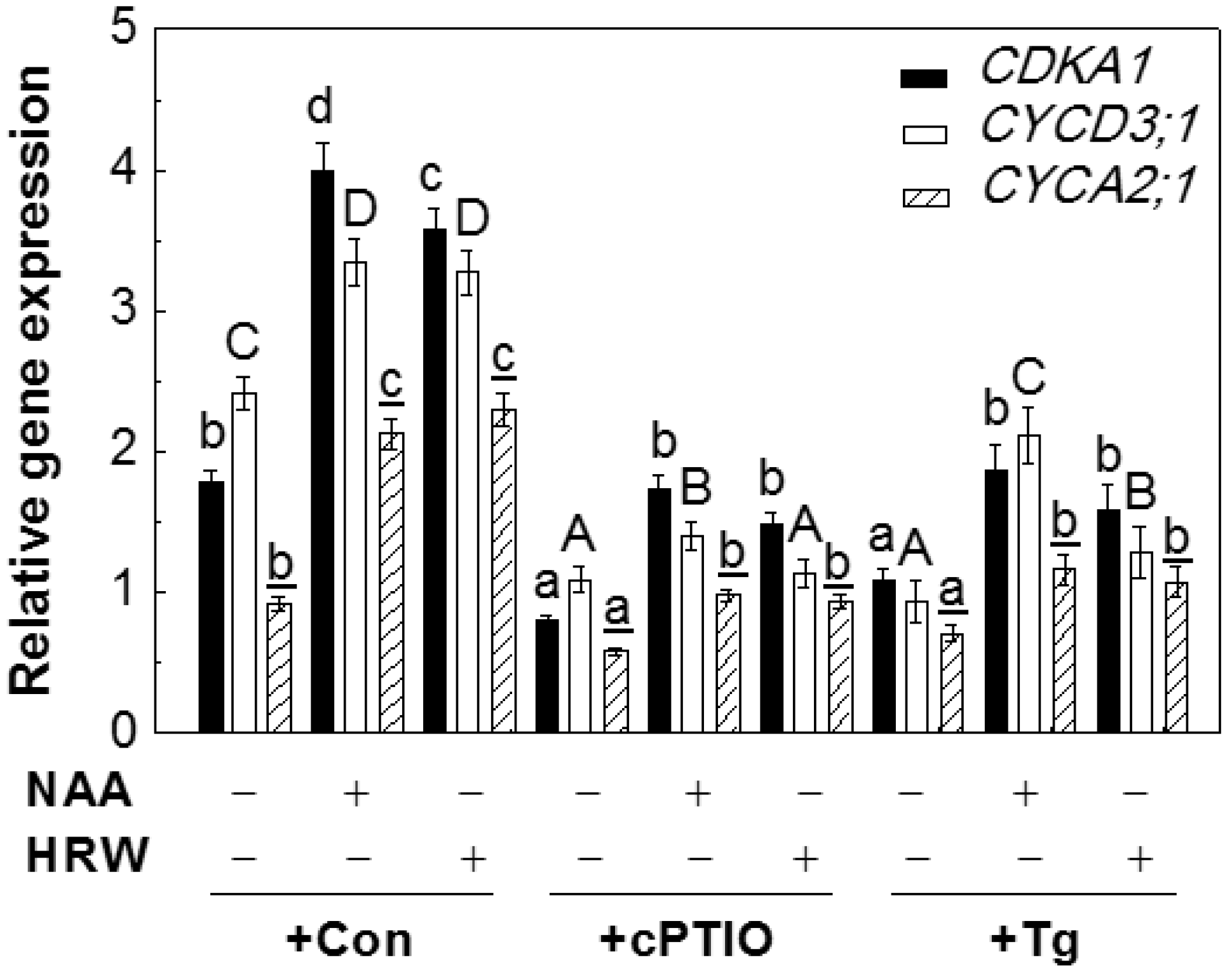
2.6. NO Participated in H2-Induced Transcript Levels of Cell Cycle Regulatory Genes
2.7. Genetic Evidence Supporting the Role of NR-Derived NO in H2-Induced Lateral Root Formation
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials and Growth Conditions
4.3. Preparation of H2-Rich Water
4.4. Measurement of Endogenous H2
4.5. Detection of Endogenous NO
4.6. Determination of Nitrate Reductase (NR) Activity
4.7. RNA Extraction and Real-Time Quantitative RT-PCR (qPCR) Analysis
4.8. Data Analysis
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
Abbreviations
| LR | lateral root |
| NAA | 1-naphthyl acetic acid |
| NPA | N-1-naphthylphthalamic acid |
| EPR | electron paramagnetic resonance |
| cPTIO | 2-4-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide |
| NR | nitrate reductase |
| NO | nitric oxide |
| NOS | nitric oxide synthetase |
| H2 | hydrogen gas |
| GC | gas chromatography |
| LSCM | laser scanning confocal microscopy |
| NAME | NG-nitro-l-arginine methyl ester hydrochloride |
| GSNOR | S-nitrosoglutathione reductase |
| Tg | tungstate |
References
- Casimiro, I.; Beeckman, T.; Graham, N.; Bhalerao, R.; Zhang, H.; Casero, P.; Sandberg, G.; Bennett, M.J. Dissecting Arabidopsis lateral root development. Trends Plant Sci. 2003, 8, 165–171. [Google Scholar] [CrossRef]
- Péret, B.; Rybe, B.; Casimiro, I.; Benková, E.; Swarup, R.; Laplaze, L.; Beeckman, T.; Bennett, M.J. Arabidopsis lateral root development: An emerging story. Trends Plant Sci. 2009, 14, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Lavenus, J.; Goh, T.; Roberts, I.; Guyomarch, S.; Lucas, M.; Smet, I.D. Lateral root development in Arabidopsis: Fifty shades of auxin. Trends Plant Sci. 2013, 18, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Perrine-Walker, F.M.; Jublanc, E. The localization of auxin transporters PIN3 and LAX3 during lateral root development in Arabidopsis thaliana. Biol. Plantarum. 2014, 58, 778–782. [Google Scholar] [CrossRef]
- Correa-Aragunde, N.; Graziano, M.; Chevalier, C.; Lamattina, L. Nitric oxide modulates the expression of cell cycle regulatory genes during lateral root formation in tomato. J. Exp. Bot. 2006, 57, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Graham, N.; Inzé, D.; Sandberg, G.; Casero, P.J. Auxin transport promotes Arabidopsis lateral root initiation. Plant Cell 2001, 13, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Correa-Aragunde, N.; Graziano, M.; Lamattina, L. Nitric oxide plays a central role in determining lateral root development in tomato. Planta 2004, 218, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.C.; Graziano, M.; Polacco, J.C.; Lamattina, L. Nitric oxide functions as a positive regulator of root hair development. Plant Signal. Behav. 2006, 1, 28–33. [Google Scholar] [CrossRef]
- Pagnussat, G.C.; Simontacchi, M.; Puntarulo, S.; Lamattina, L. Nitric oxide is required for root organogenesis. Plant Physiol. 2002, 129, 954–956. [Google Scholar] [CrossRef] [PubMed]
- Correa-Aragunde, N.; París, R.; Foresi, N.; Terrile, C.; Casalongué, C.; Lamattina, L. The auxin-nitric oxide highway: A right direction in determining the plant root system. In Gasotransmitters in Plants; Lamattina, L., García-Mata, C., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 117–136. [Google Scholar]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci. 2010, 16, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Renwick, G.M.; Giumarro, C.; Siegel, S.M. Hydrogen metabolism in higher plants. Plant Physiol. 1964, 39, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.S.; Kawamura, T.; Toyoda, Y.; Nakao, A. Recent advances in hydrogen research as a therapeutic medical gas. Free Radic. Res. 2010, 44, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.F.; Sun, X.J.; Xia, Z.F. Hydrogen resuscitation, a new cytoprotective approach. Clin. Exp. Pharmacol. Physiol. 2011, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Cai, J.; Liu, S.; Liu, Y.; Xu, W.; Tao, H.; Sun, X. Hydrogen-rich saline provides protection against hyperoxic lung injury. J. Surg. Res. 2011, 165, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cui, W.; Zhu, K.; Xie, Y.; Zhang, C.; Shen, W. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production. J. Hazard. Mater. 2014, 267, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, W.; Qi, F.; Cui, W.; Xie, Y.; Shen, W. Hydrogen-rich water regulates cucumber adventitious root development in a heme oxygenase-1/carbon monoxide-dependent manner. J. Plant Physiol. 2014, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ruiz, M.; Mioto, P.; Palma, J.M.; Corpas, F.J. S-nitrosoglutathione reductase (GSNOR) activity is down-regulated during pepper (Capsicum annuum L.) fruit ripening. Nitric Oxide 2017, 27, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Lamattina, L.; Garc´ıa-Mata, C.; Graziano, M.; Pagnussat, G. Nitric oxide: The versatility of an extensive signal molecule. Annul. Rev. Plant Biol. 2003, 54, 109–136. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Desikan, R.; Hancock, J.T. Nitric oxide signaling in plants. New Phytol. 2003, 159, 11–35. [Google Scholar] [CrossRef]
- Xie, Y.; Mao, Y.; Zhang, W.; Lai, D.; Wang, Q.; Shen, W. Reactive oxygen species dependent nitric oxide production contributes to hydrogen promoted stomatal closure in Arabidopsis. Plant Physiol. 2014, 165, 759–773. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Zhang, M.; Sun, X. Molecular hydrogen is involved in phytohormone signaling and stress response in plants. PLoS ONE 2013, 8, e71038. [Google Scholar] [CrossRef] [PubMed]
- Maimaiti, J.; Zhang, Y.; Yang, J.; Cen, Y.P.; Layzell, D.B.; Peoples, M.; Dong, Z. Isolation and characterization of hydrogen-oxidizing bacteria induced following exposure of soil to hydrogen gas and their impact on plant growth. Environ. Microbiol. 2007, 9, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Mao, Y.; Lai, D.; Zhang, W.; Shen, W. H2 enhances Arabidopsis salt tolerance by manipulating ZAT10/12-mediated antioxidant defence and controlling sodium exclusion. PLoS ONE 2012, 7, e49800. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhu, S.S.; Long, J.Y.; Wang, N.; Wang, R.; Shen, W. Hydrogen-rich water alleviates salt stress in rice during seed germination. Plant Soil 2013, 370, 47–57. [Google Scholar] [CrossRef]
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of heme oxygenase-1 signalling system. Plant Cell Environ. 2013, 36, 956–969. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Gao, C.; Fang, P.; Lin, G.; Shen, W. Alleviation of cadmium toxicity in Medicago sativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Fang, P.; Zhu, K.; Mao, Yu.; Gao, C.; Xie, Y.; Wang, J.; Shen, W.B. Hydrogen-rich water confers plant tolerance to mercury toxicity in alfalfa seedlings. Ecotoxicol. Environ. Saf. 2014, 5, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Cao, H.; Fang, W.; Pan, J.; Chen, J.; Zhang, J.; Shen, W. Linking hydrogen-enhanced rice aluminum tolerance with the reestablishment of GA/ABA balance and miRNA-modulated gene expression: A case study on germination. Ecotoxicol. Environ. Saf. 2017, 145, 303–312. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H.; Zhang, H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2003, 33, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Beligni, M.V.; Lamattina, L. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta 2000, 210, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Stohr, C.; Stremlau, S. Formation and possible roles of nitric oxide in plant roots. J. Exp. Bot. 2006, 57, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annul. Rev. Plant Biol. 2008, 59, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Lamattina, L.; Spoel, S.H.; Loake, G.J. Nitric oxide function in plant biology: A redox cue in deconvolution. New Phytol. 2014, 202, 1142–1156. [Google Scholar] [CrossRef] [PubMed]
- Sakihama, Y.; Nakamura, S.; Yamasaki, H. Nitric oxide production mediated by nitrate reductase in the green alga Chlamydomonas reinhardtii: an alternative NO production pathway in photosynthetic organisms. Plant Cell Physiol. 2002, 43, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Sakihama, Y. Simultaneous production of nitric oxide and peroxynitrite by plant nitrate reductase: In vitro evidence for the NR-dependent formation of active nitrogen species. FEBS Lett. 2000, 468, 89–92. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, L.; Zhang, L.; Zhang, W. Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, J.; Song, W.; Tao, J.; Huang, S.; Chen, S.; Zhang, Y. Nitric oxide generated by nitrate reductase increases nitrogen uptake capacity by inducing lateral root formation and inorganic nitrogen uptake under partial nitrate nutrition in rice. J. Exp. Bot. 2015, 66, 2449–2459. [Google Scholar] [CrossRef] [PubMed]
- Perrine-Walker, F.M.; Gartner, E.; Hocart, C.H.; Becker, A.; Rolfe, B.G. Rhizobium-initiated rice growth inhibition caused by nitric oxide accumulation. Mol. Plant Micorbe Interact. 2007, 20, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liao, W.; Niu, L.; Wang, M.; Ma, Z. Nitric oxide is involved in hydrogen gas-induced cell cycle activation during adventitious root formation in cucumber. BMC Plant Biol. 2016, 16, 146. [Google Scholar] [CrossRef] [PubMed]
- Himanen, K.; Boucheron, E.; Vanneste, S.; de Almeida Engler, J.; Inzé, D.; Beeckman, T. Auxin-mediated cell cycle activation during early lateral root initiation. Plant Cell 2002, 14, 2339–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Ma, X.; Zhang, Y.; Sun, T.; Xu, F.; Chen, Y.; Liu, X.; Yue, M. Role and interrelationship of Ga protein, hydrogen peroxide, and nitric oxide in ultraviolet B-induced stomatal closure in Arabidopsis leaves. Plant Physiol. 2013, 161, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Mao, Y.; Lai, D.; Zhang, W.; Zheng, T.; Shen, W. Roles of NIA/NR/ NOA1-dependent nitric oxide production and HY1 expression in the modulation of Arabidopsis salt tolerance. J. Exp. Bot. 2013, 64, 3045–3060. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Cao, Y.; Guo, P.; Tao, Y.; Zhao, B. Detection of nitric oxide in plants by electron spin resonance. Phytopathology 2004, 94, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.Z.; Nie, S.J.; Xing, D. Nitric oxide-mediated maintenance of redox homeostasis contributes to NPR1-dependent plant innate immunity triggered by lipopolysaccharides. Plant Physiol. 2012, 160, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Expósito-Rodríguez, M.; Borges, A.A.; Borges-Pérez, A.; Pérez, J.A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Boil. 2008, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Treatment | LR Number (seedling−1) | LR Length (mm seedling−1) | LRP Number (seedling−1) |
|---|---|---|---|---|
| Lycopersicon esculentum | Con | 1.89 ± 0.17 | 4.03 ± 0.48 | 1.25 ± 0.25 |
| 0.39 mM H2 | 4.13 ± 0.26 * | 15.03 ± 1.30 * | 3.38 ± 0.20 * | |
| Brassica napus | Con | 1.67 ± 0.38 | 4.00 ± 0.31 | 1.30 ± 0.11 |
| 0.39 mM H2 | 4.00 ± 0.26 * | 15.00 ± 0.51 * | 3.48 ± 0.20 * | |
| Brassica chinensis | Con | 1.74 ± 0.19 | 4.21 ± 0.43 | 2.17 ± 0.39 |
| 0.078 mM H2 | 5.14 ± 0.28 * | 16.62 ± 1.23 * | 3.58 ± 0.12 * | |
| Oryza sativa | Con | 5.60 ± 1.18 | 13.00 ± 2.74 | 1.50 ± 0.24 |
| 0.39 mM H2 | 12.05 ± 1.31 * | 38.50 ± 4.78 * | 2.92 ± 0.23 * | |
| Zea mays | Con | 16.67 ± 1.62 | 40.00 ± 3.85 | 15.73 ± 0.52 |
| 0.78 mM H2 | 32.41 ± 1.03 | 89.60 ± 2.48 * | 35.33 ± 2.00 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Z.; Duan, X.; Yao, P.; Cui, W.; Cheng, D.; Zhang, J.; Jin, Q.; Chen, J.; Dai, T.; Shen, W. Hydrogen Gas Is Involved in Auxin-Induced Lateral Root Formation by Modulating Nitric Oxide Synthesis. Int. J. Mol. Sci. 2017, 18, 2084. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102084
Cao Z, Duan X, Yao P, Cui W, Cheng D, Zhang J, Jin Q, Chen J, Dai T, Shen W. Hydrogen Gas Is Involved in Auxin-Induced Lateral Root Formation by Modulating Nitric Oxide Synthesis. International Journal of Molecular Sciences. 2017; 18(10):2084. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102084
Chicago/Turabian StyleCao, Zeyu, Xingliang Duan, Ping Yao, Weiti Cui, Dan Cheng, Jing Zhang, Qijiang Jin, Jun Chen, Tianshan Dai, and Wenbiao Shen. 2017. "Hydrogen Gas Is Involved in Auxin-Induced Lateral Root Formation by Modulating Nitric Oxide Synthesis" International Journal of Molecular Sciences 18, no. 10: 2084. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102084