Stem Cells as Potential Candidates for Psoriasis Cell-Replacement Therapy

 , ,
, ,
Abstract
:1. Introduction
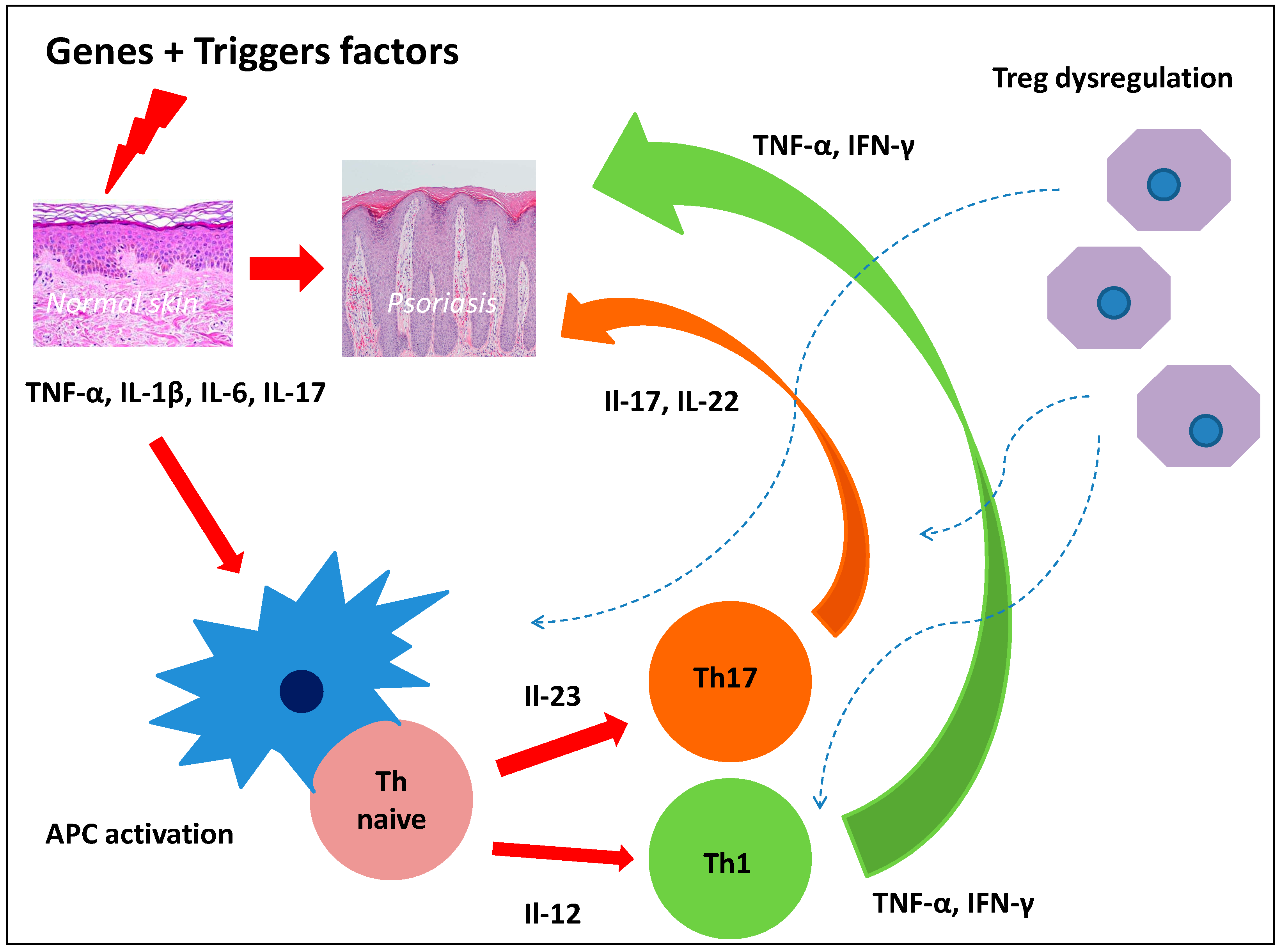
2. Pathogenesis of Psoriasis and Dermal Stem Cells
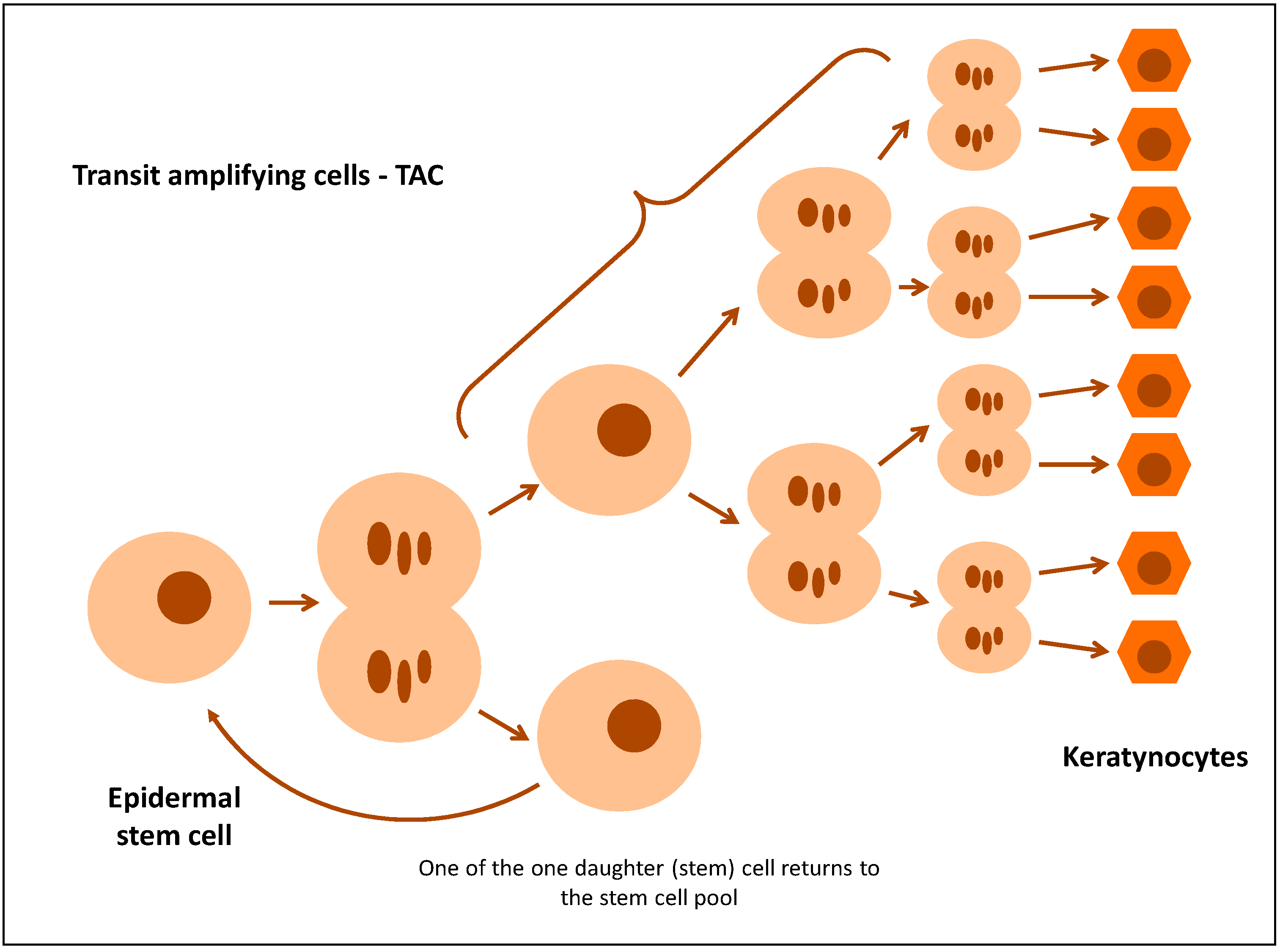
2.1. Epidermal Stem Cells in Psoriasis
2.2. Mesenchymal Stem Cells (MSC) in Psoriasis
2.3. Role of Telocytes in Psoriasis
3. Psoriasis and Bone Marrow Stem Cells
3.1. Associations of Psoriasis with Bone Marrow Cells
3.2. Proliferation of Bone Marrow Stem Cells (Progenitor Cells) in Psoriatic Patients Is Not Normal
3.3. T-Cells from Haematopoietic Cells of Bone Marrow of Psoriatic Patients Are Functionally Different from T-Cells in Healthy People
3.4. The Role of Bone Marrow Mesenchymal Stem Cells in Psoriatic Patients
3.5. Reduction of the Number of Progenitor Cells in Psoriatic Patients
4. Stem Cells in Psoriasis Treatment
4.1. Autologous Haematopoietic Stem Cell Transplantation
4.2. Umbilical Cord-Wharton’s Jelly Stem Cells
5. Conclusions
Conflicts of Interest
References
- Danielsen, K.; Olsen, A.O.; Wilsgaard, T.; Furberg, A.S. Is the prevalence of psoriasis increasing? A 30-year follow-up of a population-based cohort. Br. J. Dermatol. 2013, 168, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Symmons, D.P.; Griffiths, C.E.; Ashcroft, D.M. Identification and Management of Psoriasis and Associated ComorbidiTy (IMPACT) project team. Global epidemiology of psoriasis: A systematic review of incidence and prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Young, P.; Armstrong, A.W. An update on psoriasis and metabolic syndrome: A meta-analysis of observational studies. PLoS ONE 2017, 12, e0181039. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Taylor, C.; Kornmehl, H.; Armstrong, A.W. Psoriasis and suicidality: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2017, 77, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Randa, H.; Todberg, T.; Skov, L.; Larsen, L.S.; Zachariae, R. Health-related Quality of Life in Children and Adolescents with Psoriasis: A Systematic Review and Meta-analysis. Acta Derm. Venereol. 2017, 97, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Belinchón, I.; Rivera, R.; Blanch, C.; Comellas, M.; Lizán, L. Adherence, satisfaction and preferences for treatment in patients with psoriasis in the European Union: A systematic review of the literature. Patient Preference Adherence 2016, 10, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Chang, C.; Lu, Q. The inflammatory response in psoriasis: A comprehensive review. Clin. Rev. Allergy Immunol. 2016, 50, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.A.; Towne, J.E.; Kricorian, G.; Klekotka, P.; Gudjonsson, J.E.; Krueger, J.E.; Russel, C.B. The emerging role of IL-17 in the pathogenesis of psoriasis: Preclinical and clinical findings. J. Investig. Dermatol. 2013, 133, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.Y.; Shi, Y.; Luo, L.F.; Jiang, G.; Zhou, Q.; Xu, S.Z.; Lei, T.C. Asymmetric stem-cell division ensures sustained keratinocyte hyperproliferation in psoriatic skin lesions. Int. J. Mol. Med. 2016, 37, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Franssen, M.E.; Zeeuwen, P.L.; Vierwinden, G.; van de Kerkhof, P.C.; Schalkwijk, J.; van Erp, P.E. Phenotypical and functional differences in germinative subpopulations derived from normal and psoriatic epidermis. J. Investig. Dermatol. 2005, 124, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Joachimiak, R.; Bajek, A.; Drewa, T. Hair follicle as a novel source of stem cells. Postepy Hig. Med. Dosw. 2012, 66, 181–186. [Google Scholar] [CrossRef]
- Ma, D.R.; Yang, E.N.; Lee, S.T. A review: The location, molecular characterisation and multipotency of hair follicle epidermal stem cells. Ann. Acad. Med. Singap. 2004, 33, 784–788. [Google Scholar] [PubMed]
- O’Shaughnessy, R.F.; Seery, J.P.; Celis, J.E.; Frischauf, A.; Watt, F.M. PA-FABP, a novel marker of human epidermal transit amplifying cells revealed by 2D protein gel electrophoresis and cDNA array hybridisation. FEBS Lett. 2000, 486, 149–154. [Google Scholar] [CrossRef]
- Jones, P.; Simons, B.D. Epidermal homeostasis: Do committed progenitors work while stem cells sleep? Nat. Rev. Mol. Cell Biol. 2008, 9, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Watarai, A.; Amoh, Y.; Maejima, H.; Hamada, Y.; Katsuoka, K. Nestin expression is increased in the suprabasal epidermal layer in psoriasis vulgaris. Acta Derm. Venereol. 2013, 93, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Charruyer, A.; Fong, S.; Vitcov, G.G.; Sklar, S.; Tabernik, L.; Taneja, M.; Caputo, M.; Soeung, C.; Yue, L.; Uchida, Y.; et al. Brief report: Interleukin-17A-dependent asymmetric stem cell divisions are increased in human psoriasis: A mechanism underlying benign hyperproliferation. Stem Cells 2017, 35, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, T.; Saito, C.; Torii, K.; Nishida, E.; Yamazaki, S.; Morita, A. Photo(chemo)therapy reduces circulating Th17 cells and restores circulating regulatory T cells in psoriasis. PLoS ONE 2013, 8, e54895. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Reynolds, J.M.; Stout, R.D.; Bernlohr, D.A.; Suttles, J. Regulation of Th17 differentiation by epidermal fatty acid-binding protein. J. Immunol. 2009, 182, 7625–7633. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.M.; Liu, Q.; Brittingham, K.C.; Liu, Y.; Gruenthal, M.; Gorgun, C.Z.; Liu, Y.; Gruenthal, M.; Gorgun, C.Z.; Hotamisligil, G.S.; et al. Deficiency of fatty acid-binding proteins in mice confers protection from development of experimental autoimmune encephalomyelitis. J. Immunol. 2007, 179, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-González, L.H.; Ludwig, C.; Hohoff, C.; Rademacher, M.; Hanhoff, T.; Rüterjans, H.; Spener, F.; Lücke, C. Solution structure and backbone dynamics of human epidermal-type fatty acid-binding protein (E-FABP). Biochem. J. 2002, 15, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Katayama, I.; Nishioka, K. Possible contribution of stem cell factor in psoriasis vulgaris. J. Dermatol. Sci. 2000, 24, 171–176. [Google Scholar] [CrossRef]
- Liu, R.F.; Wang, F.; Wang, Q.; Zhao, X.C.; Zhang, K.M. Research note mesenchymal stem cells from skin lesions of psoriasis patients promote proliferation and inhibit apoptosis of HaCaT cells. Genet. Mol. Res. 2015, 14, 17758–17767. [Google Scholar] [CrossRef] [PubMed]
- Orciani, M.; Campanati, A.; Salvolini, E.; Lucarini, G.; Di Benedetto, G.; Offidani, A.; Di Primio, R. The mesenchymal stem cell profile in psoriasis. Br. J. Dermatol. 2011, 165, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Campanati, A.; Orciani, M.; Consales, V.; Lazzarini, R.; Ganzetti, G.; Di Benedetto, G.; Di Primio, R.; Offidani, A. Characterization and profiling of immunomodulatory genes in resident mesenchymal stem cells reflect the Th1-Th17/Th2 imbalance of psoriasis. Arch. Dermatol. Res. 2014, 306, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Yan, H.; Niu, X.; Chang, W.; An, P.; Wang, C.; Yang, Y.; Yan, X.; Li, J.; Liu, R.; et al. Gene expression profile of dermal mesenchymal stem cells from patients with psoriasis. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1782–1791. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.X.; Liu, R.F.; Zhao, X.C.; Jia, Y.R.; An, P.; Hao, Z.P.; Li, J.Q.; Li, X.H.; Yin, G.H.; Zhang, K.M. Increased miR-155-5p expression in dermal mesenchymal stem cells of psoriatic patients: Comparing the microRNA expression profile by microarray. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ren, G.; Cao, G.; Chen, Q.; Zheng, C.; Du, L.; Han, X.; Jiang, M.; Yang, Q.; Lin, L.; et al. miR-155 Regulates immune modulatory properties of mesenchymal stem cells by targeting TAK1-binding protein 2. J. Biol. Chem. 2013, 288, 11074–11079. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Huang, X.; Zhang, X.; Roensch, K.; Cao, Q.; Nakayama, K.I.; Blazar, B.R.; Zeng, Y.; Zhou, X. miR-221 and miR-155 regulate human dendritic cell development, apoptosis, and IL-12 production through targeting of p27kip1, KPC1, and SOCS-1. Blood 2011, 16, 4293–4303. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.J.; Wernicke, D.; Alder, H.; Costinean, S.; Volinia, S.; Croce, C.M. Mutator activity induced by microRNA-155 (miR-155) links inflammation and cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 4908–4913. [Google Scholar] [CrossRef] [PubMed]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. miR-155 regulates IFN-g production in natural killer cells. Blood 2012, 15, 3478–3485. [Google Scholar] [CrossRef] [PubMed]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Li, J.; Niu, X.; Liu, R.; Chang, W.; Zhao, X.; Wang, Q.; Li, X.; Yin, G.; Zhang, K. Stem cells in psoriasis. J. Dermatol. Sci. 2017, 86, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, Y.; Zhao, X.; Yang, Y.; Zhang, K. Lymphocyte inhibition is compromised in mesenchymal stem cells from psoriatic skin. Eur. J. Dermatol. 2014, 24, 560–567. [Google Scholar] [PubMed]
- Sah, S.K.; Park, K.H.; Yun, C.O.; Kang, K.S.; Kim, T.Y. Effects of human mesenchymal stem cells transduced with superoxide dismutase on imiquimod-induced psoriasis-like skin inflammation in mice. Antioxid. Redox Signal. 2016, 24, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Henno, A.; Blacher, S.; Lambert, C.; Colige, A.; Seidel, L.; Noël, A.; Lapière, C.; de la Brassinne, M.; Nusgens, B.V. Altered expression of angiogenesis and lymphangiogenesis markers in the uninvolved skin of plaque-type psoriasis. Br. J. Dermatol. 2009, 160, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Liu, R.; Niu, X.; Chang, W.; Yan, X.; Wang, C.; Li, J.; An, P.; Li, X.; Yin, G.; et al. Biological characteristics and gene expression pattern of bone marrow mesenchymal stem cells in patients with psoriasis. Exp. Dermatol. 2014, 23, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Chang, W.; Liu, R.; Hou, R.; Li, J.; Wang, C.; Li, X.; Zhang, K. Expression of pro-angiogenic genes in mesenchymal stem cells derived from dermis of patients with psoriasis. Int. J. Dermatol. 2016, 55, e280–e288. [Google Scholar] [CrossRef] [PubMed]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.M.; Simionescu, O. Telocytes in human skin-are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Manole, C.G.; Gherghiceanu, M.; Simionescu, O. Telocyte dynamics in psoriasis. J. Cell. Mol. Med. 2015, 19, 1504–1519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Li, X.; Yin, G.; Liu, Y.; Niu, X.; Hou, R. Functional characterization of CD4+CD25+ regulatory T cells differentiated in vitro from bone marrow-derived haematopoietic cells of psoriasis patients with a family history of the disorder. Br. J. Dermatol. 2008, 158, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Adkins, D.R.; Abidi, M.H.; Brown, R.A.; Khoury, H.; Goodnough, L.T.; Vij, R.; Westervelt, P.; DiPersio, J.F. Resolution of psoriasis after allogeneic bone marrow transplantation for chronic myelogenous leukemia: Late complications of therapy. Bone Marrow Transplant. 2000, 26, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J.; Wang, L.; Niu, X.; Hou, R.; Liu, R.; Hao, Z.; Wang, C.; Yin, G.; Zhang, K. Transmission of psoriasis by allogeneic bone marrow transplantation and blood transfusion. Blood Cancer J. 2015, 13, e288. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.C.; Mant, M.J. Amelioration of severe psoriasis with psoriatic arthritis for 20 years after allogeneic haematopoietic stem cell transplantation. Ann. Rheum. Dis. 2006, 65, 697. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Hou, R.; Niu, X.; Zhang, J.; Yin, G.; Li, X.; Jia, Y. Decreased colony formation of high proliferative potential colony-forming cells and granulocyte-macrophage colony-forming units and increased Hes-1 expression in bone marrow mononuclear cells from patients with psoriasis. Br. J. Dermatol. 2010, 163, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ablin, J.N.; Goldstein, Z.; Aloush, V.; Matz, H.; Elkayam, O.; Caspi, D.; Swartzenberg, S.; George, J.; Wohl, Y. Normal levels and function of endothelial progenitor cells in patients with psoriatic arthritis. Rheumatol. Int. 2009, 29, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Batycka-Baran, A.; Paprocka, M.; Krawczenko, A.; Kantor, A.; Duś, D.; Szepietowski, J.C. Reduced number of circulating endothelial progenitor cells (CD133+/KDR+) in patients with plaque psoriasis. Dermatology 2012, 22, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Głowińska-Olszewska, B.; Łuczyński, W.; Bossowski, A. Endothelial progenitor cells as a new marker of endothelial function with respect to risk of cardiovascular disorders. Postepy Hig. Med. Dosw. 2011, 65, 8–15. [Google Scholar] [CrossRef]
- Hayek, S.S.; Neuman, R.; Kavtaradze, N.; Sher, S.; Jones, D.; Li, Q.; Zhao, L.; Chen, S.C.; Waller, E.; Quyyumi, A. Tumor necrosis factor-alpha antagonism with etanercept improves endothelial progenitor cell counts in patients with psoriasis: Etanercept, vascular function and endothelial progenitor cells in psoriasis. Int. J. Cardiol. 2015, 182, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Hristov, M.; Erl, W.; Weber, P.C. Endothelial progenitor cells: Mobilization, differentiation, and homing. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Zurita, G.; Geffner, L.; Maldonado, B.; Uraga, E.; Armijos, L. Autologous Stem Cells Implants in Psoriasis. Available online: http://www.medicosecuador.com/espanol/articulos_medicos/ autologous_stem_cells_implants.htm (accessed on 20 September 2017).
- Kaffenberger, B.H.; Wong, H.K.; Jarjour, W.; Andritsos, L.A. Remission of psoriasis after allogeneic, but not autologous, hematopoietic stem-cell transplantation. J. Am. Acad. Dermatol. 2013, 68, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Al-Robaee, A.A.; Al-Zolibani, A.A.; Al-Shobili, H.A.; Kazamel, A.; Settin, A. IL-10 implications in psoriasis. Int. J. Health Sci. Qassim 2008, 2, 53–58. [Google Scholar] [PubMed]
- Flisiak, I.; Zaniewski, P.; Chodynicka, B. Plasma TGF-beta1, TIMP-1, MMP-1 and IL-18 as a combined biomarker of psoriasis activity. Biomarkers 2008, 13, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Zaher, H.; Shaker, O.G.; EL-Komy, M.H.; El-Tawdi, A.; Fawzi, M.; Kadry, D. Serum and tissue expression of transforming growth factor beta 1 in psoriasis. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Niu, J.W.; Ning, H.M.; Li, X.B.; Li, Y.; Wang, D.H.; Hu, L.D.; Sheng, H.X.; Xu, M.; Zhang, L.; et al. Treatment of psoriasis with Mesenchymal Stem Cells. Am. J. Med. 2016, 29, 13–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Staples, M.; Shinozuka, K.; Pantcheva, P.; Kang, S.D.; Borlongan, C.V. Wharton’s jelly-derived mesenchymal stem cells: Phenotypic characterization and optimizing their therapeutic potential for clinical applications. Int. J. Mol. Sci. 2013, 14, 11692–11712. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, S.J.; Gopalakrishnan, D.; Shankar, S.R.; Vasandan, A.B. Pro-inflammatory cytokines, IFNgamma and TNFalpha, influence immune properties of human bone marrow and Wharton jelly mesenchymal stem cells differentially. PLoS ONE 2010, 5, e9016. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, Q.; Wang, Z.; Tong, H.; Ma, L.; Zhang, Y.; Shan, F.; Meng, Y.; Yuan, Z. Comparative analysis of human mesenchymal stem cells from fetal-bone marrow, adipose tissue, and Warton’s jelly as sources of cell immunomodulatory therapy. Hum. Vaccin. Immunother. 2016, 12, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Chen, X.Y.; Li, J.; Zhang, H.Y.; Liu, J.; Sun, L.D. MiR-200a expression in CD4+ T cells correlates with the expression of Th17/Treg cells and relevant cytokines in psoriasis vulgaris: A case control study. Biomed. Pharmacother. 2017, 93, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, X.Q.; Cheng, J.; Hui, R.S.; Gao, T.W. Increased Th17 cells are accompanied by FoxP3(+) Treg cell accumulation and correlated with psoriasis disease severity. Clin. Immunol. 2010, 135, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Amari, A.; Ebtekar, M.; Moazzeni, S.M.; Soleimani, M.; Mohammadi-Amirabad, L.; Tahoori, M.T.; Massumi, M. In vitro generation of IL-35-expressing Human Wharton’s Jelly-derived Mesenchymal Stem Cells using lentiviral vector. Iran. J. Allergy Asthma Immunol. 2015, 14, 416–426. [Google Scholar] [PubMed]
- Aractingi, S.; Briand, N.; Le Danff, C.; Viguier, M.; Bachelez, H.; Michel, L.; Dubertret, R.; Carosella, E.D. HLA-G and NK receptors are expressed in psoriatic skin: A possible pathway for regulating infiltrating T cells? Am. J. Pathol. 2001, 159, 71–77. [Google Scholar] [CrossRef]
- Borghi, A.; Fogli, E.; Stignani, M.; Melchiorri, L.; Altieri, E.; Baricordi, O.; Rizzo, R.; Virgili, A. Soluble human leukocyte antigen-G and interleukin-10 levels in plasma of psoriatic patients: Preliminary study on a possible correlation between generalized immune status, treatments and disease. Arch. Dermatol. Res. 2008, 300, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Fong, C.Y.; Chak, L.L.; Biswas, A.; Tan, J.H.; Gauthaman, K.; Chan, W.K.; Bongso, A. Human Wharton’s jelly stem cells have unique transcriptome profiles compared to human embryonic stem cells and other mesenchymal stem cells. Stem Cell Rev. 2011, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- La Rocca, G.; Anzalone, R.; Corrao, S.; Magno, F.; Loria, T.; Lo Iacono, M.; Di Stefano, A.; Giannuzzi, P.; Marasà, L.; Cappello, F.; et al. Isolation and characterization of Oct-4+/HLA-G+ mesenchymal stem cells from human umbilical cord matrix: Differentiation potential and detection of new markers. Histochem. Cell Biol. 2009, 131, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Keijsers, R.R.; van der Velden, H.M.; van Erp, P.E.; de Boer-van Huizen, R.T.; Joosten, I.; Koenen, H.J.; van de Kerkhof, P.C. Balance of Treg vs. T-helper cells in the transition from symptomless to lesional psoriatic skin. Br. J. Dermatol. 2013, 168, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Owczarczyk-Saczonek, A.; Placek, W. Interleukin-17 as a factor linking the pathogenesis of psoriasis with metabolic disorders. Int. J. Dermatol. 2017, 56, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Rafei, M.; Campeau, P.M.; Aguilar-Mahecha, A.; Buchanan, M.; Williams, P.; Birman, E.; Yuan, S.; Young, Y.K.; Boivin, M.N.; Forner, K.; et al. Mesenchymal stromal cells ameliorate experimental autoimmune encephalomyelitis by inhibiting CD4 Thl7 T cells in a CC chemokine ligand 2-dependent manner. Immunology 2009, 182, 5994–6002. [Google Scholar] [CrossRef] [PubMed]
- Bovenschen, H.J.; van de Kerkhof, P.C.; van Erp, P.E.; Woestenenk, R.; Joosten, I.; Koenen, H.J. Foxp3+ regulatory T cells of psoriasis patients easily differentiate into IL-17A-producing cells and are found in lesional skin. J. Investig. Dermatol. 2011, 131, 1853–1860. [Google Scholar] [CrossRef] [PubMed]
- Alunno, A.; Montanucci, P.; Bistoni, O.; Basta, G.; Caterbi, S.; Pescara, T.; Pennoni, I.; Bini, V.; Bartoloni, E.; Gerli, R.; et al. In vitro immunomodulatory effects of microencapsulated umbilical cord Wharton jelly-derived mesenchymal stem cells in primary Sjögren’s syndrome. Rheumatol. Oxf. 2015, 54, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Karamehic, J.; Zecevic, L.; Resic, H.; Jukic, M.; Jukic, T.; Ridjic, O.; Panjeta, M.; Coric, J. Immunophenotype lymphocyte of peripheral blood in patients with psoriasis. Med. Arch. 2014, 68, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Khasawneh, A.; Baráth, S.; Medgyesi, B.; Béke, G.; Dajnoki, Z.; Gáspár, K.; Jenei, A.; Pogácsás, L.; Pázmándi, K.; Gaál, J.; et al. Myeloid but not plasmacytoid blood DCs possess Th1 polarizing and Th1/Th17 recruiting capacity in psoriasis. Immunol. Lett. 2017, 189, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Kastelan, M.; Prpić-Massari, L.; Brajac, I. Apoptosis in psoriasis. Acta Dermatovenerol. Croat. 2009, 17, 182–186. [Google Scholar] [PubMed]
- Han, I.; Yun, M.; Kim, E.O.; Kim, B.; Jung, M.H.; Kim, S.H. Umbilical cord tissue-derived mesenchymal stem cells induce apoptosis in PC-3 prostate cancer cells through activation of JNK and downregulation of PI3K/AKT signaling. Stem Cell Res. Ther. 2014, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Mori, J.; Kakihana, K.; Ohashi, K. Sustained remission of psoriasis vulgaris after allogeneic bone marrow transplantation. Br. J. Haematol. 2012, 159, 121. [Google Scholar] [CrossRef] [PubMed]
- Braiteh, F.; Hymes, S.R.; Giralt, S.A.; Jones, R. Complete remission of psoriasis after autologous hematopoietic stem-cell transplantation for multiple myeloma. J. Clin. Oncol. 2008, 26, 4511–4513. [Google Scholar] [CrossRef] [PubMed]
- Damien, P.; Allan, D.S. Regenerative therapy and immune modulation using Umbilical Cord Blood-Derived Cells. Biol. Blood Marrow Transplant. 2015, 21, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise review: Mesenchymal stem cells: Their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Batsali, A.K.; Kastrinaki, M.C.; Papadaki, H.A.; Pontikoglou, C. Mesenchymal stem cells derived from Wharton’s Jelly of the umbilical cord: Biological properties and emerging clinical applications. Curr. Stem Cell Res. Ther. 2013, 8, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.M.; Kalamegam, G.; Pushparaj, P.N.; Matta, C.; Memic, A.; Khademhosseini, A.; Mobasheri, R.; Poletti, F.L.; Hoyland, J.A.; Mobasheri, A. Mesenchymal stem cells in regenerative medicine: Focus on articularcartilage and intervertebral disc regeneration. Methods 2016, 99, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Sabapathy, V.; Sundaram, B.; Mankuzhy, P.; Kumar, S. Human Wharton’s Jelly Mesenchymal Stem Cells plasticity augments scar-free skin wound healing with hair growth. PLoS ONE 2014, 9, e93726. [Google Scholar] [CrossRef] [PubMed]
- Mohren, M.; Daikeler, T.; Benz, D.; Günaydin, I.; Kanz, L.; Kötter, I. Myeloablative immunosuppressive treatment with autologous haematopoietic stem cell transplantation in a patient with psoriatic arthropathy and monoclonal gammopathy of undetermined significance. Ann. Rheum. Dis. 2004, 63, 466–467. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Rahmetulla, R.; Loew, T.W.; Radhi, M.A. Complete resolution of guttate psoriasis following autologous SCT for Ewing’s sarcoma in a pediatric patient. Bone Marrow Transplant. 2012, 47, 1585–1586. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, Y.; Yamamoto, Y.; Ito, T.; Matsumoto, N.; Ichiyoshi, H.; Katsurada, T.; Date, M.; Ohga, S.; Kitajima, H.; Ikehara, S.; et al. Transfer of autoimmune thyroiditis and resolution of palmoplantar pustular psoriasis following allogeneic bone marrow transplantation. Bone Marrow Transplant. 1997, 19, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Rossi, H.A.; Becker, P.S.; Emmons, R.V.; Westervelt, P.; Levy, W.; Liu, Q.; Clark, Y.; Ballen, K. High-dose cyclophosphamide, BCNU, and VP-16 (CBV) conditioning before allogeneic stem cell transplantation for patients with non-Hodgkin's lymphoma. Bone Marrow Transplant. 2003, 31, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, H.; Tanaka, M.; Kawaguchi, H.; Yamaji, S.; Fujimaki, K.; Tomita, N.; Fujisawa, S.; Ishigatsubo, J. Resolution of psoriasis following allogeneic bone marrow transplantation for chronic myelogenous leukemia: Case report and review of the literature. Am. J. Hematol. 2002, 71, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Slavin, S.; Nagler, A.; Varadi, G.; Or, R. Graft vs autoimmunity following allogeneic non-myeloablative blood stem cell transplantation in a patient with chronic myelogenous leukemia and severe systemic psoriasis and psoriatic polyarthritis. Exp. Hematol. 2000, 28, 853–857. [Google Scholar] [CrossRef]
- Park, M.J.; Park, H.; Cho, M.L.; Oh, H.J.; Cho, Y.G.; Min, S.Y.; Chung, B.H.; Lee, J.W.; Kim, H.Y.; Cho, S.G. Transforming growth factor beta-transduced mesenchymal stem cells ameliorate experimental autoimmune arthritis through reciprocal regulation of Treg/Thl7 cells and osteoclastogenesis. Arthritis Rheum. 2011, 63, 1668–1680. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.D.; Hou, D.Y.; Liu, Y.; Koni, P.; Metz, R.; Chandler, P.; Mellor, A.L.; He, Y.; Munn, D.H. Indoleamine 2,3-dioxygenase controls conversion of Foxp3+ Tregs to TH17-like cells in tumor-draining lymph nodes. Blood 2009, 113, 6102–6111. [Google Scholar] [CrossRef] [PubMed]
- Rey Nores, J.E.; Bensussan, A.; Vita, N.; Stelter, F.; Arias, M.A.; Jones, M.; Lefort, S.; Borysiewicz, L.K.; Ferrara, P.; Labéta, M.O. Soluble CD14 acts as a negative regulator of human T cell activation and function. Eur. J. Immunol. 1999, 29, 265–276. [Google Scholar] [CrossRef]
- Kalaszczynska, I.; Ferdyn, K. Wharton’s jelly derived mesenchymal stem cells: Future of regenerative medicine? Recent findings and clinical significance. Biomed. Res. Int. 2015, 2015, 430847. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Ju, G.Q.; Du, T.; Zhu, Y.J.; Liu, G.H. Microvesicles derived from human umbilical cord Wharton’s jelly mesenchymal stem cells attenuate bladder tumor cell growth in vitro and in vivo. PLoS ONE 2013, 8, e61366. [Google Scholar] [CrossRef] [PubMed]
- Gotherstrom, C.; Ringden, O.; Tammik, C.; Zetterberg, E.; Westgren, M.; Le Blanc, K. Immunologic properties of human fetal mesenchymal stem cells. Am. J. Obstet. Gynecol. 2004, 190, 239–245. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials. Available online: https://clinicaltrials.gov/ (accessed on 20 September 2017).



{kind=link}
{kind=link}
| Element of Psoriasis Pathogenesis | Effect of Stem Cells | References |
|---|---|---|
| Deficiency of IL-10 and TGF-β in serum and skin [52,53,54] | WJSCs produce IL-10 and TGF-β | Chen et al., 2016 [55] Kim et al., 2013 [56] Prasanna et al., 2010 [57] Wang et al., 2016 [58] |
| Hyperactivity of Th17 and the dysfunction of Treg [59,60] | WJSCs produce IL-35, which induces the proliferation of Treg cell populations, reduces the activity of Th17 and Th1 cells | Amari et al., 2014 [61] |
| Significantly lower plasma levels of sHLA-G and IL-10 in psoriatic patients; the treatment of psoriasis leads to suppression of Th1 activation because of induced sHLA-G secretion via an IL-10-dependent pathway [62,63] | WJSCs express of HLA-G | Kim et al., 2013 [56] Prasanna et al., 2010 [57] Fong et al., 2011 [64] La Rocca et al., 2009 [65] |
| A crucial role of Th17 and IL-17 in psoriatic plaques and general inflammation [66,67] | MSCs inhibit the activity of the Thl7 cell, reducing the expression of interleukin IL-17 | Rafei et al., 2009 [68] |
| Decrease in FOXP3 and an increase in IL-17-producing Tregs; the conversion from Treg to IL-17/Treg is a continuum of converting cells, as evidenced by FOXP3+ RORγt+ co-expression and a gradual loss of FOXP3 [69] | WJSCs modulate FoxP3 and RORγt expression, leading to the conversion of Th17 into Treg cells | Alluno et al., 2014 [70] |
| Excessively aberrant Th1/Th2 homeostasis and Th17/Treg balance; the dysfunction of Treg [7,17,71,72] | WJ-MSCs inhibit the proliferative response of Th1 and Th17 but augment Th2 and Treg | Wang et al., 2016 [58] |
| Dendritic cells play a crucial role in the development of psoriatic inflammation, because of the production IL-12, IL-23, IL-6 [72] | WJSCs inhibit the maturation and activation of dendritic cell precursors | Kim et al., 2013 [56] |
| Psoriatic keratinocytes are particularly resistant to apoptosis, in psoriatic lesions over-expressed Bcl-XL, stimulated by TNF-α, is observed [73] | hWJSCs up-regulate the induction of apoptosis by attenuation of Bcl-2, Bcl-XL activation | Fong et al., 2011 [64] Han et al., 2014 [74] |
| Author | Patient | Psoriasis Course | Reason of HSCT | Myeloablative Chemotherapy | HSCT Type | Remission of Psoriasis | Comments |
|---|---|---|---|---|---|---|---|
| Adkins, 2000 [41] | K, 55 years old | Severe PS for 33 years, BSA > 60%, treated earlier with CsA, PUVA, MTX, with no improvement | CML | BU, CTX | Allo-HSCT | 2 years 4 months | Post-surgery period complicated with recurring infections and acute and chronic GVHD, treated with GCS, CsA and AZA. Died on 887th day following transplant because of pneumonia and AKF |
| Braiteh, 2008 [76] | M, 35 years old | PS and PsA for 15 years, BSA 50% | MM | L-PAM | Auto-HSCT | >2 years follow-up | 1 year of remission of MM |
| Mohren, 2004 [83] | M, 34 years old | PS and severe PsA for 15 years, ineffectively treated with MTX, CsA, MMF, sulfasalazine, NSAIDs and drugs in combination | PSA | CTX, L-PAM and selection of CD34+ cells from graft | PBSCT | 16 months | Mild recurring PSA, with good response to MTX. Also, history of monoclonal gammopathy IgAκ, resolved months following PBSCT, no recurrence. |
| Mori, 2012 [75] | M, 54 years old | PS for 10 years | MDS | BU, CTX | Allo-BMT | 8 months follow-up | |
| Woods, 2006 [43] | M, 29 years old | PS for 16 years, severe PSA for 1 year, heavily restricts performance | AA | CTX, radiotherapy | Allo-HSCT | 12 months PS 5 years PsA | The 20-year follow-up after HSCT showed a recurrence of mild psoriasis limited to head skin and recurrence of PSA, well-controlled with drugs and not causing significant disability. |
| Held, 2012 [84] | M, 9 years old | Guttate psoriasis, erythroderma | Edwing sarcoma | BU, L-PAM | Auto-SCT (ASCR) | 15 months follow-up | 13 months of remission of Edwing sarcoma |
| Kishimoto 1997 [85] | M, 40 years old | PPP following chemotherapy (DRB, 6-MP and BH-AC), treated with local GCS and etretinate, no improvement | AML | BU, CTX | Allo-HSCT | 2 years follow-up | 5 months after allo-HSCT the patient developed autoimmune thyroiditis and chronic GVHD, treated with CsA and GCS for 7 months with improvement. |
| Rossi, 2006 [86] | M, 27 years old | PS for 2 years, treated with local GCS | Acute AA | ATG, CTX | Allo-BMT | 10 years follow-up | Received short-term MTX and CsA for 314 days following BMT as a preventive measure against GVHD |
| Rossi, 2006 [86] | M, 50 years old | PS for 20 years (scalp, elbows) | NHL | BEAM regimen (BCNU, AC, ETO, L-PAM) | Auto-BMT | 21 months | After 21 months, recurring PS restricted to elbows |
| Kanamori, 2002 [87] | M, 49 years old | PS for 20 years, treated with GCS externally | CML | BU, CTX, AC | Allo-BMT | 2 years 6 months follow-up | Patient received short-term MTX and CsA for 150 days as a preventive measure against GVHD. After BMT, developed liver function disorder (probably related to chronic GVHD) |
| Slavin, 2000 [88] | M, 38 years old | Severe PS and PSA for 8 years, periodically treated with MTX and phototherapy | CML | FLU, ATG, BU | Allo-BMT NST | 2 years follow-up | Patient received CsA as a preventive measure against GVHD. 32 days after BMT there was recurrence of PS, PSA and CML. CsA was discontinued with a view to inducing GVL against CML and GVA against PSA. Within a month, the patient developed macular-papular eruptions, like in GVHD, treated with GCS. Symptoms of CML, PS and PSA were resolved. |
| Study | Application Method | Phase | Trial Institution and Country | NCT Number and Duration Period | |
|---|---|---|---|---|---|
| 1 | Safety and Efficacy of UC-MSCs in Patients With Psoriasis Vulgaris | Patients will receive six UC-MSCs infusions (1 × 106/kg). The first to fourth infusion will be given once a week for four weeks, then the last two infusions will be given once every two weeks. | I, II | Hospital to Academy of Military Medical Sciences, China | NCT02491658 2015–2016 |
| 2 | Safety of FURESTEM-CD Inj. in Patients With Moderate to Severe Plaque-type Psoriasis | Patients will receive FURESTEM-CD (allogeneic hUCB-MSC) injection subcutaneous: 5.0 × 107 cells, 1.0 × 107 cells and 2.0 × 108 cells for four weeks. | I | The Catholic Univ. Korea Seoul, St. Marry’s Hospital, Seoul, Republic of Korea | NCT02918123 2016–2020 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Owczarczyk-Saczonek, A.; Krajewska-Włodarczyk, M.; Kruszewska, A.; Placek, W.; Maksymowicz, W.; Wojtkiewicz, J. Stem Cells as Potential Candidates for Psoriasis Cell-Replacement Therapy. Int. J. Mol. Sci. 2017, 18, 2182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102182
Owczarczyk-Saczonek A, Krajewska-Włodarczyk M, Kruszewska A, Placek W, Maksymowicz W, Wojtkiewicz J. Stem Cells as Potential Candidates for Psoriasis Cell-Replacement Therapy. International Journal of Molecular Sciences. 2017; 18(10):2182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102182
Chicago/Turabian StyleOwczarczyk-Saczonek, Agnieszka, Magdalena Krajewska-Włodarczyk, Anna Kruszewska, Waldemar Placek, Wojciech Maksymowicz, and Joanna Wojtkiewicz. 2017. "Stem Cells as Potential Candidates for Psoriasis Cell-Replacement Therapy" International Journal of Molecular Sciences 18, no. 10: 2182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102182