Intermittent Administration of Parathyroid Hormone 1–34 Enhances Osteogenesis of Human Mesenchymal Stem Cells by Regulating Protein Kinase Cδ


Abstract
:1. Introduction
2. Results
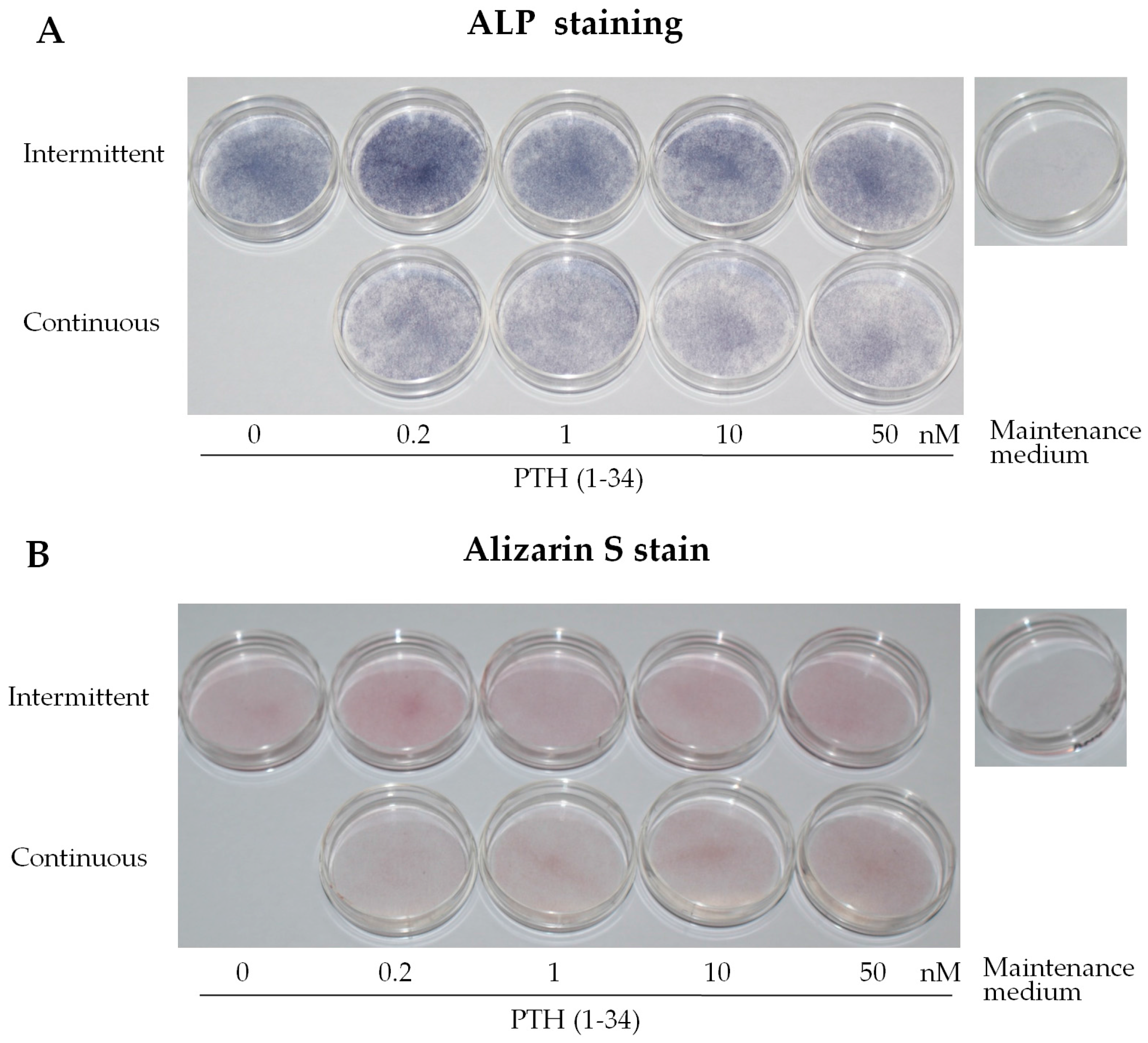
2.1. Intermittent PTH (1–34) Treatment Enhances Osteogenesis
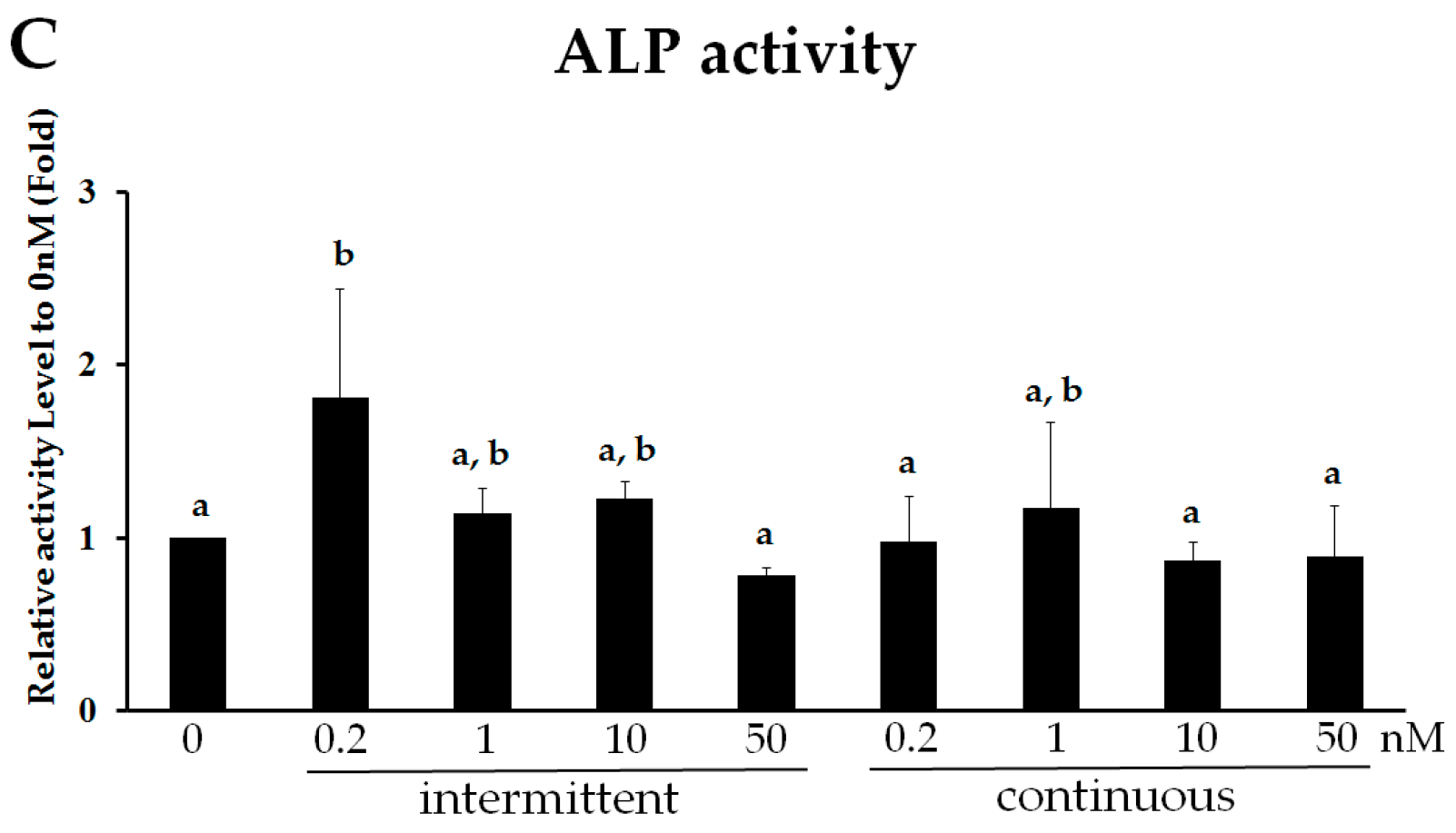
2.2. Intermittent PTH (1–34) Treatment Increases Early-Stage Osteoblast Activity during Osteogenic Differentiation
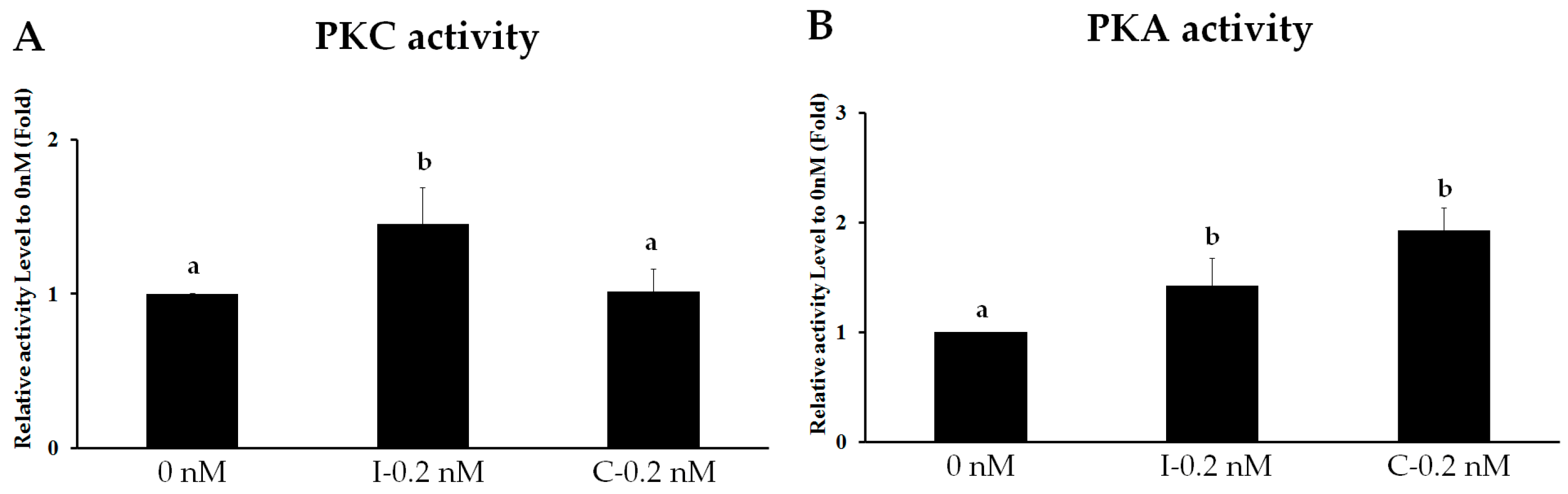
2.3. Intermittent PTH (1–34) Activates the PKC Pathway
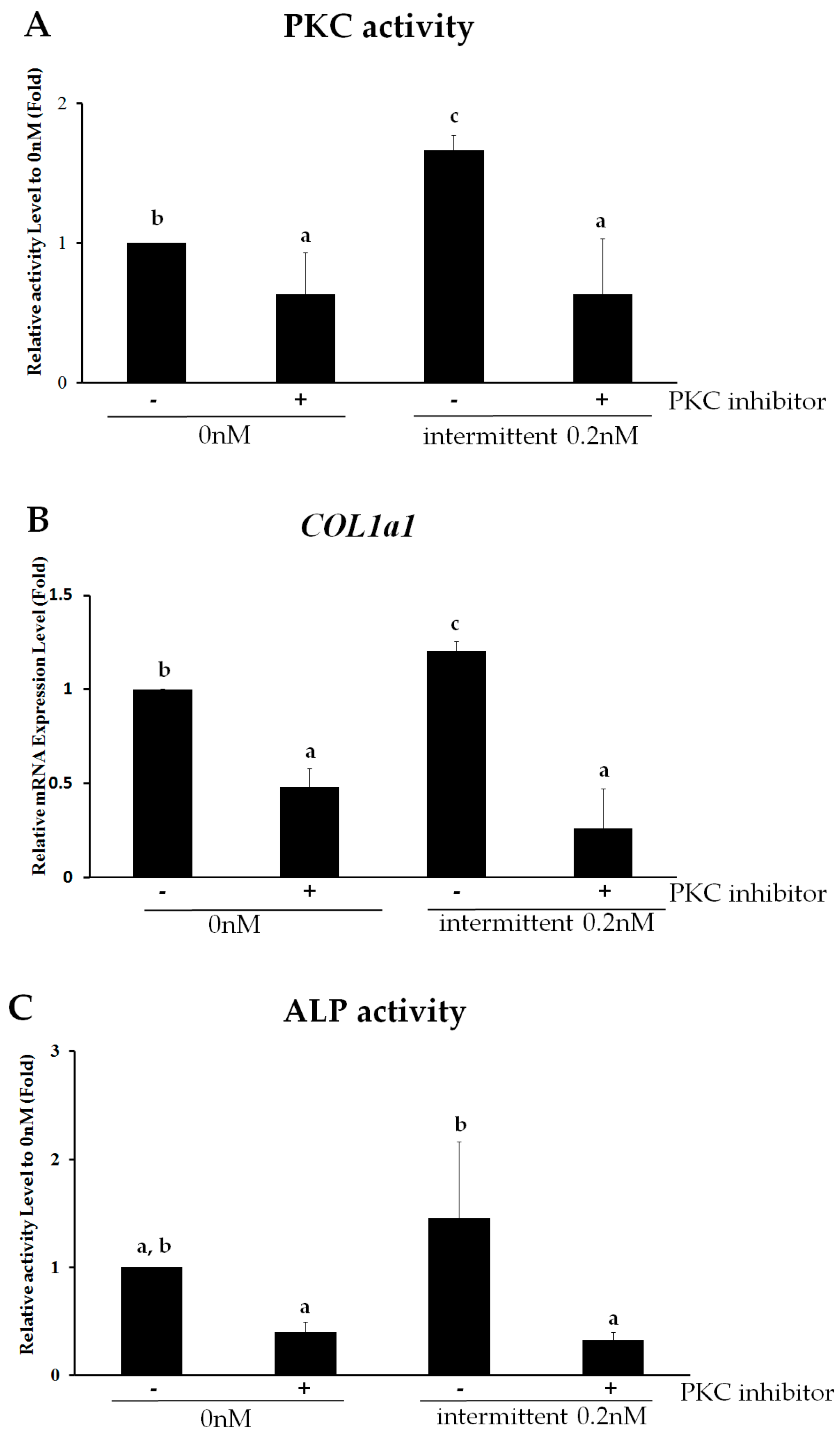
2.4. PKC Is Involved in Intermittent PTH (1–34)-Enhanced Osteogenesis
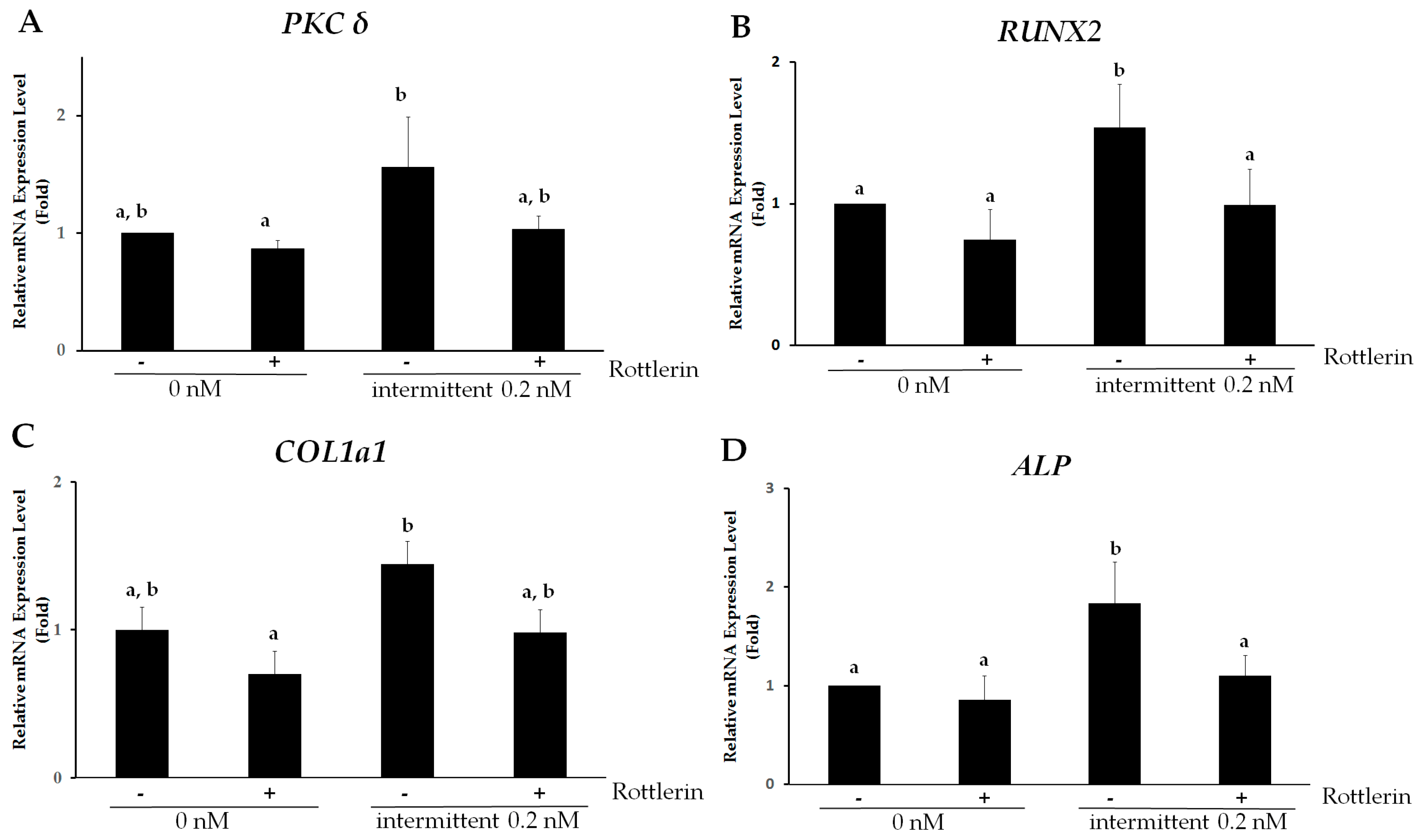
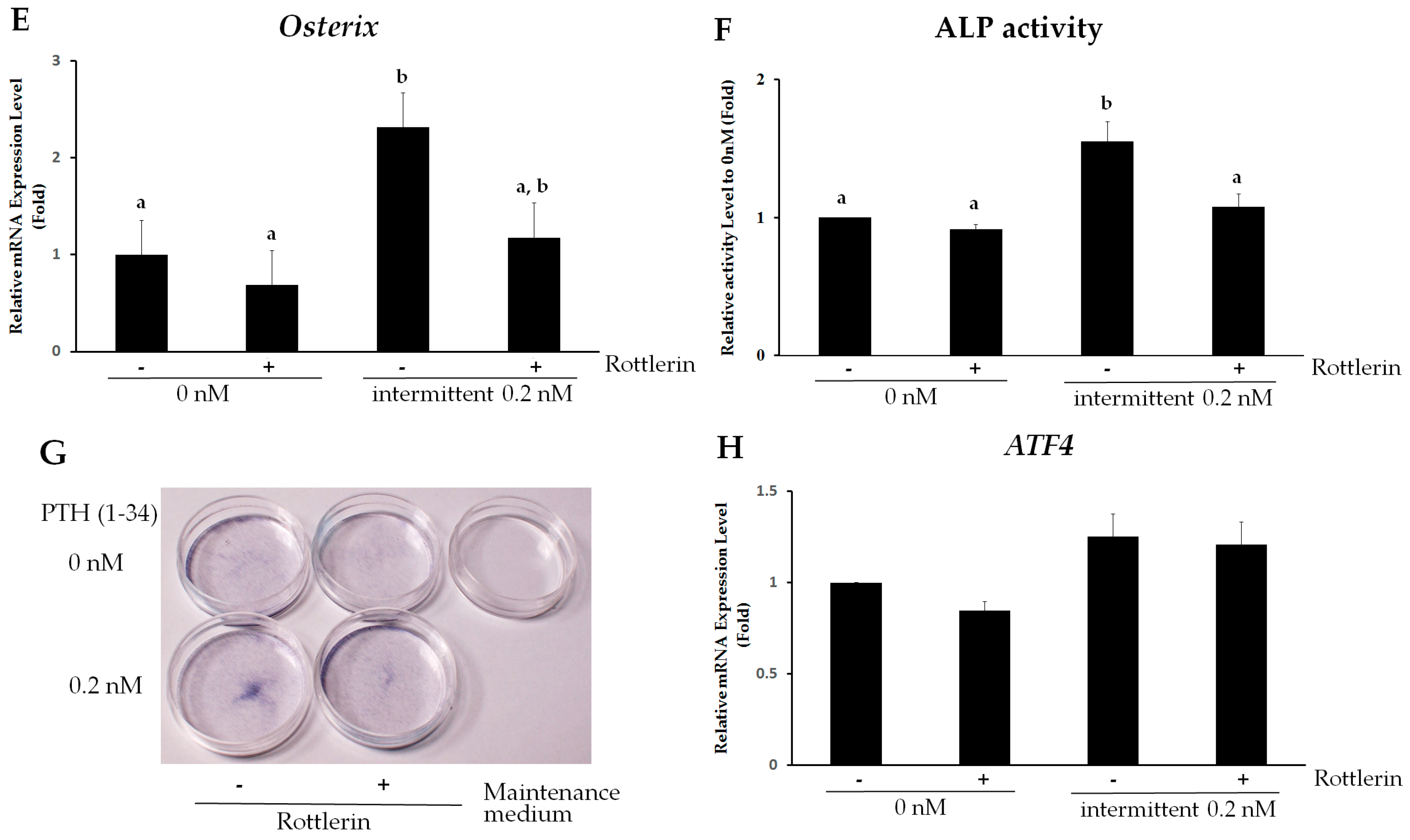
2.5. PKCδ Mediates Intermittent PTH (1–34)-Enhanced Osteogenesis of hMSCs
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Osteogenic Differentiation of hMSCs
4.2. PTH (1–34) Treatment
4.3. Inhibitors
4.4. Reverse Transcription and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.5. Alkaline Phosphatase (ALP) Staining
4.6. Alkaline Phosphatase (ALP) Activity Assay
4.7. Protein Kinase C (PKC) Kinase Activity Assay
4.8. Protein Kinase A (PKA) Kinase Activity Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal stromal cells |
| PTH | Parathyroid hormone |
| PKC | Protein kinase C |
| PKCδ | Protein kinase C δ |
| PKCα | Protein kinase C α |
| RUNX2 | Runt-related transcription factor 2 |
| COL1a1 | Collagen type 1 a1 |
| ALP | Alkaline phosphatase |
| OIM | Osteogenic induction medium |
| ATF4 | Activating transcription factor 4 |
References
- Osagie-Clouard, L.; Sanghani, A.; Coathup, M.; Briggs, T.; Bostrom, M.; Blunn, G. Parathyroid hormone 1–34 and skeletal anabolic action: The use of parathyroid hormone in bone formation. Bone Jt. Res. 2017, 6, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Neer, R.M.; Arnaud, C.D.; Zanchetta, J.R.; Prince, R.; Gaich, G.A.; Reginster, J.Y.; Hodsman, A.B.; Eriksen, E.F.; Ish-Shalom, S.; Genant, H.K.; et al. Effect of parathyroid hormone (1–34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N. Engl. J. Med. 2001, 344, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: Anabolic and catabolic actions on the skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Greenhill, C. Parathyroid function: Action of parathyroid hormone in osteocytes. Nat. Rev. Endocrinol. 2017, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wan, Q.; Yang, R.; Zhou, H.; Li, Z. Effects of intermittent versus continuous parathyroid hormone administration on condylar chondrocyte proliferation and differentiation. Biochem. Biophys. Res. Commun. 2012, 424, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Aslan, D.; Andersen, M.D.; Gede, L.B.; de Franca, T.K.; Jorgensen, S.R.; Schwarz, P.; Jorgensen, N.R. Mechanisms for the bone anabolic effect of parathyroid hormone treatment in humans. Scand. J. Clin. Lab. Investig. 2012, 72, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.C.; Costa, A.G.; Cusano, N.E.; Kousteni, S.; Bilezikian, J.P. Catabolic and anabolic actions of parathyroid hormone on the skeleton. J. Endocrinol. Investig. 2011, 34, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Juppner, H. Molecular cloning and characterization of a parathyroid hormone/parathyroid hormone-related peptide receptor: A member of an ancient family of G protein-coupled receptors. Curr. Opin. Nephrol. Hypertens. 1994, 3, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Vilardaga, J.P.; Romero, G.; Friedman, P.A.; Gardella, T.J. Molecular basis of parathyroid hormone receptor signaling and trafficking: A family B GPCR paradigm. Cell. Mol. Life Sci. 2011, 68, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cheloha, R.W.; Gellman, S.H.; Vilardaga, J.P.; Gardella, T.J. PTH receptor-1 signalling-mechanistic insights and therapeutic prospects. Nat. Rev. Endocrinol. 2015, 11, 712–724. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.S.; Abou-Samra, A.B. PTH and PTHrP signaling in osteoblasts. Cell. Signal. 2009, 21, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Swarthout, J.T.; D’Alonzo, R.C.; Selvamurugan, N.; Partridge, N.C. Parathyroid hormone-dependent signaling pathways regulating genes in bone cells. Gene 2002, 282, 1–17. [Google Scholar] [CrossRef]
- Siddappa, R.; Martens, A.; Doorn, J.; Leusink, A.; Olivo, C.; Licht, R.; van Rijn, L.; Gaspar, C.; Fodde, R.; Janssen, F.; et al. cAMP/PKA pathway activation in human mesenchymal stem cells in vitro results in robust bone formation in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 7281–7286. [Google Scholar] [CrossRef] [PubMed]
- Lampasso, J.D.; Marzec, N.; Margarone, J., 3rd; Dziak, R. Role of protein kinase C α in primary human osteoblast proliferation. J. Bone Miner. Res. 2002, 17, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Demiralp, B.; Schneider, A.; Koh, A.J.; Silve, C.; Wang, C.Y.; McCauley, L.K. Parathyroid hormone and parathyroid hormone-related protein exert both pro- and anti-apoptotic effects in mesenchymal cells. J. Biol. Chem. 2002, 277, 19374–19381. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Han, D.I.; Rhee, Y.H.; Jeong, S.J.; Kim, S.H.; Park, Y.G. Protein kinase C βII and δ/θ play critical roles in bone morphogenic protein-4-stimulated osteoblastic differentiation of MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 2010, 403, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, J.H.; Bae, S.C.; Choi, J.Y.; Kim, H.J.; Ryoo, H.M. The protein kinase C pathway plays a central role in the fibroblast growth factor-stimulated expression and transactivation activity of Runx2. J. Biol. Chem. 2003, 278, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Mellor, H.; Parker, P.J. The extended protein kinase C superfamily. Biochem. J. 1998, 332, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C. Regulation of the ABC kinases by phosphorylation: Protein kinase C as a paradigm. Biochem. J. 2003, 370, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Nakura, A.; Higuchi, C.; Yoshida, K.; Yoshikawa, H. PKCα suppresses osteoblastic differentiation. Bone 2011, 48, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Cho, H.Y.; Bui, H.T.; Kang, D. The osteogenic or adipogenic lineage commitment of human mesenchymal stem cells is determined by protein kinase C δ. BMC Cell Biol. 2014, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Franceschi, R.T.; Luo, M.; Fan, J.; Jiang, D.; Cao, H.; Kwon, T.G.; Lai, Y.; Zhang, J.; Patrene, K.; et al. Critical role of activating transcription factor 4 in the anabolic actions of parathyroid hormone in bone. PLoS ONE 2009, 4, e7583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Karsenty, G. ATF4, the osteoblast accumulation of which is determined post-translationally, can induce osteoblast-specific gene expression in non-osteoblastic cells. J. Biol. Chem. 2004, 279, 47109–47114. [Google Scholar] [CrossRef] [PubMed]
- Uzawa, T.; Hori, M.; Ejiri, S.; Ozawa, H. Comparison of the effects of intermittent and continuous administration of human parathyroid hormone(1–34) on rat bone. Bone 1995, 16, 477–484. [Google Scholar] [CrossRef]
- Ogita, M.; Rached, M.T.; Dworakowski, E.; Bilezikian, J.P.; Kousteni, S. Differentiation and proliferation of periosteal osteoblast progenitors are differentially regulated by estrogens and intermittent parathyroid hormone administration. Endocrinology 2008, 149, 5713–5723. [Google Scholar] [CrossRef] [PubMed]
- Di Bernardo, G.; Galderisi, U.; Fiorito, C.; Squillaro, T.; Cito, L.; Cipollaro, M.; Giordano, A.; Napoli, C. Dual role of parathyroid hormone in endothelial progenitor cells and marrow stromal mesenchymal stem cells. J. Cell. Phys. 2010, 222, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, Y.; Fu, Q.; He, M. Parathyroid hormone regulates osteoblast differentiation in a Wnt/β-catenin-dependent manner. Mol. Cell. Biochem. 2011, 355, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Sammons, J.; Ahmed, N.; El-Sheemy, M.; Hassan, H.T. The role of BMP-6, IL-6, and BMP-4 in mesenchymal stem cell-dependent bone development: Effects on osteoblastic differentiation induced by parathyroid hormone and vitamin D3. Stem Cells Dev. 2004, 13, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Satterwhite, J.; Heathman, M.; Miller, P.D.; Marin, F.; Glass, E.V.; Dobnig, H. Pharmacokinetics of teriparatide (rhPTH[1–34]) and calcium pharmacodynamics in postmenopausal women with osteoporosis. Calcif. Tissue Int. 2010, 87, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Rickard, D.J.; Wang, F.L.; Rodriguez-Rojas, A.M.; Wu, Z.; Trice, W.J.; Hoffman, S.J.; Votta, B.; Stroup, G.B.; Kumar, S.; Nuttall, M.E. Intermittent treatment with parathyroid hormone (PTH) as well as a non-peptide small molecule agonist of the PTH1 receptor inhibits adipocyte differentiation in human bone marrow stromal cells. Bone 2006, 39, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Bueno, E.M.; Kim, S.W.; Amato, I.; Shen, L.; Hahne, J.; Bleiberg, I.; Morley, P.; Glowacki, J. Effects of age on parathyroid hormone signaling in human marrow stromal cells. Aging Cell 2011, 10, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Rubin, M.R.; Cosman, F.; Lindsay, R.; Bilezikian, J.P. The anabolic effects of parathyroid hormone. Osteoporos. Int. 2002, 13, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Dobnig, H. A review of teriparatide and its clinical efficacy in the treatment of osteoporosis. Expert Opin. Pharmacother. 2004, 5, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Yamaguchi, A.; Tanizawa, T.; Endo, N.; Mashiba, T.; Uchiyama, Y.; Suda, T.; Yoshiki, S.; Takahashi, H.E. Increased bone formation by intermittent parathyroid hormone administration is due to the stimulation of proliferation and differentiation of osteoprogenitor cells in bone marrow. Bone 1994, 15, 717–723. [Google Scholar] [CrossRef]
- Shimizu, M.; Potts, J.T., Jr.; Gardella, T.J. Minimization of parathyroid hormone. Novel amino-terminal parathyroid hormone fragments with enhanced potency in activating the type-1 parathyroid hormone receptor. J. Biol. Chem. 2000, 275, 21836–21843. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chan, E.; Wang, S.X.; Li, B. Activation of p38 mitogen-activated protein kinase is required for osteoblast differentiation. Endocrinology 2003, 144, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, A.; Cheng, S.L.; Avioli, L.V.; Civitelli, R. Structure-function relationship of parathyroid hormone: Activation of phospholipase-C, protein kinase-A and -C in osteosarcoma cells. Endocrinology 1992, 130, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Dean, T.; Vilardaga, J.P.; Potts, J.T., Jr.; Gardella, T.J. Altered selectivity of parathyroid hormone (PTH) and PTH-related protein (PTHrP) for distinct conformations of the PTH/PTHrP receptor. Mol. Endocrinol. 2008, 22, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Gesty-Palmer, D.; Chen, M.; Reiter, E.; Ahn, S.; Nelson, C.D.; Wang, S.; Eckhardt, A.E.; Cowan, C.L.; Spurney, R.F.; Luttrell, L.M.; et al. Distinct β-arrestin- and G protein-dependent pathways for parathyroid hormone receptor-stimulated ERK1/2 activation. J. Biol. Chem. 2006, 281, 10856–10864. [Google Scholar] [CrossRef] [PubMed]
- Janulis, M.; Tembe, V.; Favus, M.J. Role of protein kinase C in parathyroid hormone stimulation of renal 1,25-dihydroxyvitamin D3 secretion. J. Clin. Investig. 1992, 90, 2278–2283. [Google Scholar] [CrossRef] [PubMed]
- Iida-Klein, A.; Varlotta, V.; Hahn, T.J. Protein kinase C activity in UMR-106–01 cells: Effects of parathyroid hormone and insulin. J. Bone Miner. Res. 1989, 4, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, G.N.; Rodrigues, T.L.; de Souza, A.P.; Line, S.R.; Marques, M.R. Parathyroid hormone (1–34) modulates odontoblast proliferation and apoptosis via PKA and PKC-dependent pathways. Calcif. Tissue Int. 2014, 95, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Halladay, D.L.; Miles, R.R.; Thirunavukkarasu, K.; Chandrasekhar, S.; Martin, T.J.; Onyia, J.E. Identification of signal transduction pathways and promoter sequences that mediate parathyroid hormone 1–38 inhibition of osteoprotegerin gene expression. J. Cell. Biochem. 2001, 84, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mochly-Rosen, D.; Das, K.; Grimes, K.V. Protein kinase C, an elusive therapeutic target? Nat. Rev. Drug Discov. 2012, 11, 937–957. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Singh, R.; Divieti, P.; Guo, J.; Bouxsein, M.L.; Bringhurst, F.R. Contributions of parathyroid hormone (PTH)/PTH-related peptide receptor signaling pathways to the anabolic effect of PTH on bone. Bone 2007, 40, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.; Joeng, K.S.; Nakayama, K.I.; Nakayama, K.; Rajagopal, J.; Carroll, T.J.; McMahon, A.P.; Long, F. Noncanonical Wnt signaling through G protein-linked PKCδ activation promotes bone formation. Dev. Cell 2007, 12, 113–127. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Oligonucleotide Sequence | Probe Number |
|---|---|---|
| RUNX2 | 5′–CTACCACCCCGCTGTCTTC–3′/5′–CAGAGGTGGCAGTGTCATCA–3′ | 29 |
| COL1a1 | 5′–ATGTTCAGCTTTGTGGACCTC–3′/5′–CTGTACGCAGGTGATTGGTG–3′ | 15 |
| ALP | 5′–AGAACCCCAAAGGCTTCTTC–3′/5′–CTTGGCTTTTCCTTCATGGT–3′ | 31 |
| Osterix | 5′–GACTGCAGAGCAGGTTCCTC-3′/5′–TAACCTGATGGGGTCATGGT–3′ | 43 |
| PKCδ | 5′–TCGACTGGGAAAAACTGGAG–3′/5′–CTTGGTTGGTTCCCTTTCCA–3′ | 80 |
| PKCα | 5’–ATTATCCCCGCTGGATCAC–3′/5′–CTCTGCTCCTTTGCCACAC–3′ | 83 |
| ATF4 | 5′–TGGTCAGTCCCTCCAACAAC–3′/5′–CTATACCCAACAGGGCATCC–3′ | 88 |
| GAPDH | 5′–GCTCTCTGCTCCTCCTGTTC–3′/5′–ACGACCAAATCCGTTGACTC–3′ | 60 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, S.-W.; Rimando, M.G.; Liu, Y.-S.; Lee, O.K. Intermittent Administration of Parathyroid Hormone 1–34 Enhances Osteogenesis of Human Mesenchymal Stem Cells by Regulating Protein Kinase Cδ. Int. J. Mol. Sci. 2017, 18, 2221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102221
Kuo S-W, Rimando MG, Liu Y-S, Lee OK. Intermittent Administration of Parathyroid Hormone 1–34 Enhances Osteogenesis of Human Mesenchymal Stem Cells by Regulating Protein Kinase Cδ. International Journal of Molecular Sciences. 2017; 18(10):2221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102221
Chicago/Turabian StyleKuo, Shu-Wen, Marilyn G. Rimando, Yi-Shiuan Liu, and Oscar K. Lee. 2017. "Intermittent Administration of Parathyroid Hormone 1–34 Enhances Osteogenesis of Human Mesenchymal Stem Cells by Regulating Protein Kinase Cδ" International Journal of Molecular Sciences 18, no. 10: 2221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18102221