Antibody-Based Protective Immunity against Helminth Infections: Antibody Phage Display Derived Antibodies against BmR1 Antigen


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
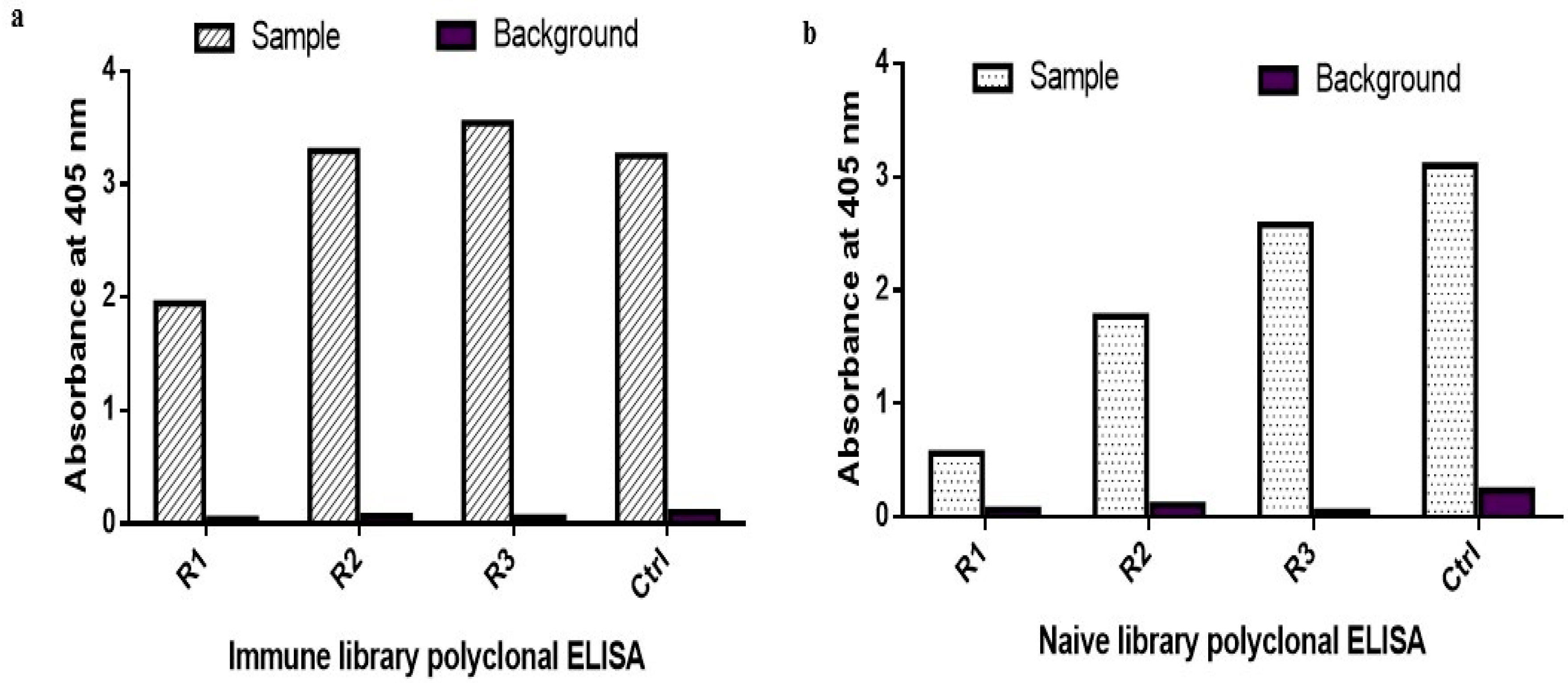
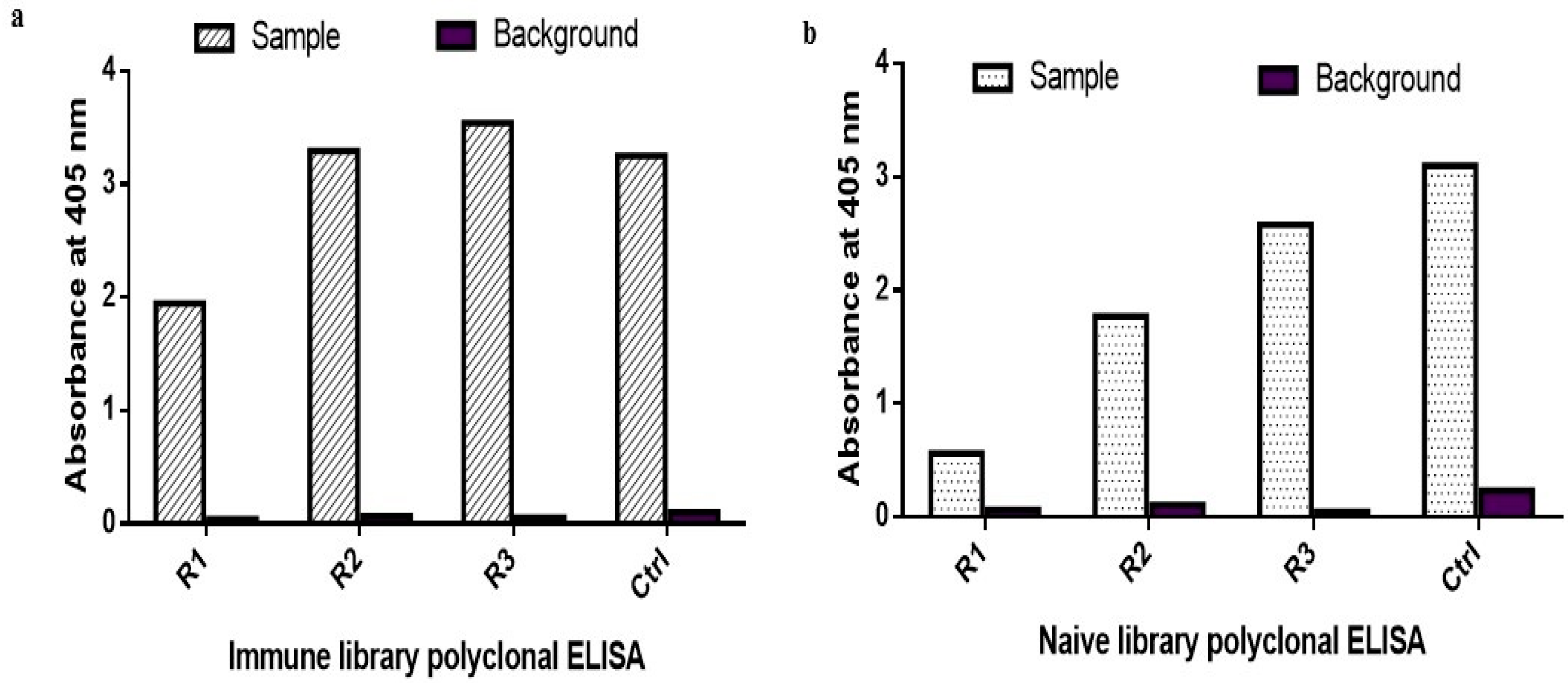
2.1. Biopanning and Polyclonal ELISA
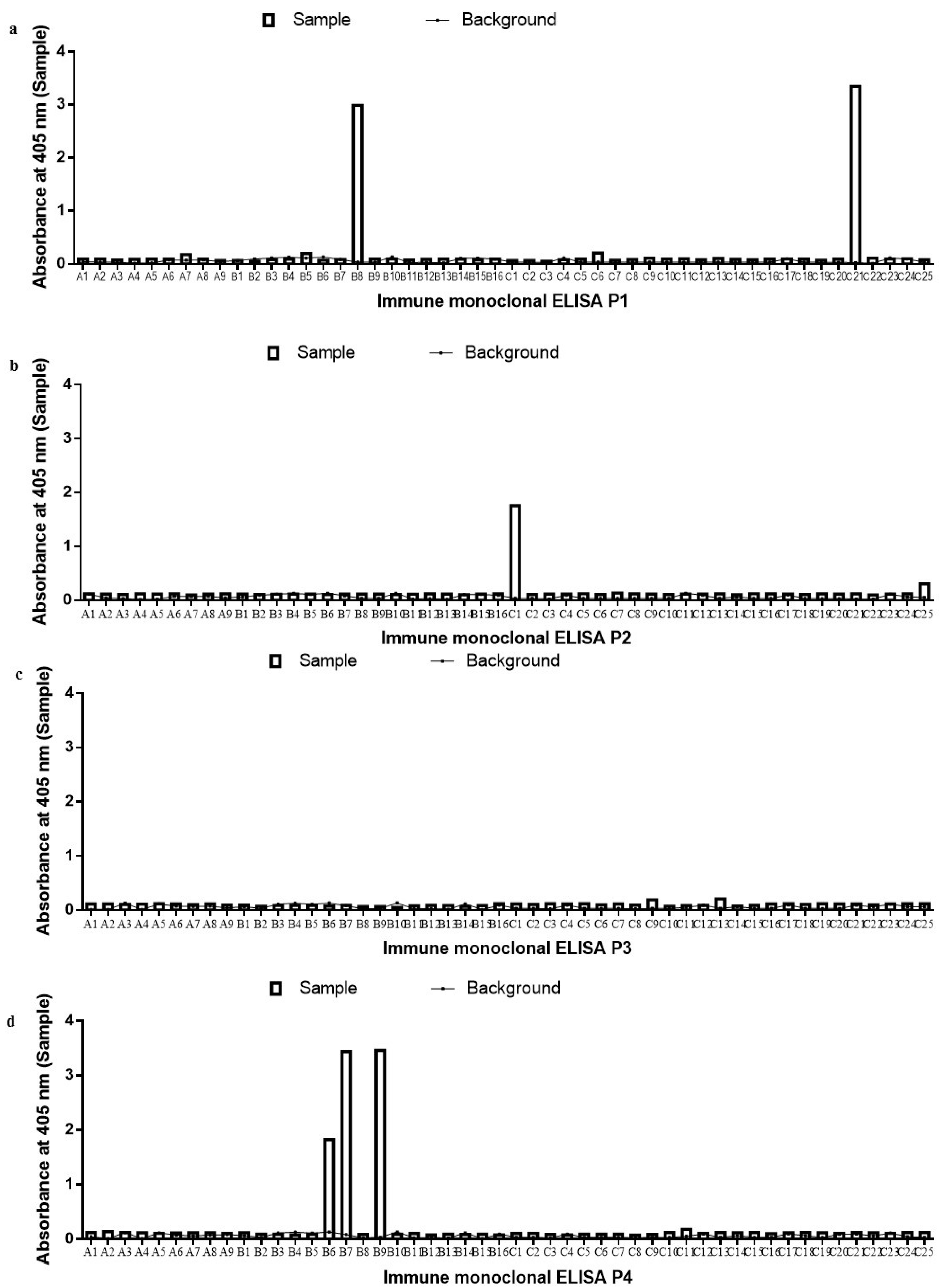
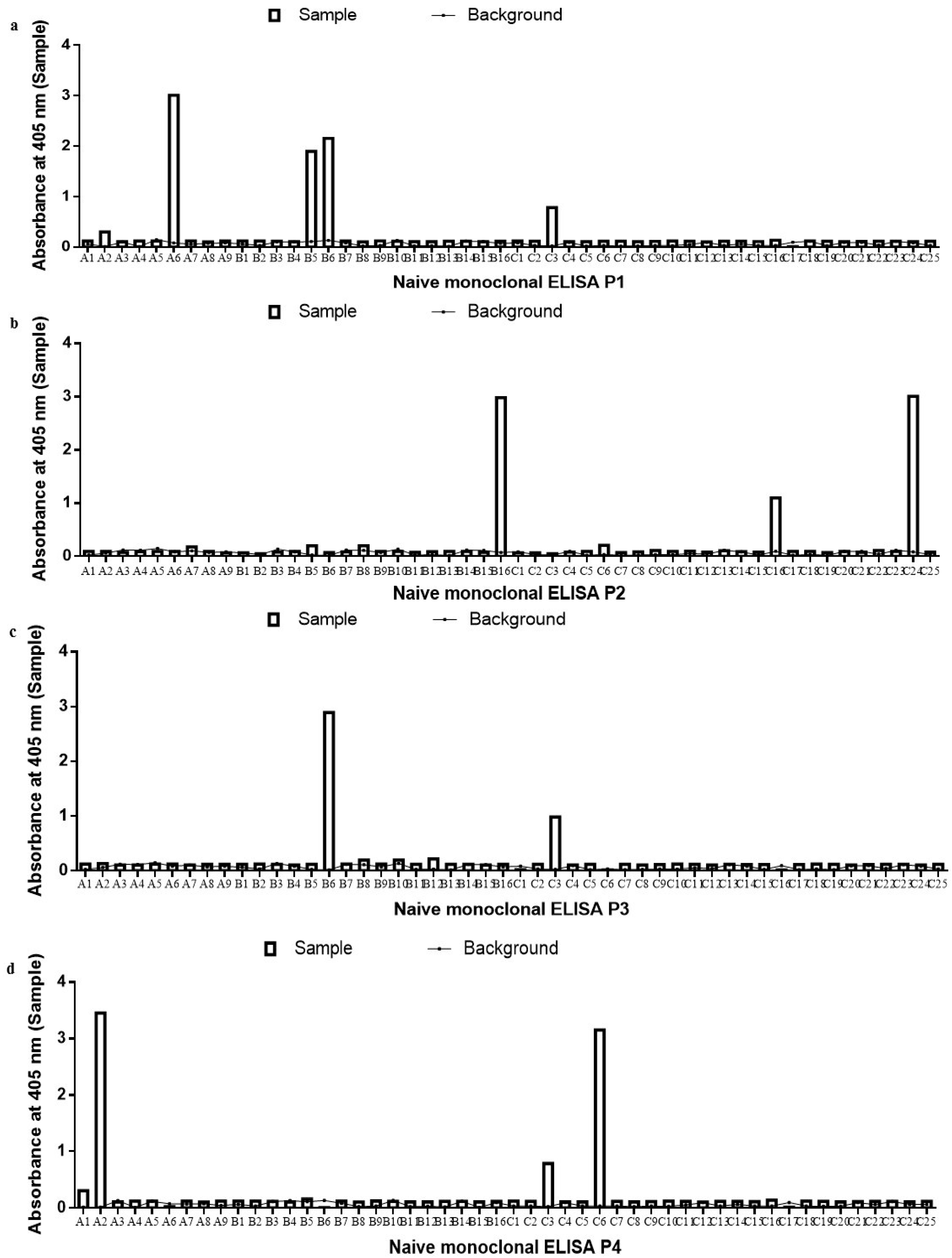
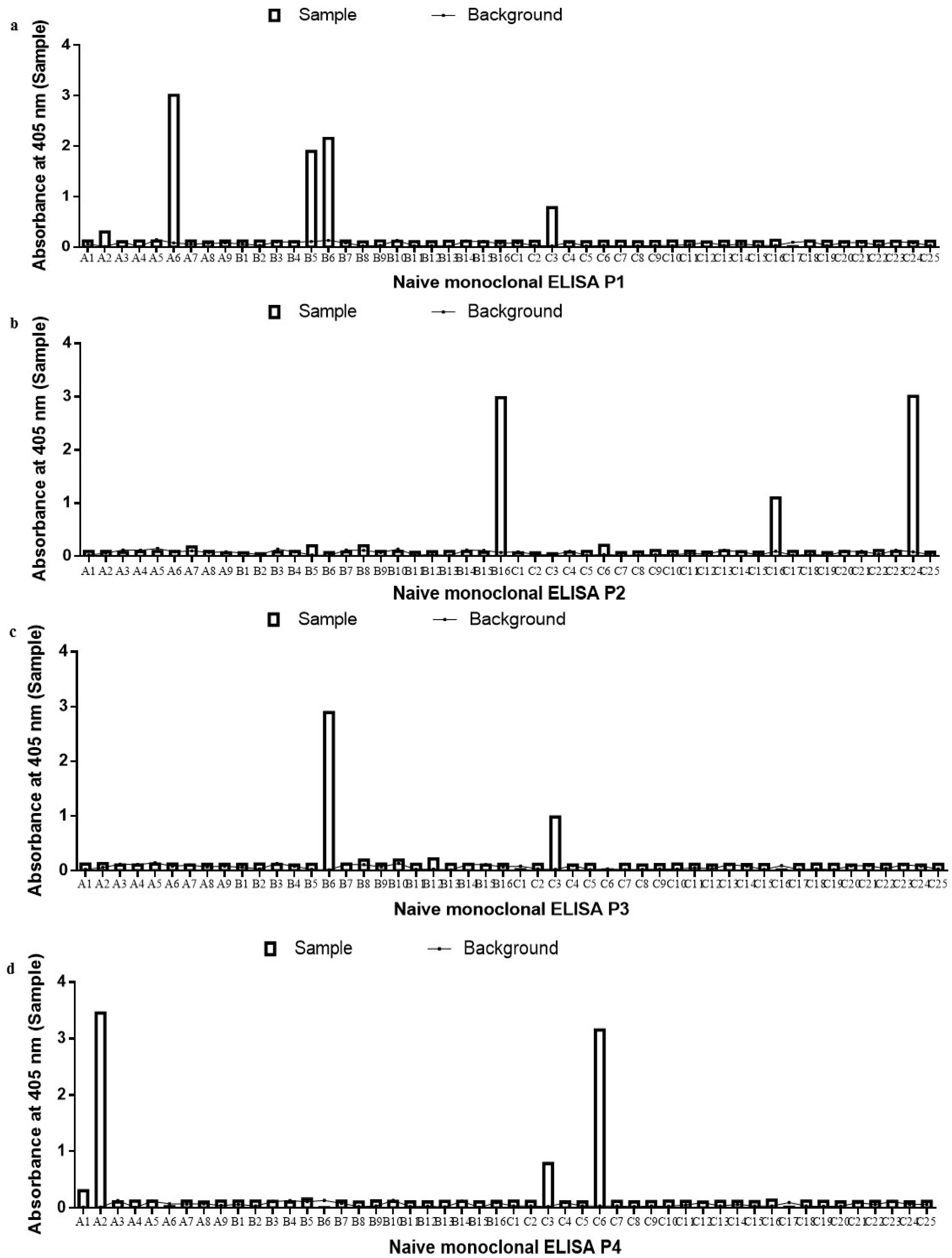
2.2. Monoclonal scFv ELISA
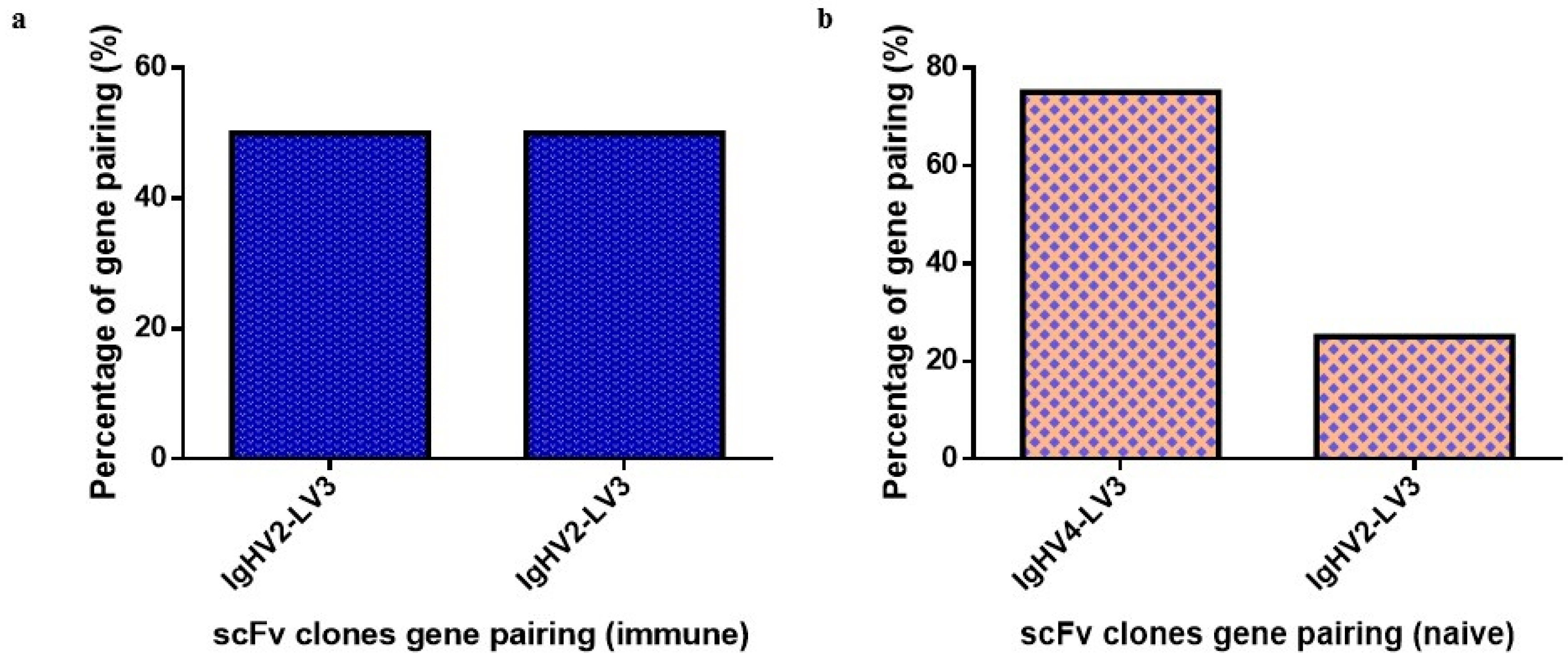
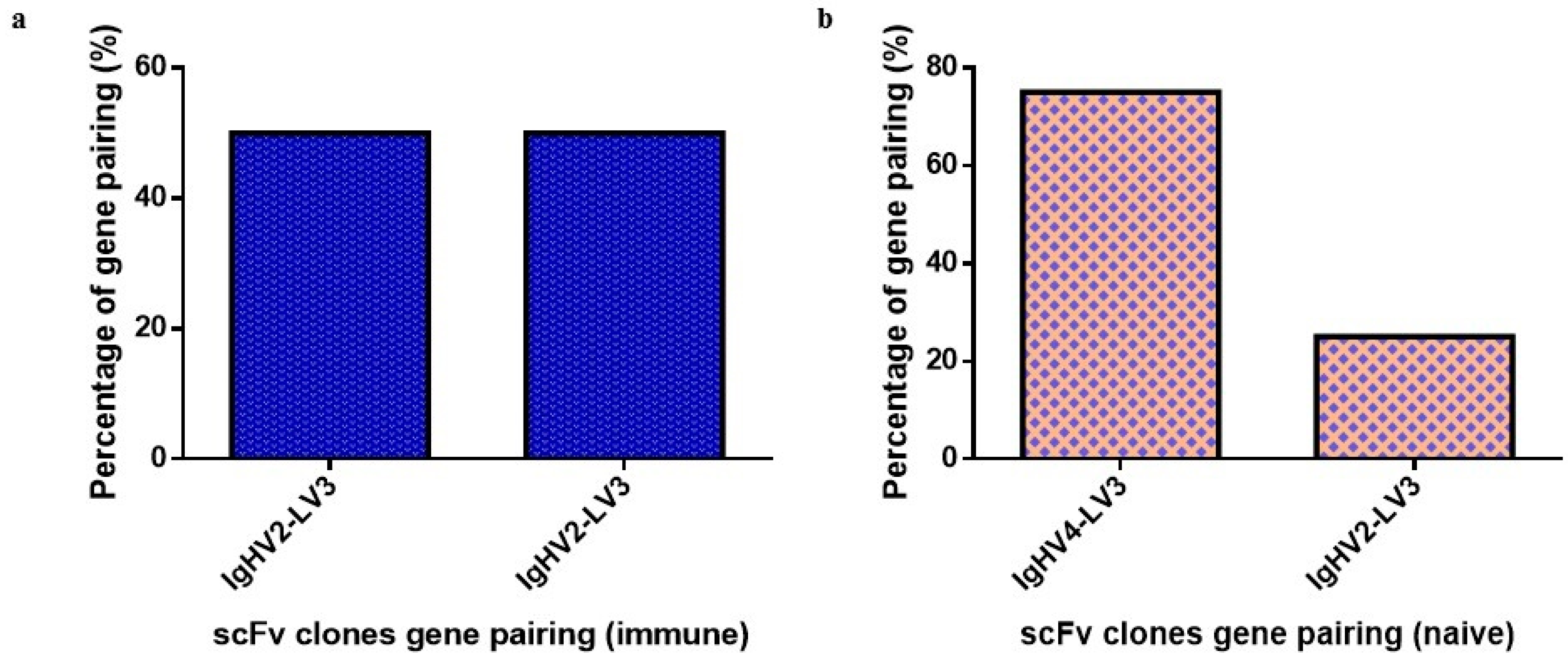
2.3. Antibody Sequence and Gene Analysis
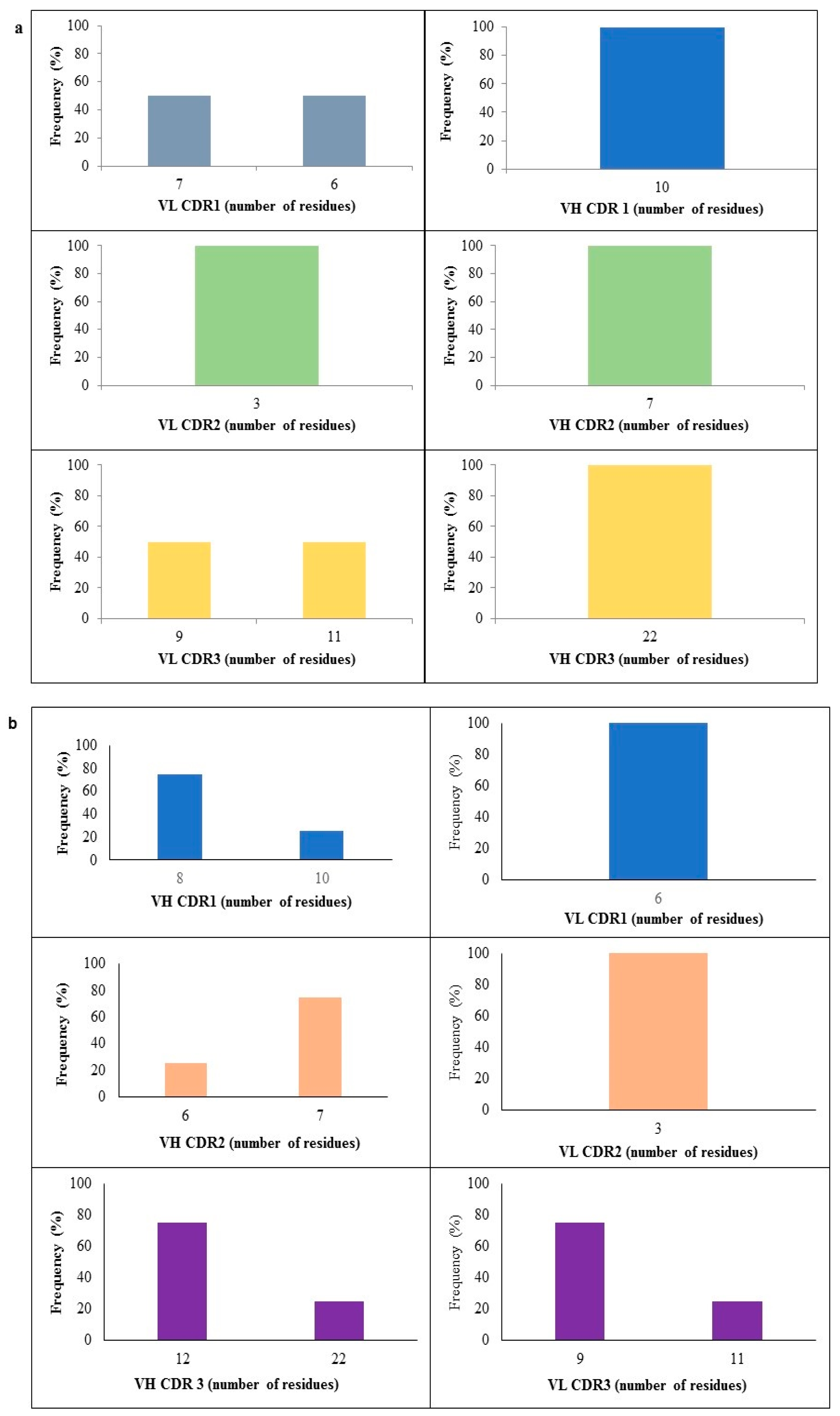
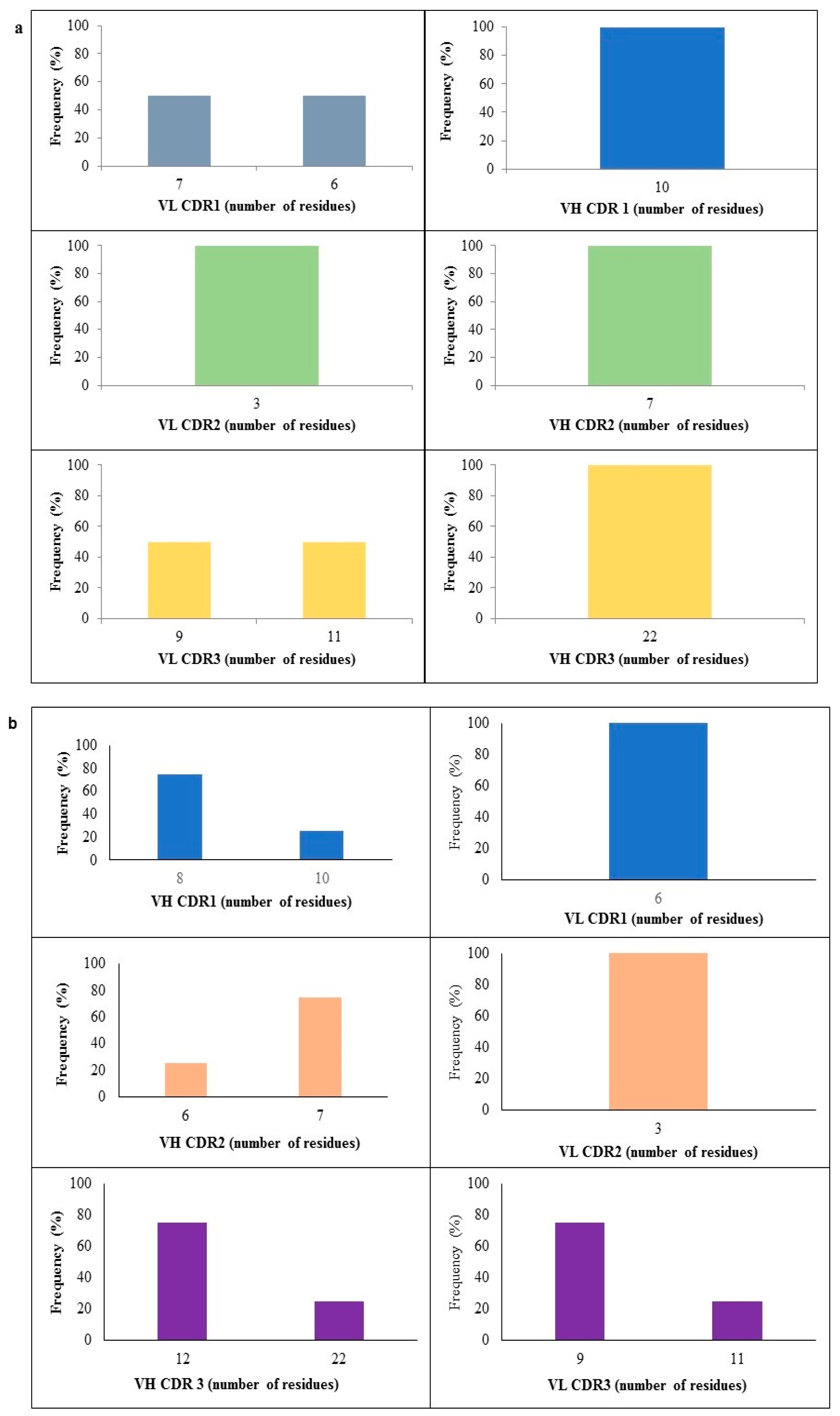
2.4. CDR Length Analysis
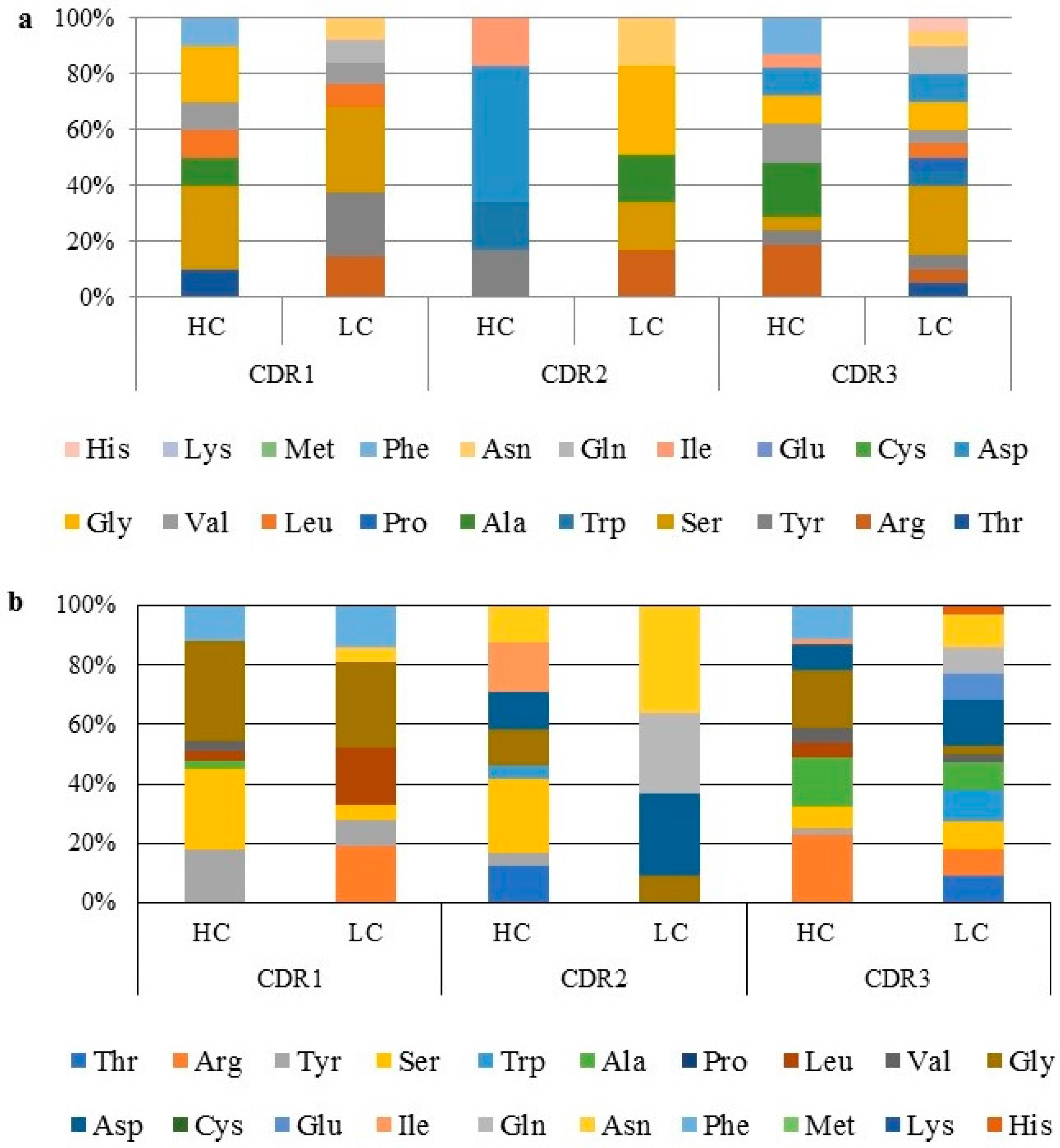
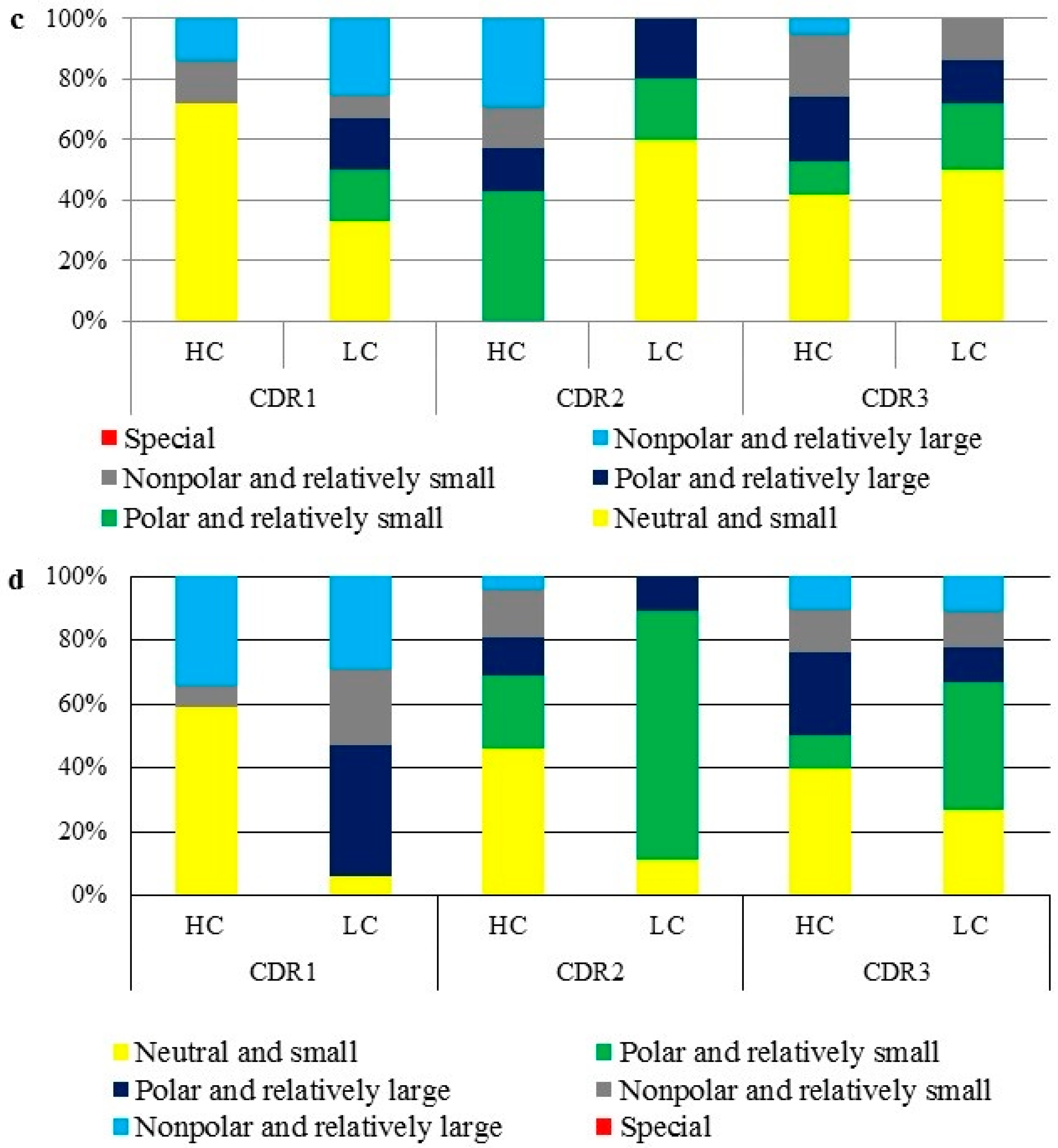
2.5. Amino Acid Distribution and Polarity Analysis
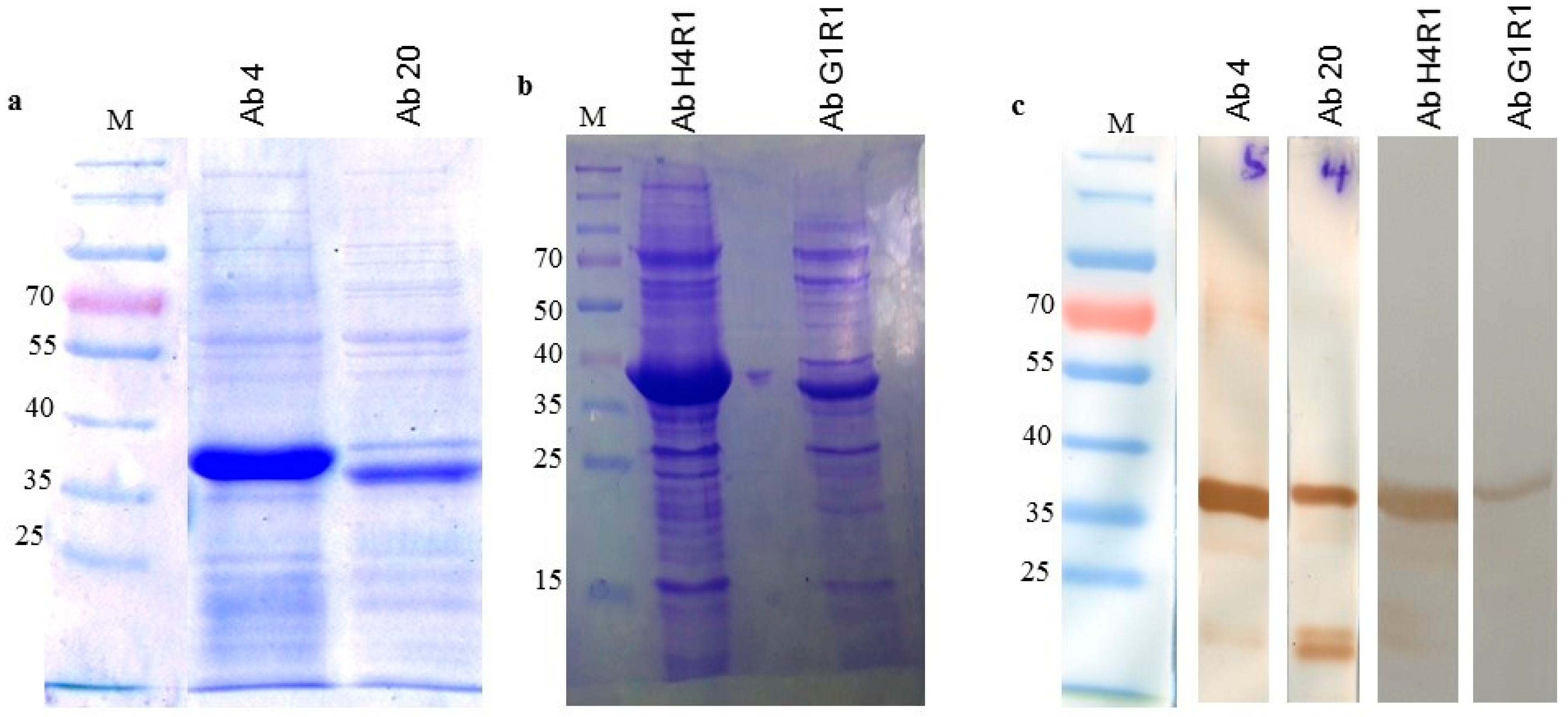
2.6. Soluble Expression of scFv Clones
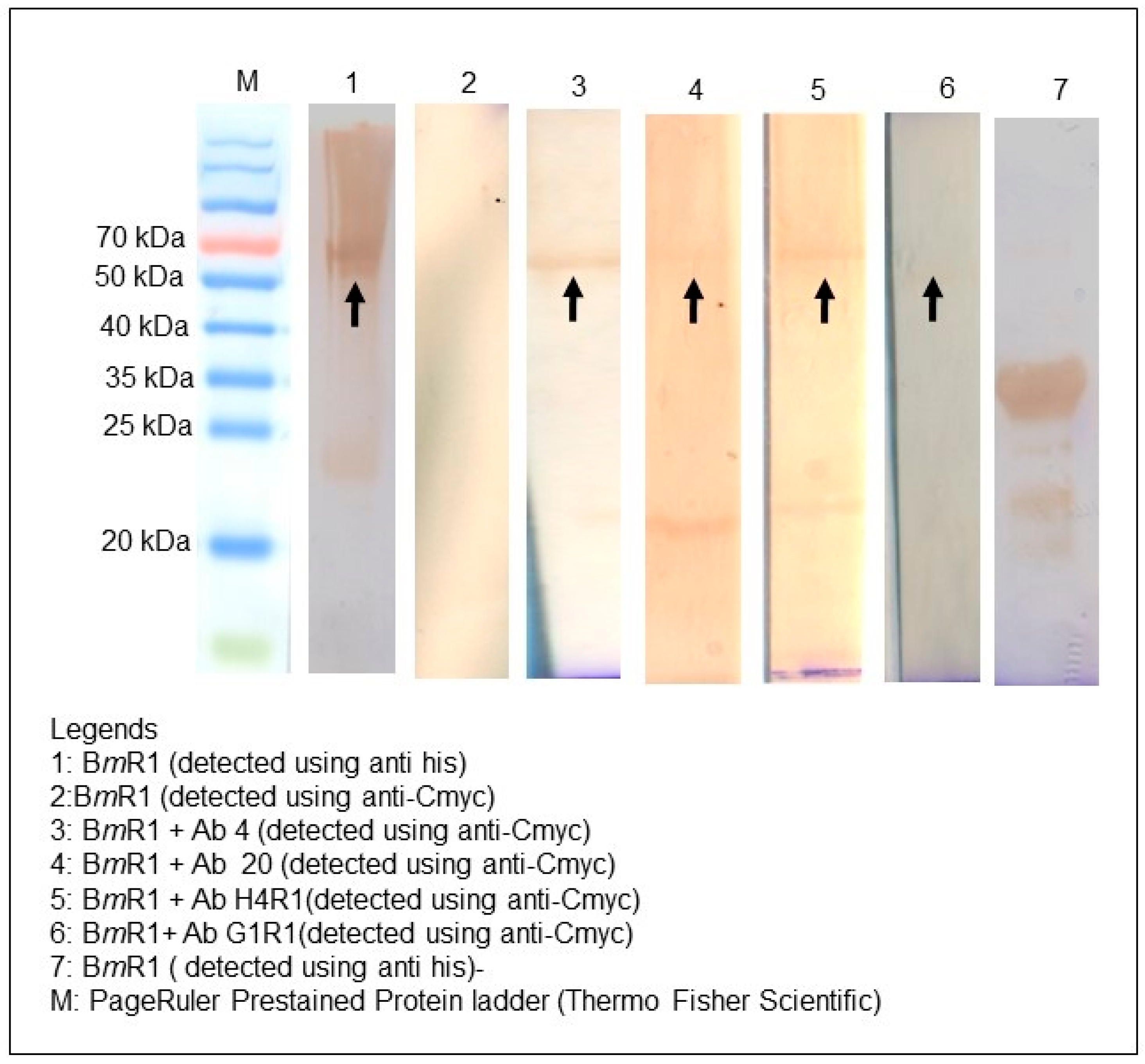
2.7. Binding Characterization
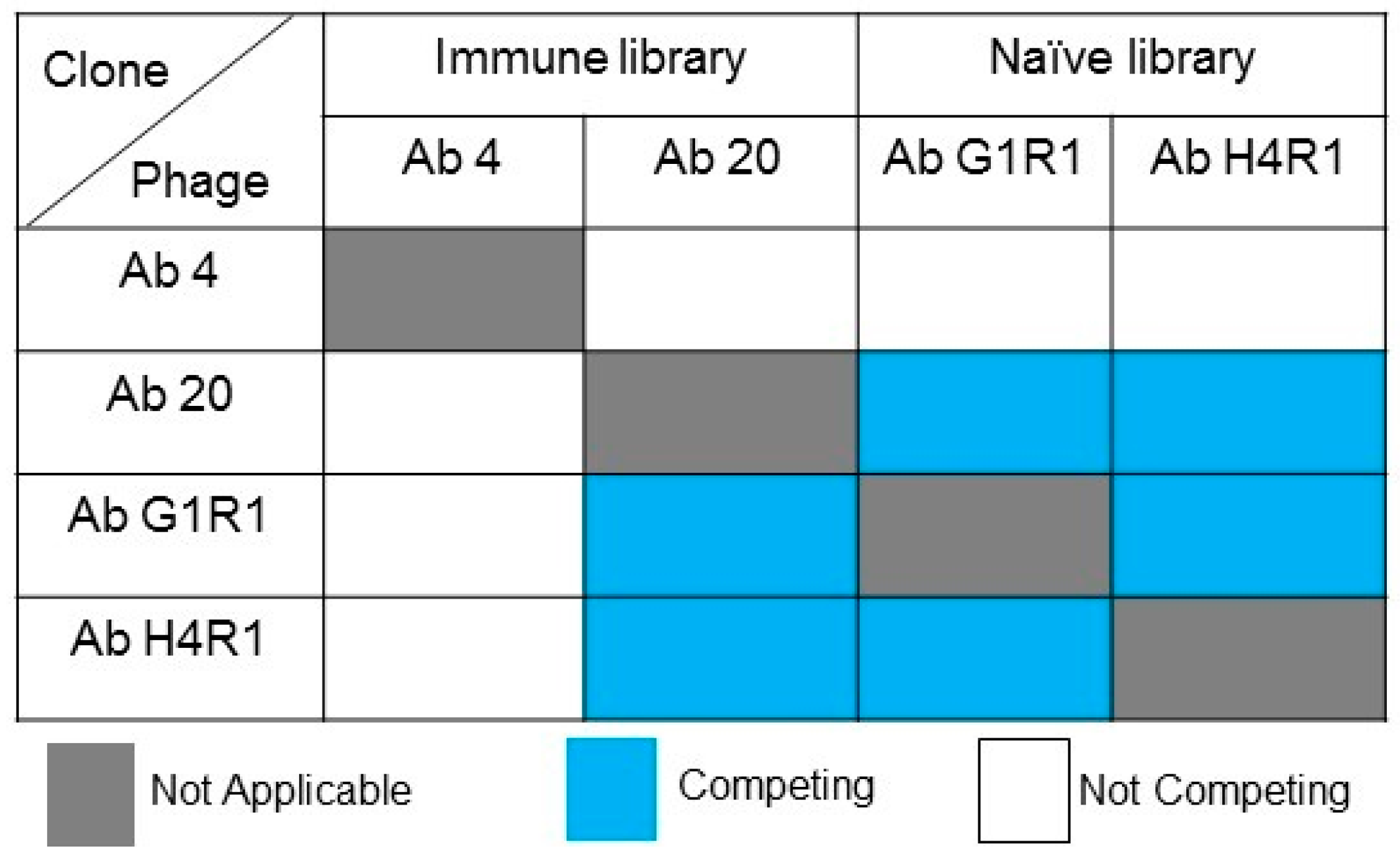
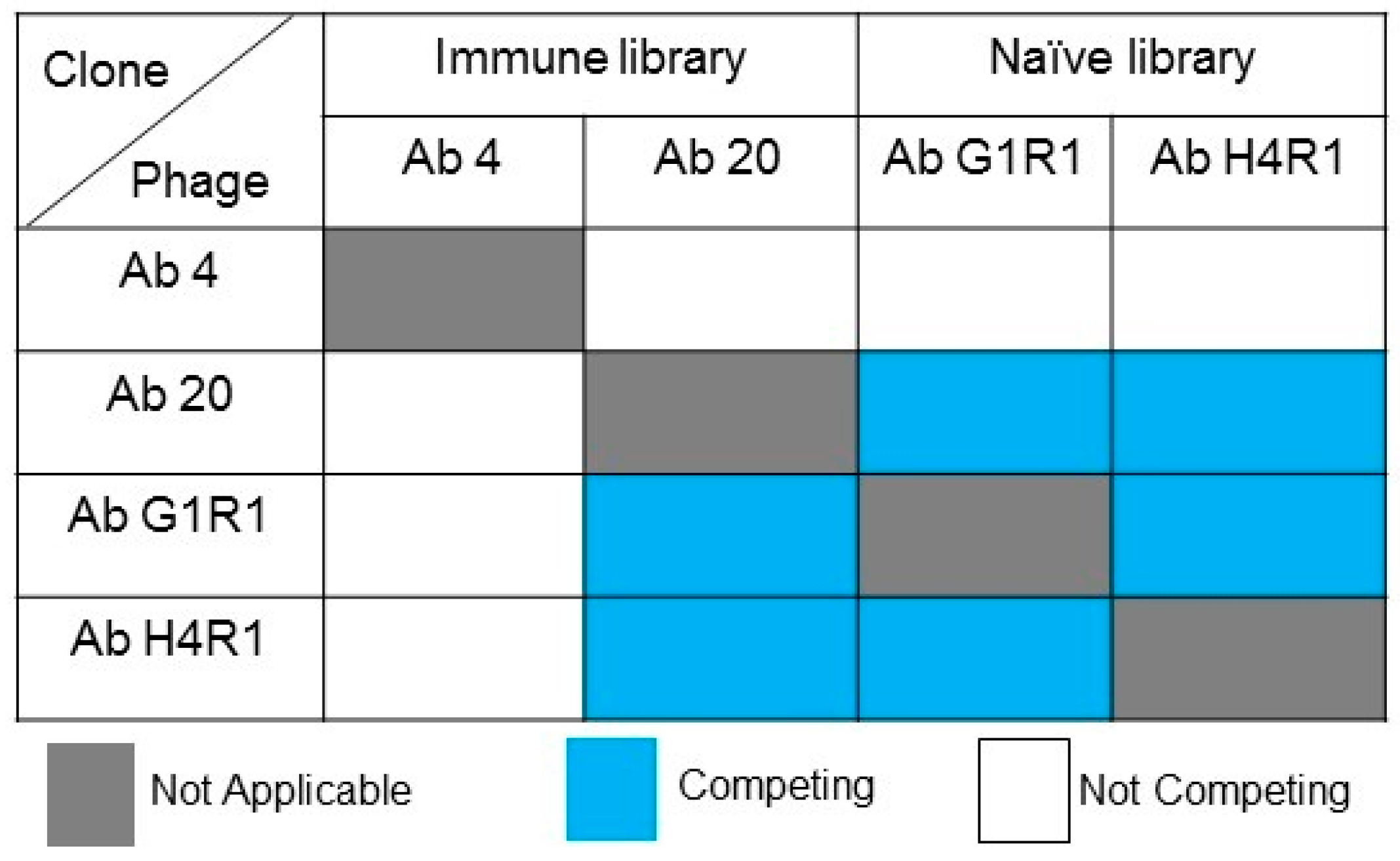
2.8. Competitive Analysis of Identified scFv Clones
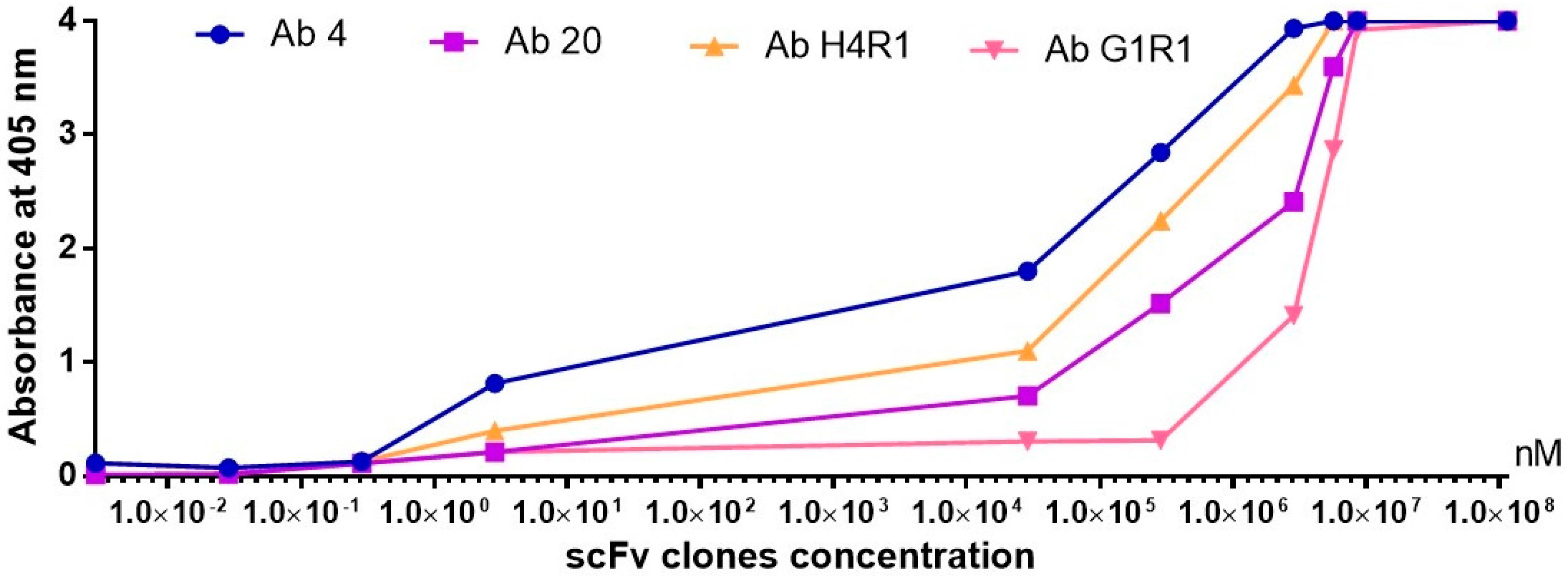
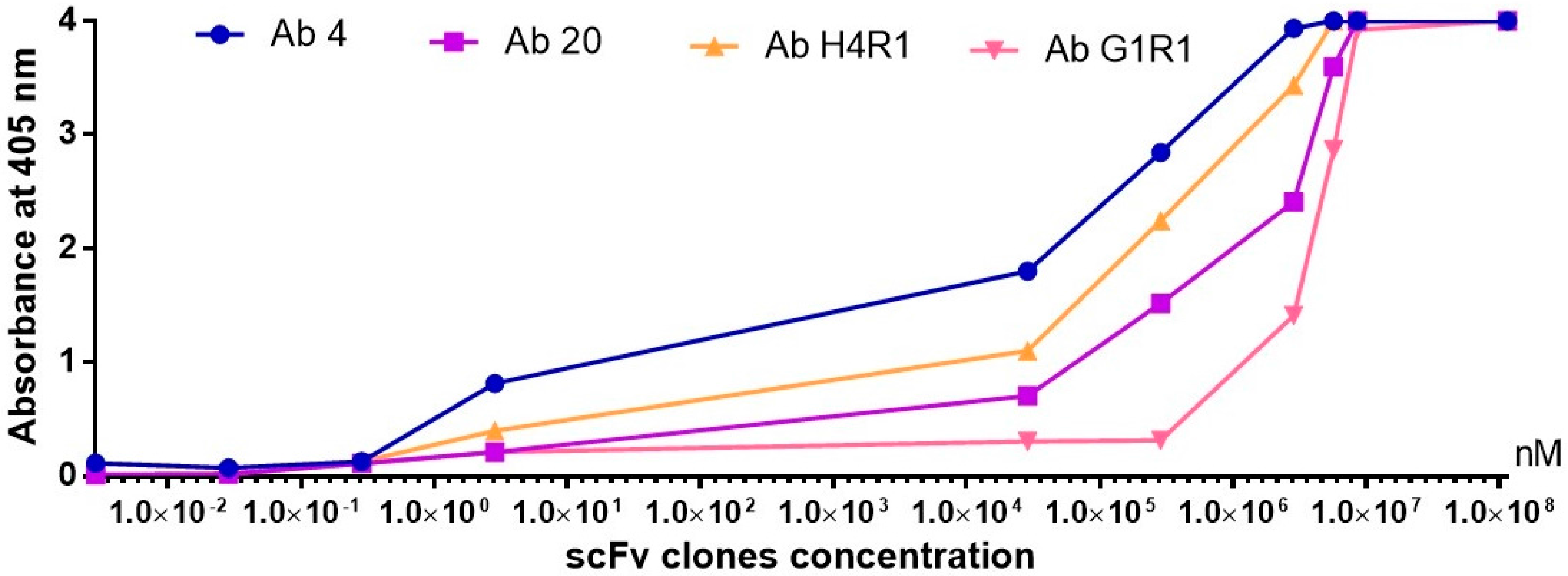
2.9. Antibody Titration ELISA
2.10. Immunoaffinity Pulldown Using Identified scFv
3. Discussion
4. Materials and Methods
4.1. Human scFv Library
4.2. Biopanning
4.3. Polyclonal and Monoclonal ELISA
4.4. Positive Clone Sequencing
4.5. Monoclonal Antibody Protein Expression
4.6. Purification of Monoclonal Antibody Protein
4.7. Western Blot Analysis
4.8. Competitive ELISA
4.9. Antibody Titration ELISA
4.10. Immunoaffinity Pulldown
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PCR | Polymerase chain reaction |
| Ig | Immunoglobulin |
| scFv | Single-chain variable fragments |
| mAb | Monoclonal antibody |
| CDR | Complementary determining region |
| HC | Heavy chain |
| LC | Light chain |
| CNBr | Cyanogen bromide |
References
- Maldjian, C.; Khanna, V.; Tandon, B.; Then, M.; Yassin, M.; Adam, R.; Klein, M.J. Lymphatic filariasis disseminating to the upper extremity. Case Rep. Radiol. 2014, 2014, 985680. [Google Scholar] [CrossRef] [PubMed]
- Bockarie, M.J.; Fischer, P.; Williams, S.A.; Zimmerman, P.A.; Griffin, L.; Alpers, M.P.; Kazura, J.W. Application of a polymerase chain reaction-elisa to detect Wuchereria bancrofti in pools of wild-caught anopheles punctulatus in a filariasis control area in papua new guinea. Am. J. Trop. Med. Hyg. 2000, 62, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Adjobimey, T.; Hoerauf, A. Induction of immunoglobulin G4 in human filariasis: An indicator of immunoregulation. Ann. Trop. Med. Parasitol. 2010, 104, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.M.; Rutitzky, L.I.; Urban, J.F., Jr.; Stadecker, M.J.; Gause, W.C. Protective immune mechanisms in helminth infection. Nat. Rev. Immunol. 2007, 7, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr. How the immune system protects the host from infection. Microbes Infect. 2001, 3, 1167–1171. [Google Scholar] [CrossRef]
- Hammers, C.M.; Stanley, J.R. Antibody phage display: Technique and applications. J. Investig. Dermatol. 2014, 134, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hust, M.; Frenzel, A.; Meyer, T.; Schirrmann, T.; Dubel, S. Construction of human naive antibody gene libraries. Methods Mol. Biol. 2012, 907, 85–107. [Google Scholar] [PubMed]
- Harris, N.; Gause, W.C. To B or not to B: B cells and the Th2-type immune response to helminths. Trends Immunol. 2011, 32, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Rohatgi, S.; Dutta, D.; Tahir, S.; Sehgal, D. Molecular dissection of antibody responses against pneumococcal surface protein A: Evidence for diverse D-H-less heavy chain gene usage and avidity maturation. J. Immunol. 2009, 182, 5570–5585. [Google Scholar] [CrossRef] [PubMed]
- Scheid, J.F.; Mouquet, H.; Feldhahn, N.; Seaman, M.S.; Velinzon, K.; Pietzsch, J.; Ott, R.G.; Anthony, R.M.; Zebroski, H.; Hurley, A.; et al. Broad diversity of neutralizing antibodies isolated from memory B cells in HIV-infected individuals. Nature 2009, 458, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Binley, J.M.; Lybarger, E.A.; Crooks, E.T.; Seaman, M.S.; Gray, E.; Davis, K.L.; Decker, J.M.; Wycuff, D.; Harris, L.; Hawkins, N.; et al. Profiling the specificity of neutralizing antibodies in a large panel of plasmas from patients chronically infected with human immunodeficiency virus type 1 subtypes B and C. J. Virol. 2008, 82, 11651–11668. [Google Scholar] [CrossRef] [PubMed]
- Throsby, M.; van den Brink, E.; Jongeneelen, M.; Poon, L.L.M.; Alard, P.; Cornelissen, L.; Bakker, A.; Cox, F.; van Deventer, E.; Guan, Y.; et al. Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1 and H1N1 recovered from human IgM(+) memory B cells. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahumatullah, A.; Ahmad, A.; Noordin, R.; Lim, T.S. Delineation of BmSXP antibody V-gene usage from a lymphatic filariasis based immune scFV antibody library. Mol. Immunol. 2015, 67, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.N.; Chin, C.F.; Choong, Y.S.; Ismail, A.; Lim, T.S. Generation of a naive human single chain variable fragment (scFV) library for the identification of monoclonal scFV against Salmonella Typhi Hemolysin E antigen. Toxicon 2016, 117, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Venet, S.; Kosco-Vilbois, M.; Fischer, N. Comparing CDRH3 diversity captured from secondary lymphoid organs for the generation of recombinant human antibodies. mAbs 2013, 5, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Iorno, N.; Sierro, F.; Christ, D. Selection of human antibody fragments by phage display. Nat. Protoc. 2007, 2, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.J.; Deng, S.J.; Huang, D.G.; He, Y.; Lei, M.; Zhou, L.; Jin, P. Frontier of therapeutic antibody discovery: The challenges and how to face them. World J. Biol. Chem. 2012, 3, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Watkins, N.A.; Ouwehand, W.H. Introduction to antibody engineering and phage display. Vox Sang. 2000, 78, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.; Hamid, M. scFV antibody: Principles and clinical application. Clin. Dev. Immunol. 2012, 2012, 980250. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.N.; Tye, G.J.; Choong, Y.S.; Ong, E.B.; Ismail, A.; Lim, T.S. Principles and application of antibody libraries for infectious diseases. Biotechnol. Lett. 2014, 36, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.A.; Ki, M.K.; Lee, S.; Hong, M.L.; Kim, M.; Kim, S.; Chung, J.; Rhee, S.G.; Shim, H. Antibodies against non-immunizing antigens derived from a large immune scFV library. Mol. Cells 2011, 31, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Loset, G.A.; Lobersli, I.; Kavlie, A.; Stacy, J.E.; Borgen, T.; Kausmally, L.; Hvattum, E.; Simonsen, B.; Hovda, M.B.; Brekke, O.H. Construction, evaluation and refinement of a large human antibody phage library based on the IgD and IgM variable gene repertoire. J. Immunol. Methods 2005, 299, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Hust, M.; Meyer, T.; Voedisch, B.; Rulker, T.; Thie, H.; El-Ghezal, A.; Kirsch, M.I.; Schutte, M.; Helmsing, S.; Meier, D.; et al. A human scFV antibody generation pipeline for proteome research. J. Biotechnol. 2011, 152, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Grippo, V.; Mahler, E.; Elias, F.E.; Cauerhff, A.; Gomez, K.A.; Tentori, M.C.; Ruiz, A.; Vigliano, C.A.; Laguens, R.P.; Berek, C.; et al. The heavy chain variable segment gene repertoire in chronic chagas’ heart disease. J. Immunol. 2009, 183, 8015–8025. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Watanabe, K.; Cheng, X.J.; Tsukamoto, H.; Kaneda, Y.; Takeuchi, T.; Ihara, S.; Petri, W.A., Jr. VH3 gene usage in neutralizing human antibodies specific for the Entamoeba histolytica Gal/GalNAc lectin heavy subunit. Infect. Immun. 2003, 71, 4313–4319. [Google Scholar] [CrossRef] [PubMed]
- Babor, M.; Kortemme, T. Multi-constraint computational design suggests that native sequences of germline antibody H3 loops are nearly optimal for conformational flexibility. Proteins Struct. Funct. Bioinform. 2009, 75, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Chothia, C.; Lesk, A.M.; Tramontano, A.; Levitt, M.; Smithgill, S.J.; Air, G.; Sheriff, S.; Padlan, E.A.; Davies, D.; Tulip, W.R.; et al. Conformations of immunoglobulin hypervariable regions. Nature 1989, 342, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.T.; Dear, P.H.; Foote, J.; Neuberger, M.S.; Winter, G. Replacing the complementarity-determining regions in a human-antibody with those from a mouse. Nature 1986, 321, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Kunik, V.; Ofran, Y. The indistinguishability of epitopes from protein surface is explained by the distinct binding preferences of each of the six antigen-binding loops. Protein Eng. Des. Sel. 2013, 26, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Fellouse, F.A.; Barthelemy, P.A.; Kelley, R.F.; Sidhu, S.S. Tyrosine plays a dominant functional role in the paratope of a synthetic antibody derived from a four amino acid code. J. Mol. Biol. 2006, 357, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Zemlin, M.; Klinger, M.; Link, J.; Zemlin, C.; Bauers, K.; Engler, J.A.; Schroeder, H.W.; Kirkham, P.M. Expressed murine and human CDR-H3 intervals of equal length exhibit distinct repertoires that differ in their amino acid composition and predicted range of structures. J. Mol. Biol. 2003, 334, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Birtalan, S.; Fisher, R.D.; Sidhu, S.S. The functional capacity of the natural amino acids for molecular recognition. Mol. Biosyst. 2010, 6, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, D.; Rahat, O.; Cohen, M.; Neuvirth, H.; Schreiber, G. The molecular architecture of protein-protein binding sites. Curr. Opin. Struct. Biol. 2007, 17, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Villar, H.O.; Kauvar, L.M. Amino-acid preferences at protein-binding sites. FEBS Lett. 1994, 349, 125–130. [Google Scholar] [CrossRef]
- Birtalan, S.; Zhang, Y.N.; Fellouse, F.A.; Shao, L.H.; Schaefer, G.; Sidhu, S.S. The intrinsic contributions of tyrosine, serine, glycine and arginine to the affinity and specificity of antibodies. J. Mol. Biol. 2008, 377, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.R. Bioinformatics for Geneticists: A Bioinformatics Primer for the Analysis of Genetic Data, 2nd ed.; Wiley: Chichester, UK; Hoboken, NJ, USA, 2007; p. xxii. 554p. [Google Scholar]
- Domina, M.; Cariccio, V.L.; Benfatto, S.; Venza, M.; Venza, I.; Borgogni, E.; Castellino, F.; Midiri, A.; Galbo, R.; Romeo, L.; et al. Functional characterization of a monoclonal antibody epitope using a lambda phage display-deep sequencing platform. Sci. Rep. 2016, 6, 31458. [Google Scholar] [CrossRef] [PubMed]
- Levenhagen, M.A.; Santos, F.D.A.; Fujimura, P.T.; Caneiro, A.P.; Costa-Cruz, J.M.; Goulart, L.R. Structural and functional characterization of a novel scFV anti-HSP60 of Strongyloides sp. Sci. Rep. 2015, 5, 10447. [Google Scholar] [CrossRef] [PubMed]
- Sela-Culang, I.; Kunik, V.; Ofran, Y. The structural basis of antibody-antigen recognition. Front. Immunol. 2013, 4, 302. [Google Scholar] [CrossRef] [PubMed]
- Lescar, J.; Pellegrini, M.; Souchon, H.; Tello, D.; Poljak, R.J.; Peterson, N.; Greene, M.; Alzari, P.M. Crystal-structure of a cross-reaction complex between Fab F9.13.7 and guinea-fowl lysozyme. J. Biol. Chem. 1995, 270, 18067–18076. [Google Scholar] [CrossRef] [PubMed]
- Grellier, B.; Le Pogam, F.; Vitorino, M.; Starck, J.P.; Geist, M.; Duong, V.; Haegel, H.; Menguy, T.; Bonnefoy, J.Y.; Marchand, J.B.; et al. 3D modeling and characterization of the human CD115 monoclonal antibody H27K15 epitope and design of a chimeric CD115 target. MAbS 2014, 6, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Vangone, A.; Abdel-Azeim, S.; Caputo, I.; Sblattero, D.; Di Niro, R.; Cavallo, L.; Oliva, R. Structural basis for the recognition in an idiotype-anti-idiotype antibody complex related to celiac disease. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olimpieri, P.P.; Chailyan, A.; Tramontano, A.; Marcatili, P. Prediction of site-specific interactions in antibody-antigen complexes: The proabc method and server. Bioinformatics 2013, 29, 2285–2291. [Google Scholar] [CrossRef] [PubMed]
- Frenzel, A.; Schirrmann, T.; Hust, M. Phage display-derived human antibodies in clinical development and therapy. MAbS 2016, 8, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Norsyahida, A.; Rahmah, N.; Ahmad, R.M.Y. Effects of feeding and induction strategy on the production of BmR1 antigen in recombinant E-coli. Lett. Appl. Microbiol. 2009, 49, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, M.P.; Giudicelli, V.; Ginestoux, C.; Jabado-Michaloud, J.; Folch, G.; Bellahcene, F.; Wu, Y.; Gemrot, E.; Brochet, X.; Lane, J.; et al. IMGT®, the international ImMunoGeneTics information system®. Nucleic Acids Res. 2009, 37, D1006–D1012. [Google Scholar] [CrossRef] [PubMed]
- Retter, I.; Althaus, H.H.; Munch, R.; Muller, W. VBASE2, an integrative V gene database. Nucleic Acids Res. 2005, 33, D671–D674. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.F.; Chan, S.K.; Tye, G.J.; Choong, Y.S.; Lim, T.S. Improved expression of single-chain fragment variable antibodies devoid of leader peptides in the cytoplasm. Curr. Proteom. 2015, 12, 117–123. [Google Scholar] [CrossRef]
- Kavran, J.M.; Leahy, D.J. Coupling antibody to cyanogen bromide-activated sepharose. In Laboratory Methods in Enzymology: Protein, Pt C; Academic Press: Cambridge, MA, USA, 2014; Volume 541, pp. 27–34. [Google Scholar]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahumatullah, A.; Abdul Karim, I.Z.; Noordin, R.; Lim, T.S. Antibody-Based Protective Immunity against Helminth Infections: Antibody Phage Display Derived Antibodies against BmR1 Antigen. Int. J. Mol. Sci. 2017, 18, 2376. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112376
Rahumatullah A, Abdul Karim IZ, Noordin R, Lim TS. Antibody-Based Protective Immunity against Helminth Infections: Antibody Phage Display Derived Antibodies against BmR1 Antigen. International Journal of Molecular Sciences. 2017; 18(11):2376. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112376
Chicago/Turabian StyleRahumatullah, Anizah, Izzati Zahidah Abdul Karim, Rahmah Noordin, and Theam Soon Lim. 2017. "Antibody-Based Protective Immunity against Helminth Infections: Antibody Phage Display Derived Antibodies against BmR1 Antigen" International Journal of Molecular Sciences 18, no. 11: 2376. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112376