Characterization of the Asiatic Acid Glucosyltransferase, UGT73AH1, Involved in Asiaticoside Biosynthesis in Centella asiatica (L.) Urban



Abstract
:1. Introduction
2. Results and Discussion
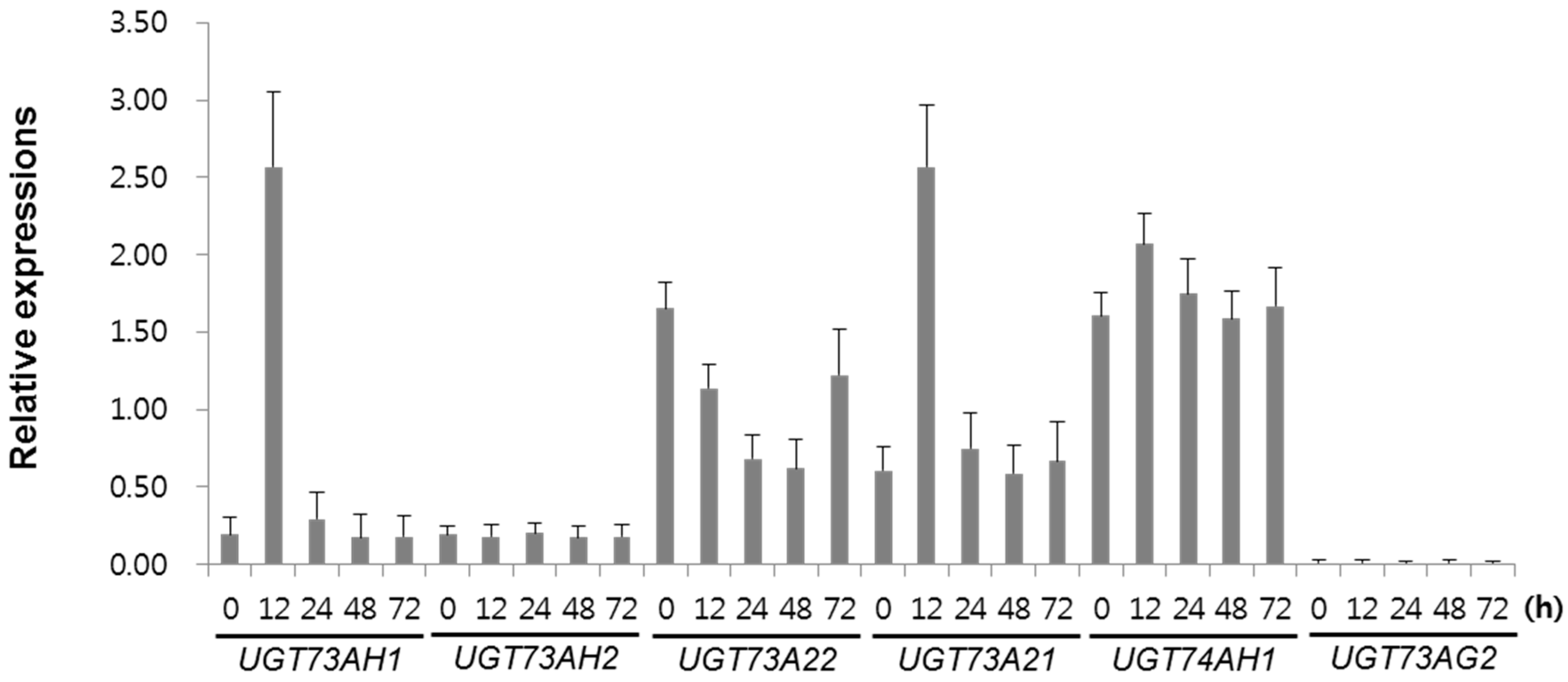
2.1. Transcriptome Analysis of Leaves Elicited with MeJA
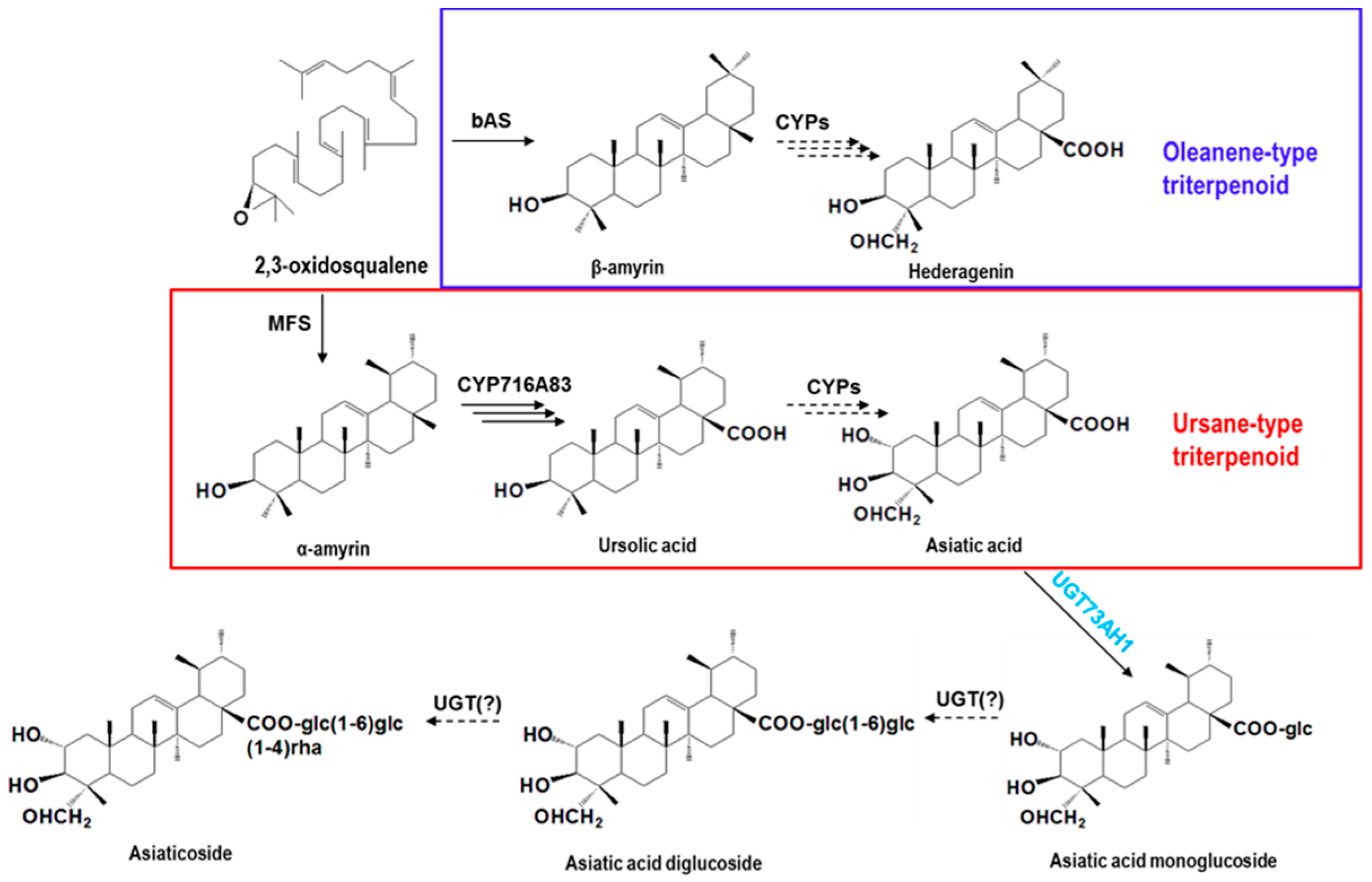
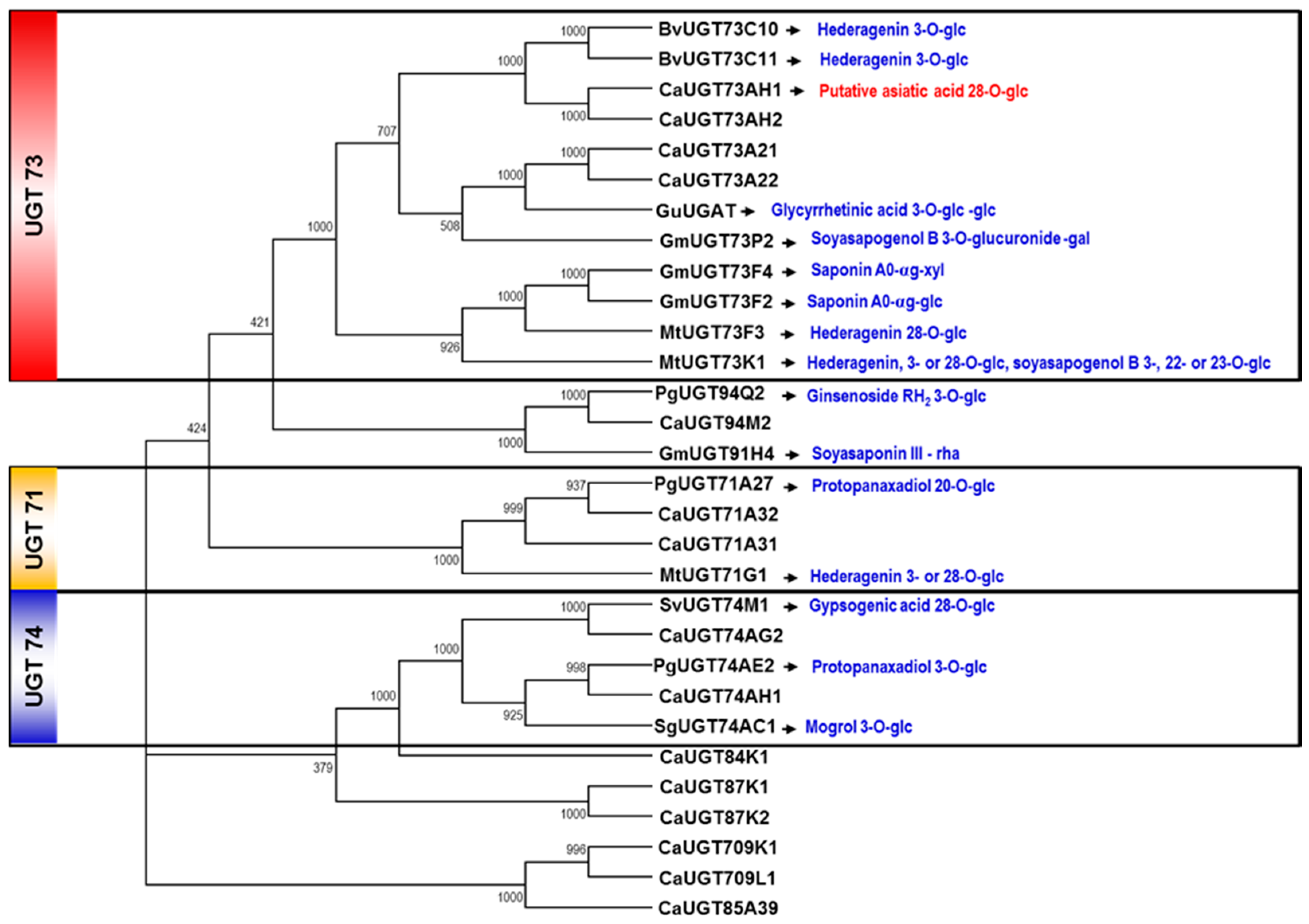
2.2. Selection of Candidate UGT Genes for Asiaticoside Biosynthesis
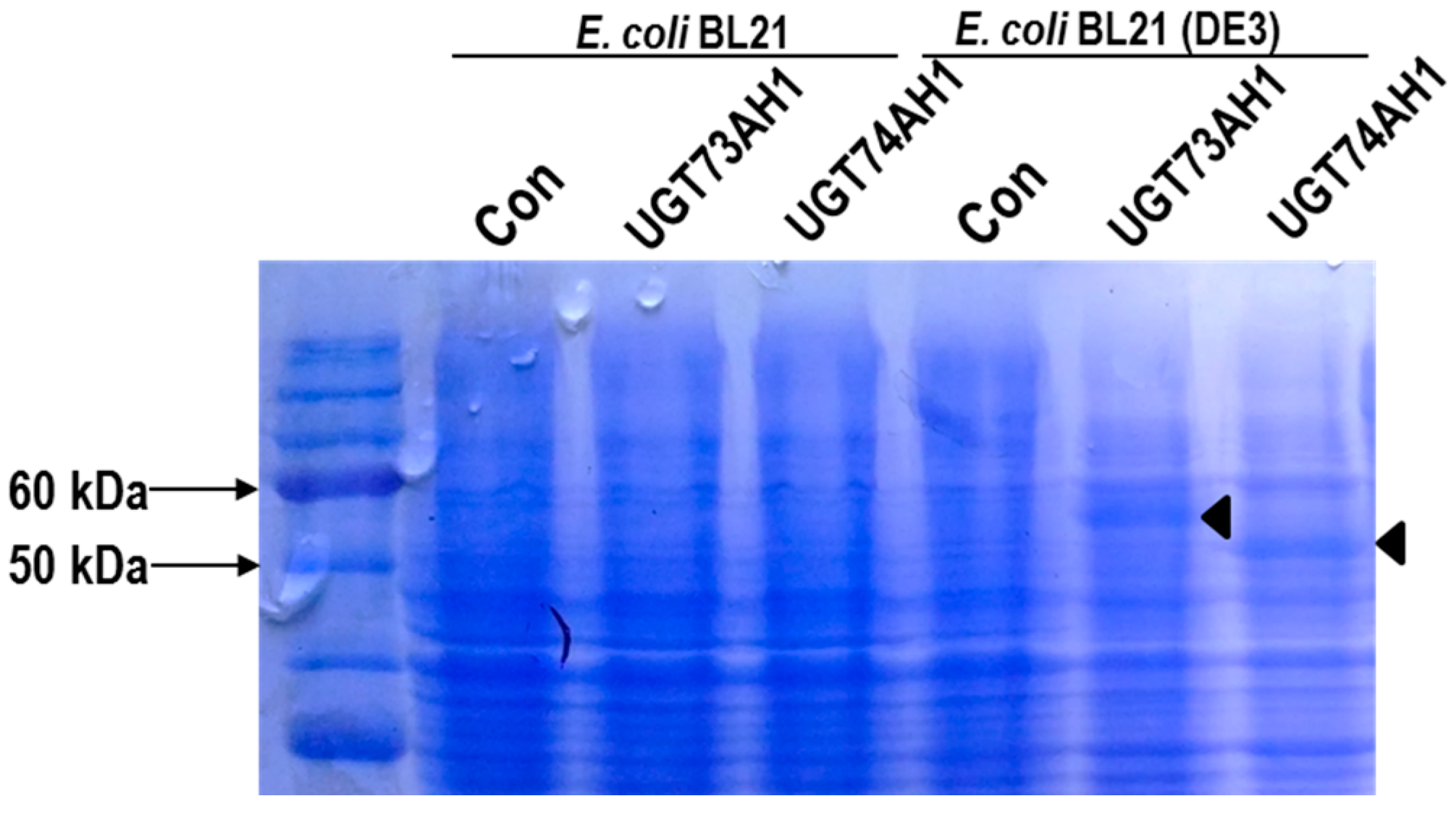
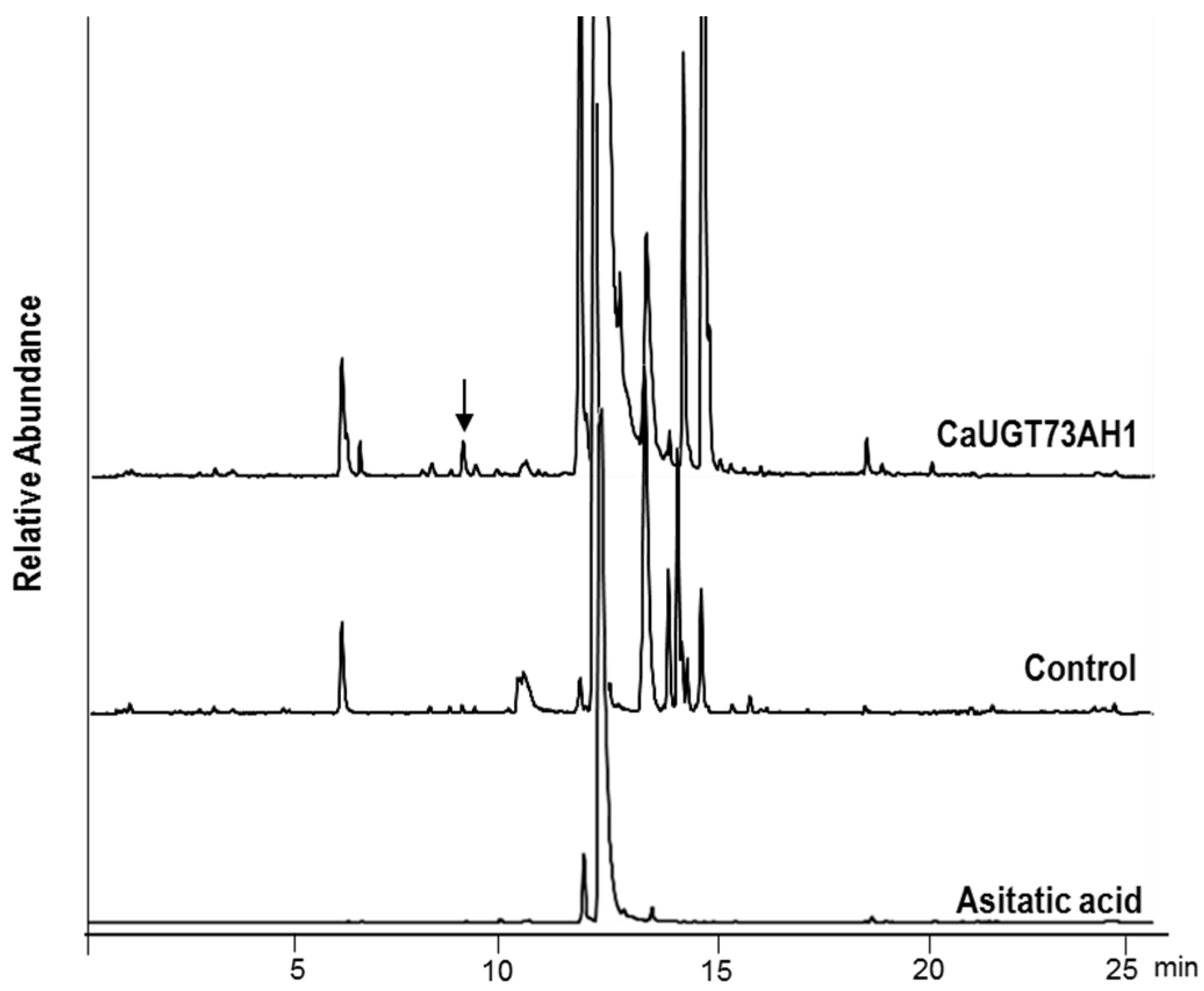
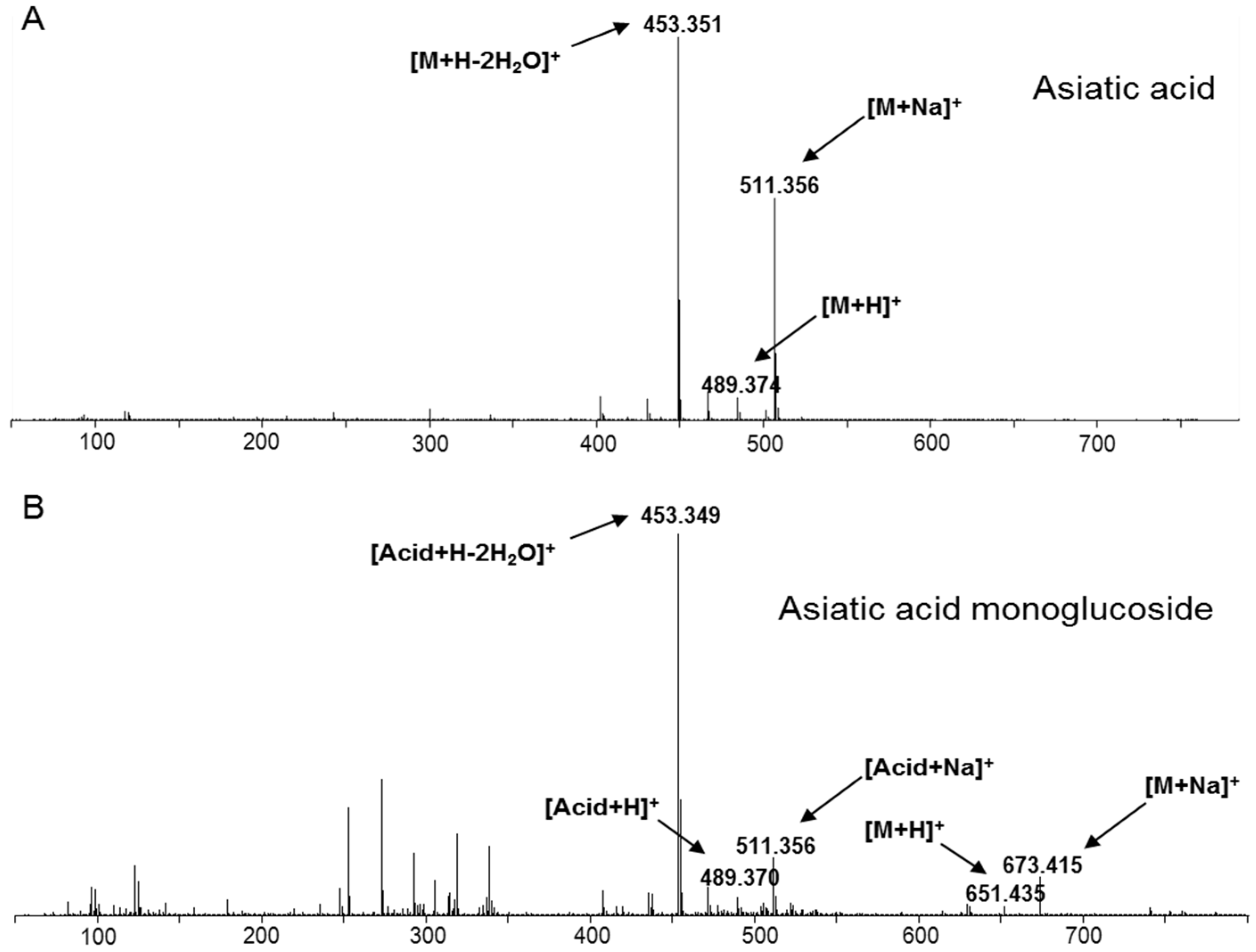
2.3. Functional Characterization of Candidate UGTs
3. Materials and Methods
3.1. Plant Material and Sampling
3.2. EST Assembly and Annotation
3.3. Expression Levels of UGT Genes in C. asiatica
3.4. Phylogenetic Tree Analysis
3.5. Real-Time RT-PCR Analysis
3.6. Expression of UGT Genes
3.7. UPLC-QTOF/MS Analysis of the Products from Enzymatic Reactions
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| UGT | UDP-glucosyltransferase |
| MeJA | Methyl jasmonate |
| RPKM | Reads per kilobase per million mapped reads |
| EST | Expressed sequence tag |
| UPLC-QTOP/MS | Ultra-high Performance Liquid Chromatography-Quadrupole Time-of-Flight Mass Spectrometry |
References
- Jones, P.; Vogt, T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta 2001, 213, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T.; Jones, P. Glycosyltransferases in plant natural product synthesis: Characterization of a supergene family. Trends Plant Sci. 2000, 5, 380–386. [Google Scholar] [CrossRef]
- Phillips, D.R.; Rasbery, J.M.; Bartel, B.; Matsuda, S.P.T. Biosynthetic diversity in plant triterpene cyclization. Curr. Opin. Plant Biol. 2006, 9, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Seki, H.; Tamura, K.; Muranaka, T. P450s and UGTs: Key players in the structural diversity of triterpenoid saponins. Plant Cell Physiol. 2015, 56, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Fazio, G.C.; Matsuda, S.P.T. On the origins of triterpenoid skeletal diversity. Phytochemistry 2014, 65, 261–291. [Google Scholar] [CrossRef]
- Ross, J.; Li, Y.; Lim, E.K.; Bowles, D.J. Higher plant glycosyltransferases. Genome Biol. 2001, 2, 3004.1–3004.6. [Google Scholar] [CrossRef]
- Li, Y.; Baldauf, S.; Lim, E.K.; Bowles, D.J. Phylogenetic analysis of the UDP-glycosyltransferase multigene family of Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 4338–4343. [Google Scholar] [CrossRef] [PubMed]
- Achnine, L.; Huhman, D.V.; Farag, M.A.; Sumner, L.W.; Blount, J.W.; Dixon, R.A. Genomics-based selection and functional characterization of triterpene glycosyltransferases from the model legume Medicago truncatula. Plant J. 2005, 41, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Paquette, S.; Møller, B.L.; Bak, S. On the origin of family 1 plant glycosyltransferases. Phytochemistry 2003, 62, 399–413. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Hanada, K. An evolutionary view of functional diversity in family 1 glycosyltransferases. Plant J. 2011, 66, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M.; Nishimura, K.; Yasuyama, N.; Ebizuka, Y. Identification and characterization of glycosyltransferases involved in the biosynthesis of soyasaponin I in Glycine max. FEBS Lett. 2010, 584, 2258–2264. [Google Scholar] [CrossRef] [PubMed]
- Owatworakit, A.; Townsend, B.; Louveau, T.; Jenner, H.; Rejzek, M.; Hughes, R.K.; Saalbach, G.; Qi, X.; Bakht, S.; Roy, A.D.; et al. Glycosyltransferases from oat (Avena) implicated in the acylation of avenacins. J. Biol. Chem. 2013, 288, 3696–3704. [Google Scholar] [CrossRef] [PubMed]
- Naoumkina, M.A.; Modolo, L.V.; Huhman, D.V.; Urbanczyk-Wochniak, E.; Tang, Y.; Sumner, L.W.; Dixon, R.A. Genomic and coexpression analyses predict multiple genes involved in triterpene saponin biosynthesis in Medicago truncatula. Plant Cell 2010, 22, 850–866. [Google Scholar] [CrossRef] [PubMed]
- Meesapyodsuk, D.; Balsevich, J.; Reed, D.W.; Covello, P.S. Saponin biosynthesis in Saponaria vaccaria. cDNAs encoding beta-amyrin synthase and a triterpene carboxylic acid glucosyltransferase. Plant Physiol. 2007, 143, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Augustin, J.M.; Drok, S.; Shinoda, T.; Sanmiya, K.; Nielsen, J.K.; Khakimov, B.; Olsen, C.E.; Hansen, E.H.; Kuzina, V.; Ekstrøm, C.T.; et al. UDP-glycosyltransferases from the UGT73C subfamily in Barbarea vulgaris catalyze sapogenin 3-O-glucosylation in saponin-mediated insect resistance. Plant Physiol. 2012, 160, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- De Costa, F.; Barber, C.J.S.; Kim, Y.B.; Reed, D.W.; Zhang, H.; Fett-Neto, A.; Covello, P.S. Molecular cloning of an ester-forming triterpenoid: UDP-glucose 28-O-glucosyltransferase involved in saponin biosynthesis from the medicinal plant Centella asiatica. Plant Sci. 2017, 262, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.T.; Kim, M.Y.; Hong, M.H.; Ahn, J.C.; Hwang, B. Stimulation of asiaticoside production from Centella asiatica whole plant cultures by elicitors. Plant Cell Rep. 2004, 23, 339–344. [Google Scholar] [CrossRef] [PubMed]
- James, J.T.; Dubery, I.A. Pentacyclic triterpenoids from the medicinal herb, Centella asiatica (L.) Urban. Molecules 2009, 14, 3922–3941. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Choi, J.Y.; Son, D.J.; Park, E.K.; Song, M.J.; Hellström, M.; Hong, J.T. Anti-inflammatory effect of titrated extract of Centella asiatica in phthalic anhydride-induced allergic dermatitis animal model. Int. J. Mol. Sci. 2017, 18, 738. [Google Scholar] [CrossRef] [PubMed]
- Pittella, F.; Dutra, R.C.; Junior, D.D.; Lopes, M.T.; Barbosa, N.R. Antioxidant and cytotoxic activities of Centella asiatica (L) Urb. Int. J. Mol. Sci. 2009, 10, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Morikawa, T.; Ueda, H.; Yoshikawa, M. Medicinal Foodstuffs. XXVII. Saponin constituents Gotu Kola (2): Structures of new ursane- and oleanane-type triterpene oligoglycosides, centellasaponin B, C, and D, from Centella asiatica cultivated in Sri Lanka. Chem. Pharm. Bull. 2001, 49, 1368–1371. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, R.S.; Tripathi, S.; Singh, J.; Narnoliya, L.K.; Sangwan, N.S. De novo sequencing and assembly of Centella asiatica leaf transcriptome for mapping of structural, functional and regulatory genes with special reference to secondary metabolism. Gene 2013, 525, 58–76. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, P.I.; Owens, I.S.; Burchell, B.; Bock, K.W.; Bairoch, A.; Bélanger, A.; Fournel-Gigleux, S.; Green, M.; Hum, D.W.; Iyanagi, T.; et al. The UDP glycosyltransferase gene superfamily: Recommended nomenclature update based on evolutionary divergence. Pharmacogenetics 1997, 7, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Gong, C.; Ren, L.; Li, X.; Song, D.; Zheng, G. A simple and effective strategy for solving the problem of inclusion bodies in recombinant protein technology: His-tag deletions enhance soluble expression. Appl. Microbiol. Biotechnol. 2013, 97, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Azerad, R. Chemical structures, production and enzymatic transformations of sapogenins and saponins from Centella asiatica (L.) Urban. Fitoterapia 2016, 114, 168–187. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.H.; Holmes, M.H. DNA sequence quality trimming and vector removal. Bioinfomatics 2001, 17, 1093–1104. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.T.; Bang, K.H.; Shin, Y.S.; Lee, M.J.; Jung, S.J.; Hyun, D.Y.; Kim, Y.C.; Seong, N.S.; Cha, S.W.; Hwang, B. Enhanced production of asiaticoside from hairy root cultures of Centella asiatica (L.) Urban elicited by methyl jasmonate. Plant Cell Rep. 2007, 26, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Ji, S.H.; Kim, G.S.; Song, K.S.; Um, Y.; Kim, O.T.; Lee, Y.; Hong, C.P.; Shin, D.H.; Kim, C.K.; et al. Global Profiling of Various Metabolites in Platycodon grandiflorum by UPLC-QTOF/MS. Int. J. Mol. Sci. 2015, 16, 26786–26796. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Public Database | No. of Unique Sequences | Annotation Percentage (%) |
|---|---|---|
| TAIR | 14,051 | 35.0 |
| Swiss-Prot | 15,171 | 37.9 |
| Nr | 17,807 | 44.4 |
| Nt | 16,709 | 41.7 |
| Total | 25,978 | 64.8 |
| Gene Name | Unique Sequence | EST Number | RPKM | Putative Function and Source | E-Value |
|---|---|---|---|---|---|
| UGT73AH1 | Isotig09551 | 235 | 118.01 | UDP-glycosyltransferase superfamily protein, putative (Theobroma cacao) | 0 |
| UGT73A21 | Isotig08753 | 232 | 101.59 | Glycosyltransferase UGT7 (Bupleurum chinense) | 0 |
| UGT74AH1 | Isotig09014 | 130 | 59.50 | UDP-glycosyltransferase 74E2-like (Vitis vinifera) | 0 |
| UGT73A22 | Isotig09265 | 115 | 55.09 | UDP-glycosyltransferase UGT7 (Bupleurum chinense) | 0 |
| UGT74AG2 | Isotig08223 | 136 | 53.51 | PREDICTED: UDP-glycosyltransferase 74E1-like (Solanum lycopersicum) | 0 |
| UGT73AH2 | Isotig08281 | 72 | 28.72 | UDP-glycosyltransferase 73C2 (Vitis vinifera) | 0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, O.T.; Jin, M.L.; Lee, D.Y.; Jetter, R. Characterization of the Asiatic Acid Glucosyltransferase, UGT73AH1, Involved in Asiaticoside Biosynthesis in Centella asiatica (L.) Urban. Int. J. Mol. Sci. 2017, 18, 2630. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122630
Kim OT, Jin ML, Lee DY, Jetter R. Characterization of the Asiatic Acid Glucosyltransferase, UGT73AH1, Involved in Asiaticoside Biosynthesis in Centella asiatica (L.) Urban. International Journal of Molecular Sciences. 2017; 18(12):2630. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122630
Chicago/Turabian StyleKim, Ok Tae, Mei Lan Jin, Dae Young Lee, and Reinhard Jetter. 2017. "Characterization of the Asiatic Acid Glucosyltransferase, UGT73AH1, Involved in Asiaticoside Biosynthesis in Centella asiatica (L.) Urban" International Journal of Molecular Sciences 18, no. 12: 2630. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122630