
Anti-Osteoclastic Activity of Artemisia capillaris Thunb. Extract Depends upon Attenuation of Osteoclast Differentiation and Bone Resorption-Associated Acidification Due to Chlorogenic Acid, Hyperoside, and Scoparone


Abstract
:1. Introduction
2. Results
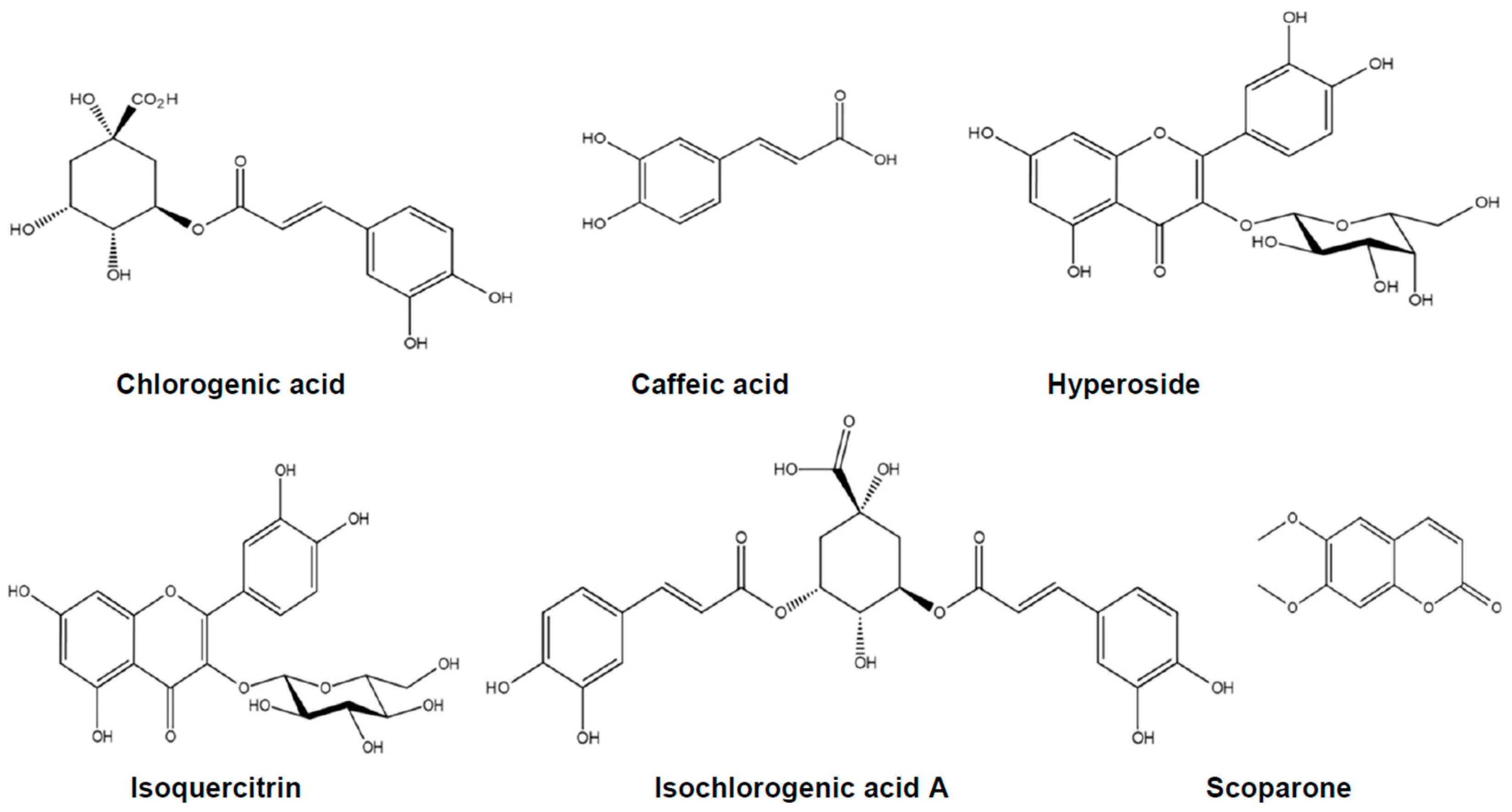
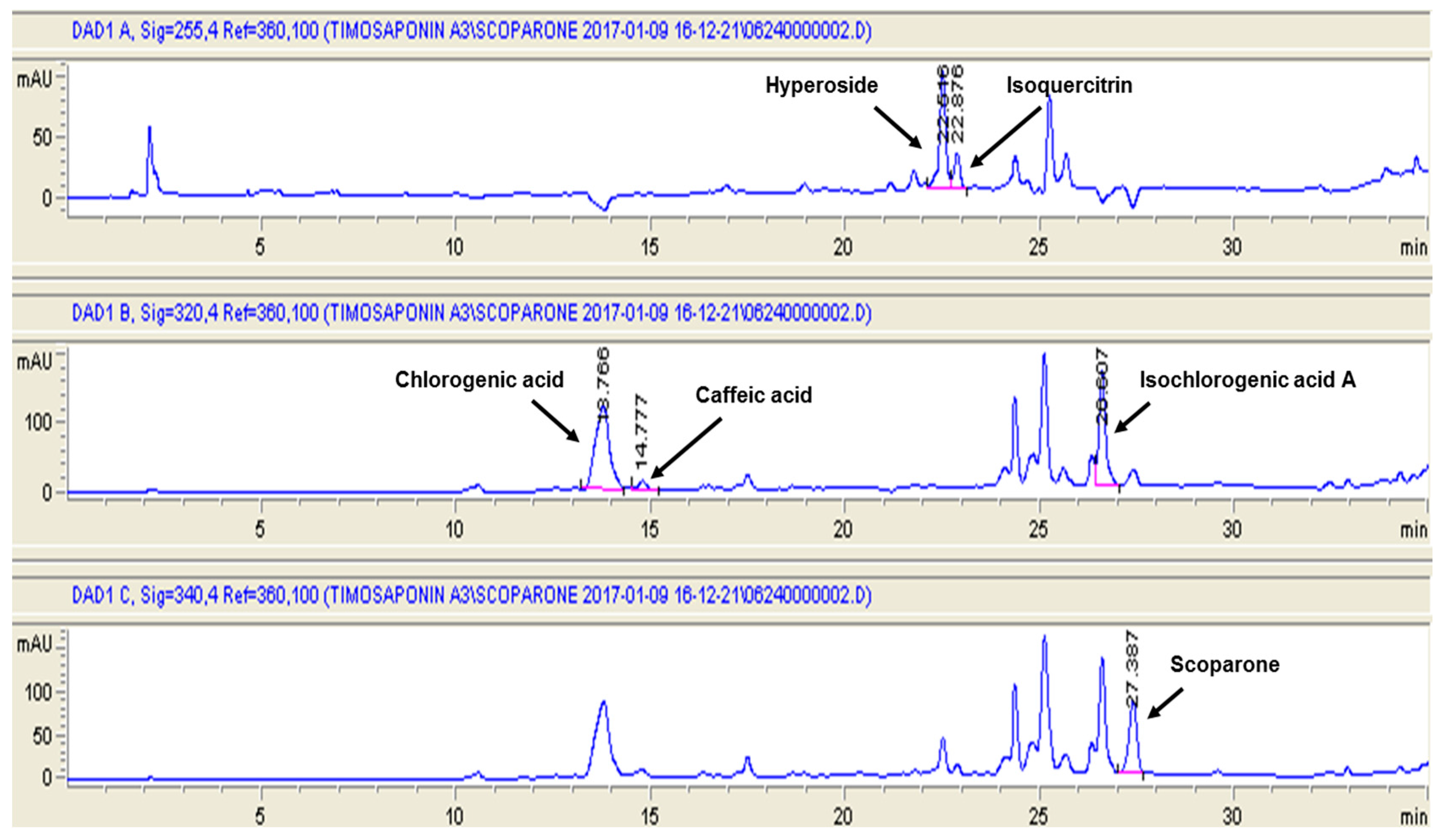
2.1. The Content of Six Marker Compounds in Artemisia capillaris Hydroethanolic Extract (ACHE)
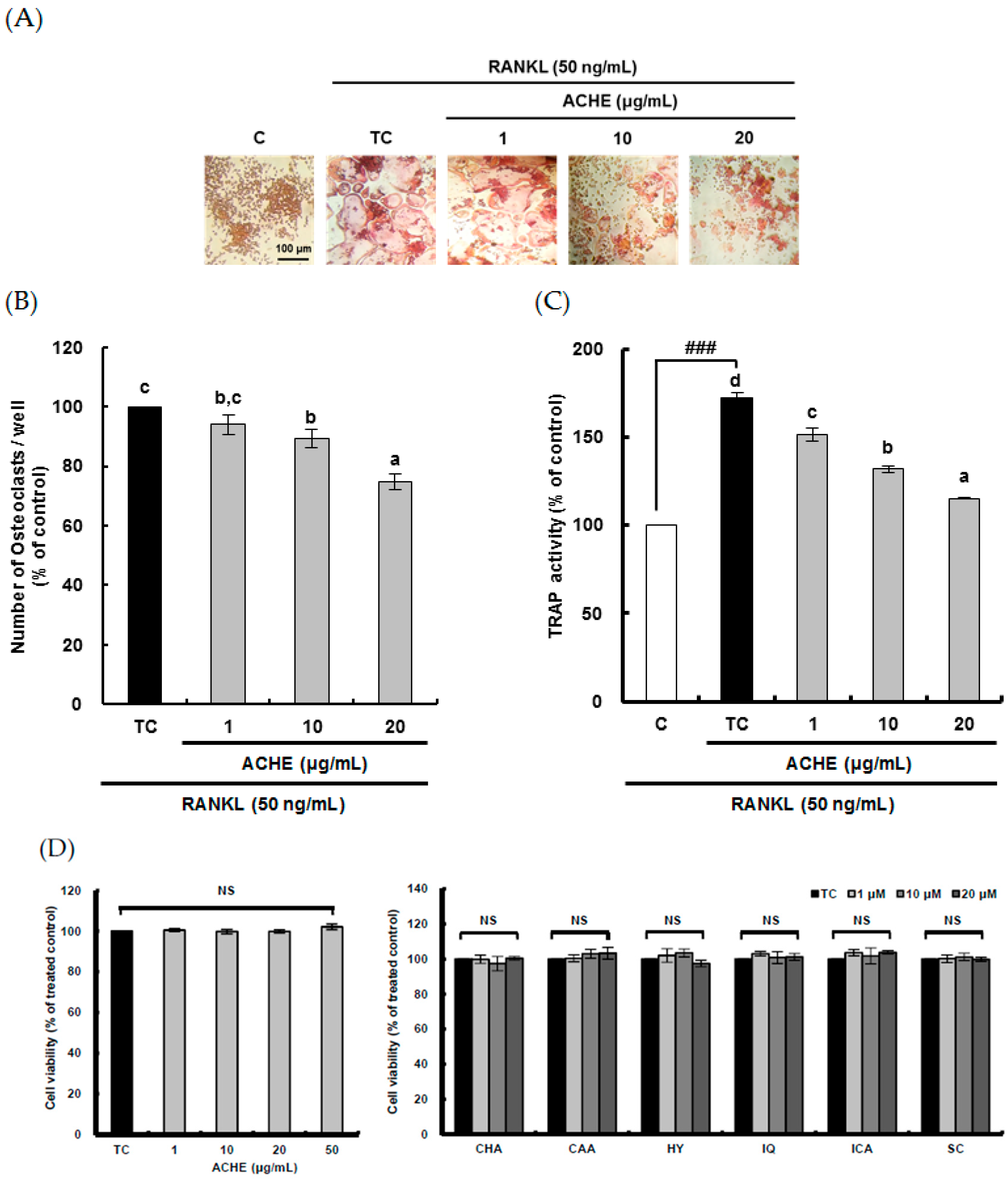
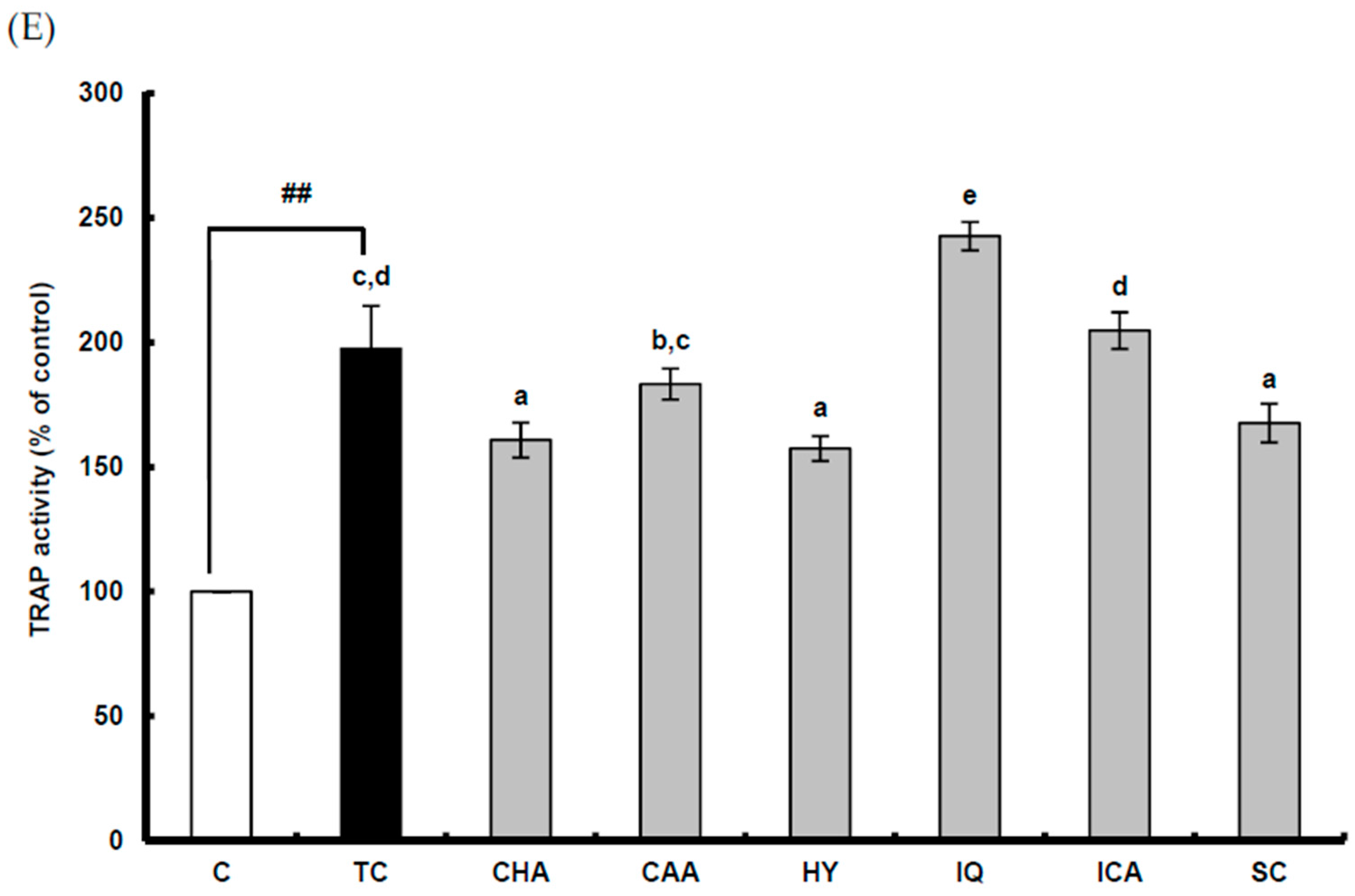
2.2. Suppressive Effect of ACHE and Its Six Marker Compounds on Osteoclast Differentiation
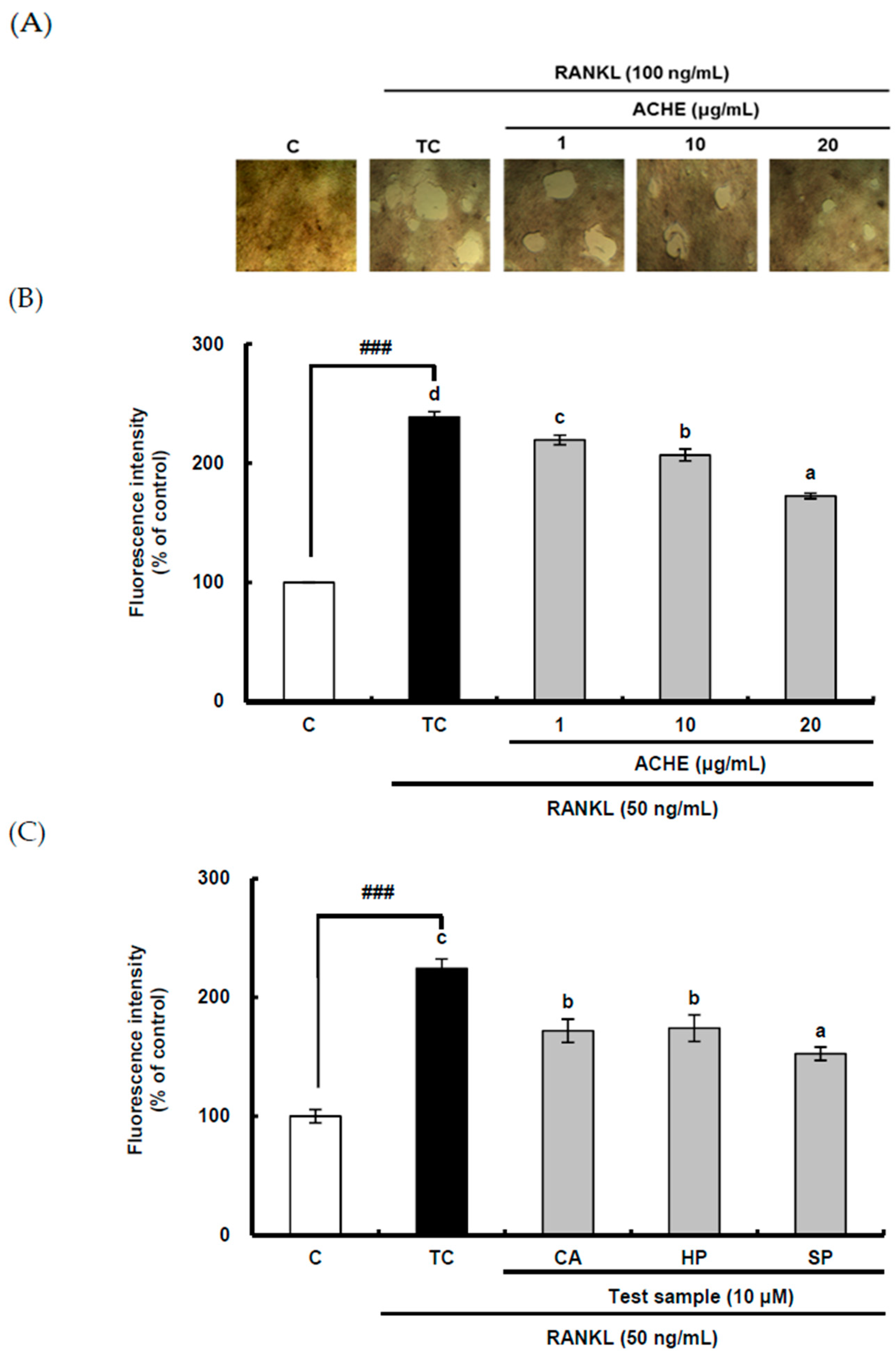
2.3. Inhibitory Effect of ACHE and Three Bioactive Compounds on Bone Resorption of Osteoclast
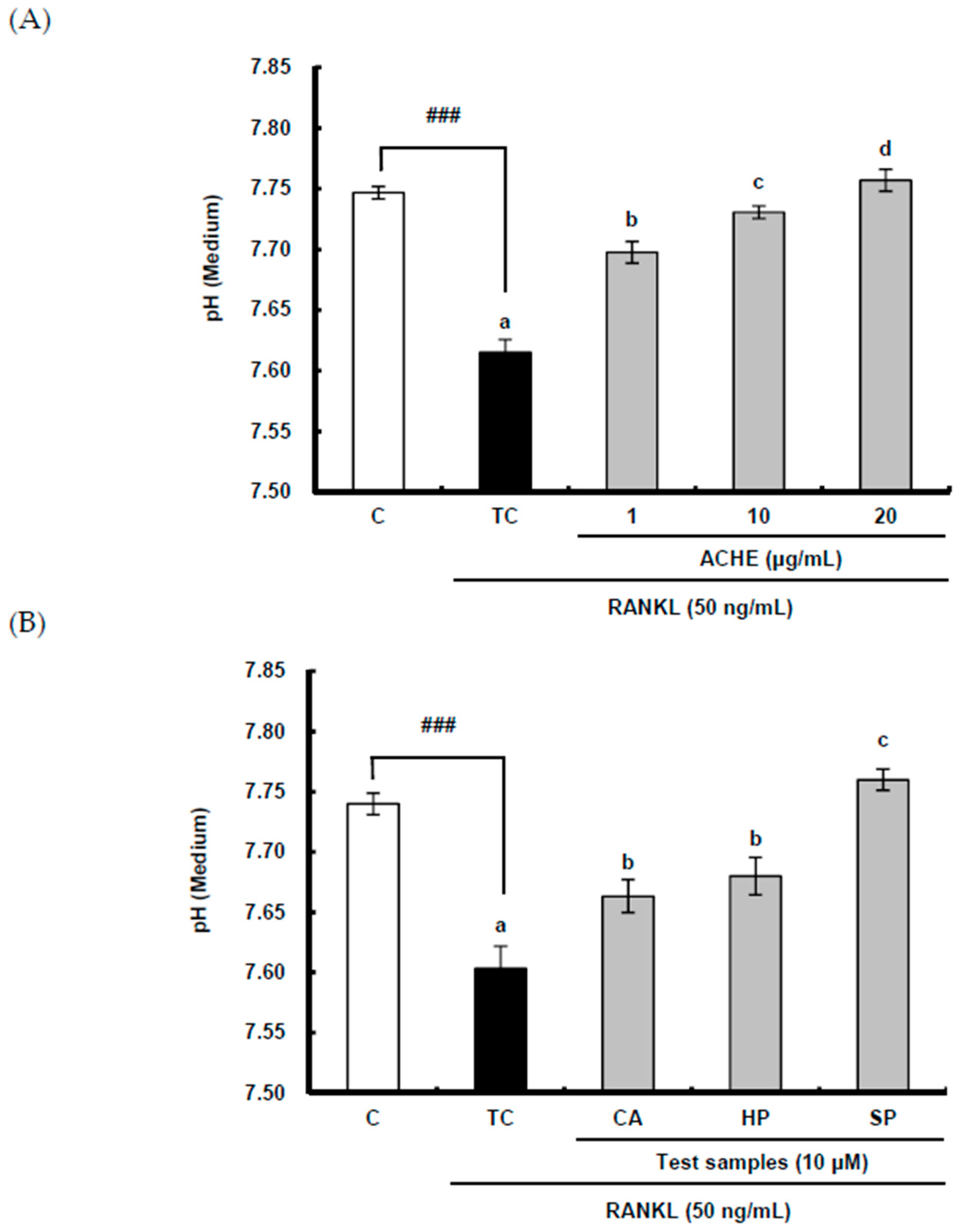
2.4. Suppressive Effect of ACHE and Three Bioactive Compounds on the Acidification by Osteoclasts
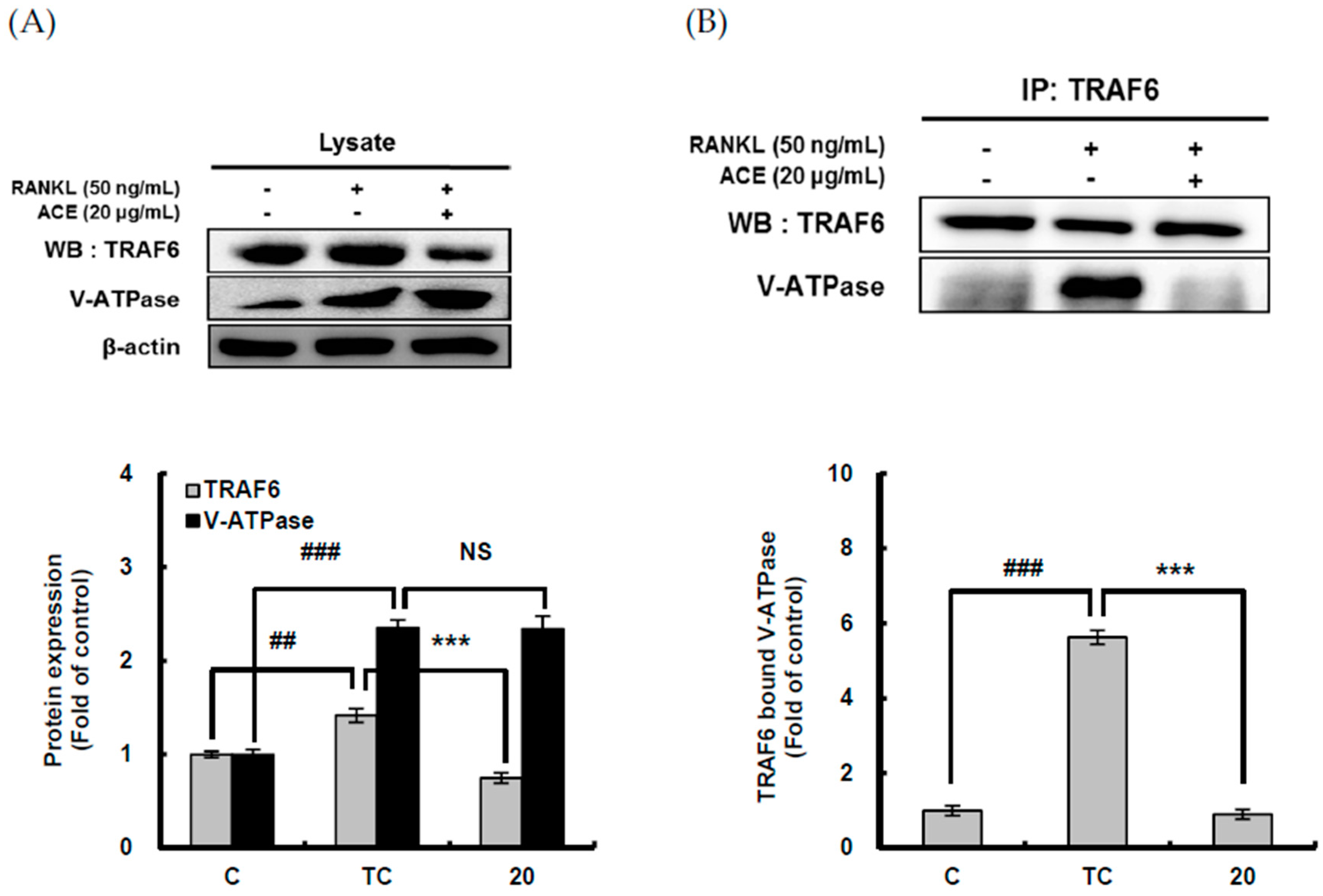
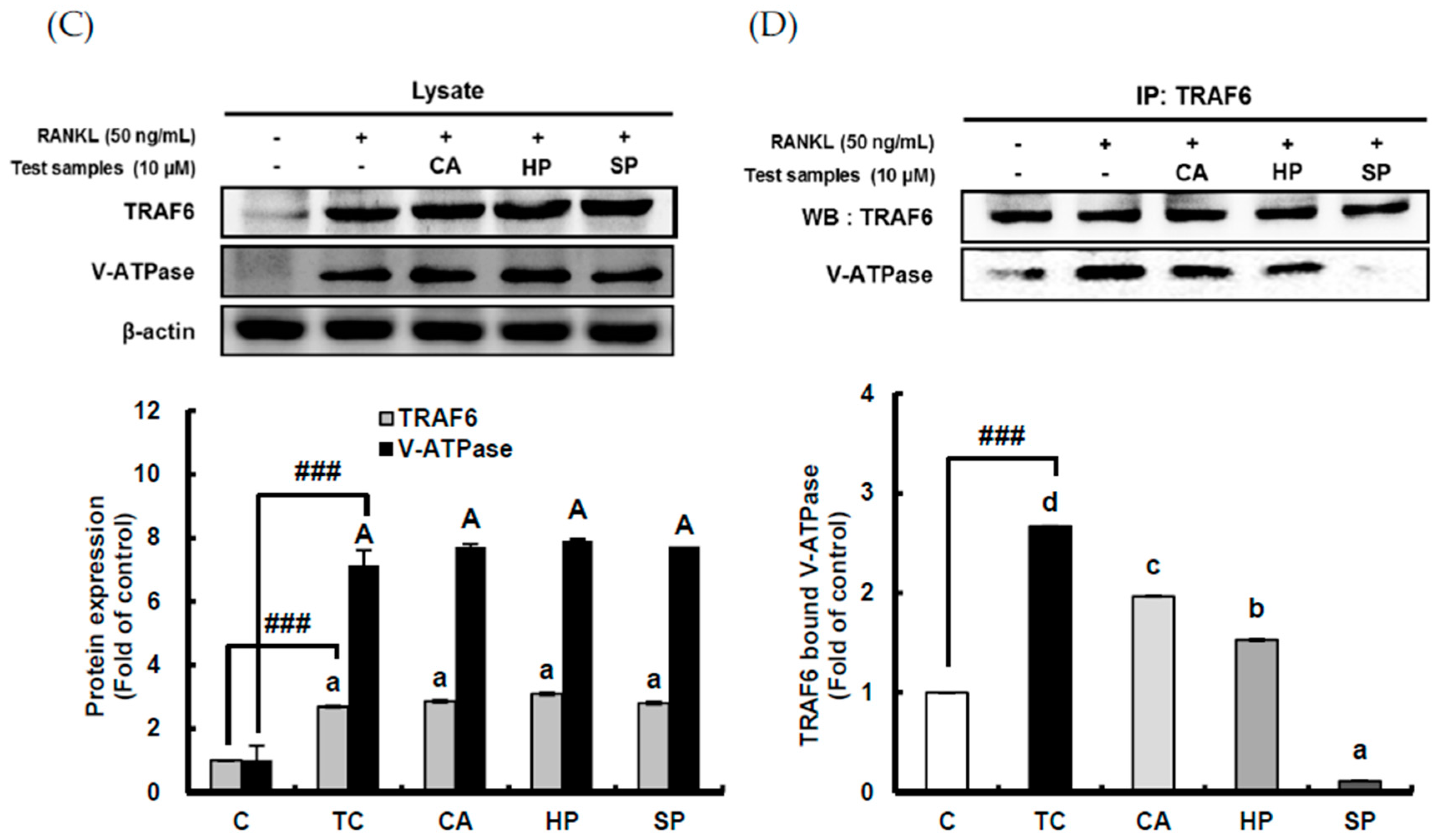
2.5. Down-Regulating Effect of ACHE and Three Bioactive Compounds on TRAF6 Expression and Its Binding to V-ATPase
3. Discussion
4. Materials and Methods
4.1. Reagents and Cell Culture Materials
4.2. Preparation of A. capillaris Hydroethanolic Extract (ACHE)
4.3. High Performance Liquid Chromatography (HPLC) Analysis
4.4. Cell Cytotoxicity by MTT Assay
4.5. TRAP Staining and Activity
4.6. Bone Resorption Assay
4.7. pH Measurement
4.8. Immunoprecipitation and Immunoblotting
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eriksen, E.F. Cellular mechanism of bone remodeling. Rev. Endocr. Metab. Disord. 2010, 11, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.B.; Zheng, C.J.; Qin, L.P.; Sun, L.N.; Han, T.; Jiao, L.; Zhang, Q.Y.; Wu, J.Z. Antiosteporotic activity of anthraquinones from Morinda officinalis on osteoblasts and osteoclasts. Molecules 2009, 14, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Sakai, E.; Shimada-Sugawara, M.; Yamaguchi, Y.; Skamoto, H.; Fumimoto, R.; Fukuma, Y.; Nishishita, K.; Okamoto, K.; Tsukuba, T. Fisetin inhibits osteoclastogenesis through prevention of RANKL-induced ROS production by Nrf2-mediated up-regulation of phase II antioxidant enzymes. J. Pharmacol. 2013, 121, 288–298. [Google Scholar] [CrossRef]
- Boyle, W.; Simonet, W.S.; Lacey, D. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.S.; Lee, S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, A.V.; Heymann, D. Osteoclastic acidification pathways during bone resorption. Bone 2002, 30, 533–540. [Google Scholar] [CrossRef]
- Ryu, J.Y.; Kim, H.S.; Lee, S.K.; Chang, E.J.; Kim, H.J.; Kim, H.H. Proteomic identification of the TRAF6 regulation of vacuolar ATPase for osteoclast function. Proteomics 2005, 5, 4152–4160. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Kim, D.W.; Yun, N.; Choi, J.S.; Islam, M.N.; Kim, Y.S.; Lee, S.M. Protective effects of hyperoside against carbon tetrachloride-induced liner damage in mice. J. Nat. Prod. 2011, 74, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Ying, L.; Kim, B.; Zhang, H.; Hwangbo, K.; Paio, D.G.; Chi, M.J.; Woo, M.H.; Choi, J.S.; Lee, J.H.; et al. High-performance liquid chromatographic analysis for quantitation of marker compounds of Artemisia capillaris Thunb. Arch. Pharm. Res. 2012, 12, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.K.; Lee, H.Y.; Seo, C.S.; Lim, H.S.; Lee, J.K.; Lee, M.Y.; Shin, H.K. Artemisia capillaris inhibits atopic dermatitis-like skin lesions in Dermatophagoides farina-sensitized Nc/Nga mice. BMC Complement. Altern. Med. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.Y.; Kim, B.J.; Lee, K.T.; Inn, K.S.; Lee, J.H. A survey of therapeutic effects of Artemisia capillaris in liver diseases. Evid. Based Complement. Altern. Med. 2015, 2015, 728137. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Lee, H.J.; Kim, Y.A.; Park, K.E.; Ahn, J.W.; Lee, B.J.; Moon, S.G.; Seo, Y.W. Antioxidant activity of Artemisia capillaris Thunberg. Food Sci. Biotechnol. 2004, 13, 328–331. [Google Scholar]
- Hong, J.H.; Lee, J.W.; Park, J.H.; Lee, I.S. Antioxidative and cytoprotective effects of Artemisia capillaris fractions. Biofactors 2007, 3, 43–53. [Google Scholar] [CrossRef]
- Cha, J.D.; Moon, S.E.; Kim, H.Y.; Cha, I.H.; Lee, K.Y. Essential oil of Artemisia capillaris induces apoptosis in KB cells via mitochondrial stress and caspase activation mediated by MAPK-stimulated signaling pathway. J. Food Sci. 2009, 74, T75–T81. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.D.; Jung, E.K.; Kil, B.S.; Lee, K.Y. Chemical Composition and ant-bacterial activity of essential oil from Artemisia feddei. J. Microbiol. Biotechnol. 2007, 17, 2061–2065. [Google Scholar] [PubMed]
- Yang, C.C.; Lee, M.R.; Hsu, S.L.; Chang, C.M.J. Supercritical fluids extraction of capillarism from Artemisia capillaris and its inhibition of in vitro growth of hepatoma cells. J. Supercrit. Fluid 2007, 42, 96–103. [Google Scholar] [CrossRef]
- Chu, C.Y.; Tseng, T.H.; Hwang, J.M.; Chou, F.P.; Wang, C.J. Protective effects of capillarism on tetra-butylhydroperoxide-induced oxidative damage in rat primary hepatocytes. Arch. Toxicol. 1999, 73, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Tien, Y.C.; Liao, J.C.; Chiu, C.S.; Huang, T.H.; Huang, C.Y.; Chang, W.T.; Peng, W.H. Esculetin ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. Int. J. Mol. Sci. 2011, 12, 4053–4067. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.J.; Shim, K.S.; Ma, J.Y. Artemisia capillaris alleviates bone loss by stimulating osteoblast mineralization and suppressing osteoclast differentiation and bone resorption. Am. J. Chin. Med. 2016, 44, 1675. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jang, H.D. Scoparone attenuates RANKL-induced osteoclastic differentiation through controlling reactive oxygen species production and scavenging. Exp. Cell Res. 2015, 331, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.G.; Chen, K.M.; Xian, C.J. Functions and action mechanisms of flavonoids genistein and icarin in regulating bone remodeling. Cell Physiol. 2012, 228, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Ahn, J.Y.; Hasegawa, S.I.; Cha, B.Y.; Yonezawa, T.; Nagai, K.; Seo, H.J.; Jeon, W.B.; Woo, J.T. Inhibitory effect of luteolin on osteoblast differentiation and function. Cytotechnology 2009, 61, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Ryu, S.Y.; Bae, M.A.; Choi, J.S.; Min, Y.K.; Kim, S.H. Baicalein inhibits osteoclast differentiation and induces mature osteoclast apoptosis. Food. Chem. Toxicol. 2008, 46, 3375–3382. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jin, H.; Shim, H.E.; Kim, H.N.; Ha, H.; Lee, Z.H. Epogallocatechin-3-gallate inhibits osteoclastogenesis by down-regulating c-Fos expression and suppressing the nuclear factor-κB signal. Mol. Pharmacol. 2010, 77, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Ding, Y.; Yan, X.T.; Kim, Y.H.; Jang, H.D. Scopoletin and scopolin isolated from Artemisia iwayomogi suppress differentiation of osteoclastic macrophage RAW 264.7 cells by scavenging reactive oxygen species. J. Nat. Prod. 2013, 76, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.H.; Lee, I.S. Effects of Artemisia capillaris fractions on oxidative stress and antioxidant enzyme in high-fat diet induced obese mice. Chem. Biol. Int. 2009, 179, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.C.; Lee, C.; Kim, J.Y.; Oh, H.M.; So, H.S.; Lee, M.S.; Rho, M.C.; Oh, J.M. Chlorogenic acid inhibits osteoclast differentiation and bone resorption by down-regulation of receptor activator of nuclear factor κ-B ligand-induced nuclear factor of activated T cells c1 expression. Biol. Pharm. Bull. 2013, 36, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.L.; Kang, S.W.; Kang, M.K.; Gong, J.H.; Lee, E.S.; Han, S.J.; Kang, Y.H. Osteoblastogenesis and osteoprotection enhanced by flanovolignin silibinin in osteoblasts and osteoclasts. J. Cell Biochem. 2011, 113, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.L.; Kang, M.K.; Gong, J.H.; Park, S.H.; Han, S.Y.; Kang, Y.H. Novel antiosteoclastogenetic activity of phloretin antagonizing RANKL-induced osteoclast differentiation of murine macrophages. Mol. Nutr. Food Res. 2012, 56, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.B.; Cho, M.K.; Lee, W.Y.; Kang, K.W. Overexpression of c-myc induces epithelial mesencchymal transition in mammary epithelial cells. Cancer Lett. 2010, 293, 230–239. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Content (1) (mg/g) |
|---|---|
| Chlorogenic acid | 38.526 ± 0.927 |
| Caffeic acid | 0.515 ± 0.048 |
| Hyperoside | 4.072 ± 0.220 |
| Isoquercitrin | 3.031 ± 0.148 |
| Isochlorogenic acid | 13.898 ± 0.667 |
| Scoparone | 6.589 ± 0.193 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Lee, J.-Y.; Kwon, Y.-I.; Jang, H.-D. Anti-Osteoclastic Activity of Artemisia capillaris Thunb. Extract Depends upon Attenuation of Osteoclast Differentiation and Bone Resorption-Associated Acidification Due to Chlorogenic Acid, Hyperoside, and Scoparone. Int. J. Mol. Sci. 2017, 18, 322. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18020322
Lee S-H, Lee J-Y, Kwon Y-I, Jang H-D. Anti-Osteoclastic Activity of Artemisia capillaris Thunb. Extract Depends upon Attenuation of Osteoclast Differentiation and Bone Resorption-Associated Acidification Due to Chlorogenic Acid, Hyperoside, and Scoparone. International Journal of Molecular Sciences. 2017; 18(2):322. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18020322
Chicago/Turabian StyleLee, Sang-Hyun, Jung-Yun Lee, Young-In Kwon, and Hae-Dong Jang. 2017. "Anti-Osteoclastic Activity of Artemisia capillaris Thunb. Extract Depends upon Attenuation of Osteoclast Differentiation and Bone Resorption-Associated Acidification Due to Chlorogenic Acid, Hyperoside, and Scoparone" International Journal of Molecular Sciences 18, no. 2: 322. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18020322