
High-Frequency Repetitive Transcranial Magnetic Stimulation (rTMS) Improves Functional Recovery by Enhancing Neurogenesis and Activating BDNF/TrkB Signaling in Ischemic Rats

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
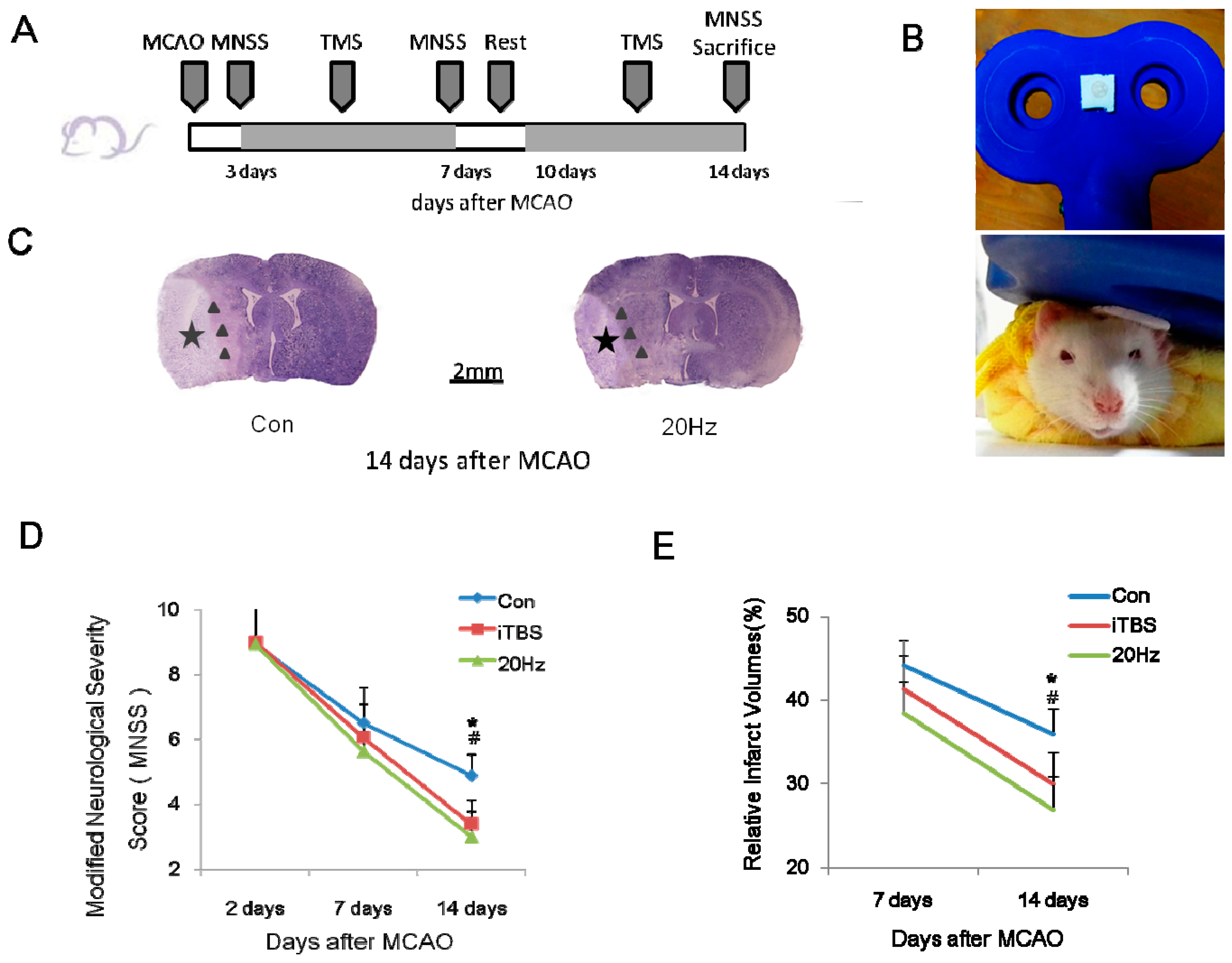
2.1. rTMS Improves the Recovery of Neurological Function
2.2. rTMS Reduces the Volume of the Infarct Area after an Ischemic Stroke
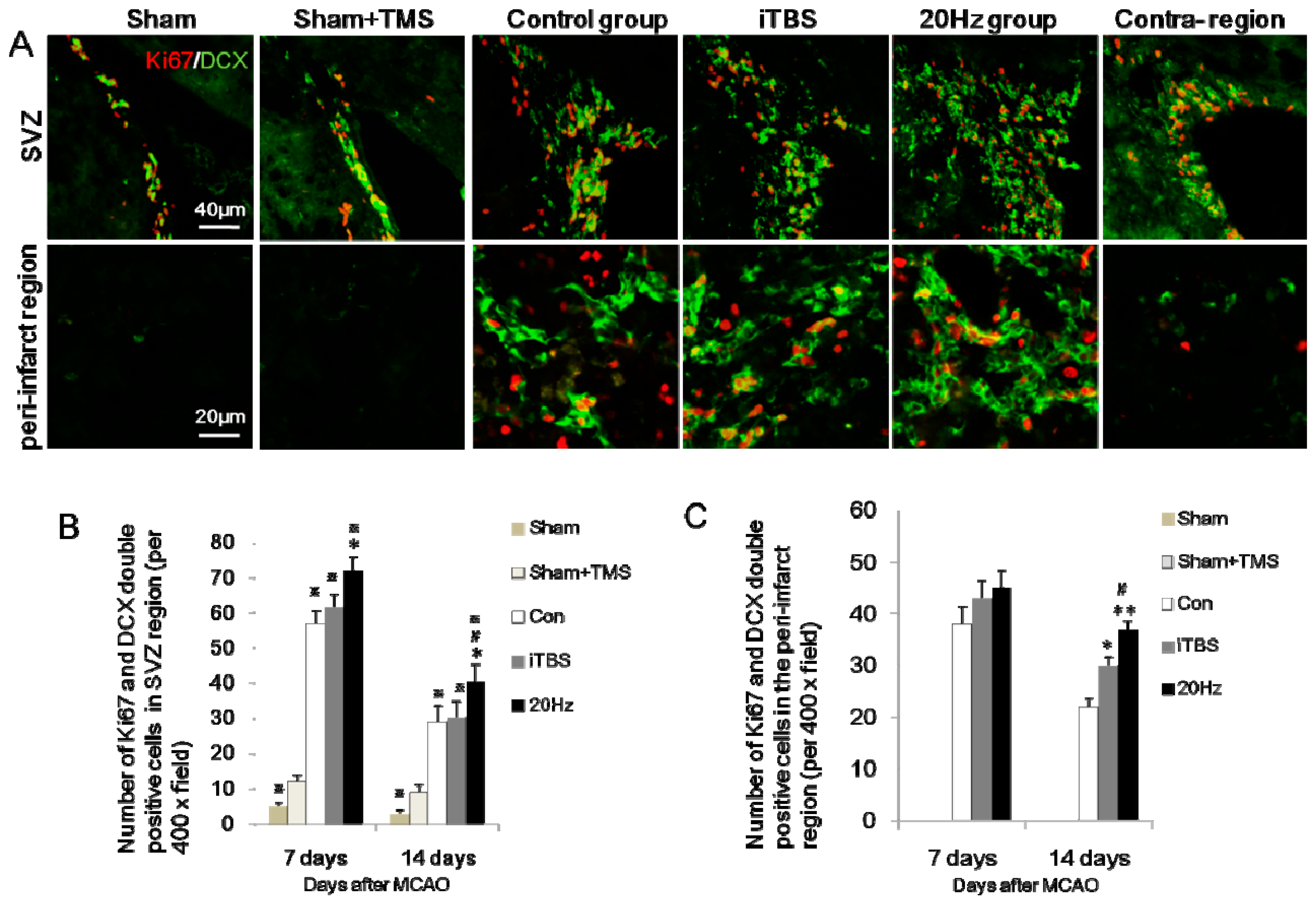
2.3. rTMS Increases the Migration of Neural Progenitor Cells in the Ipsilateral SVZ and Peri-Infarct Striatum
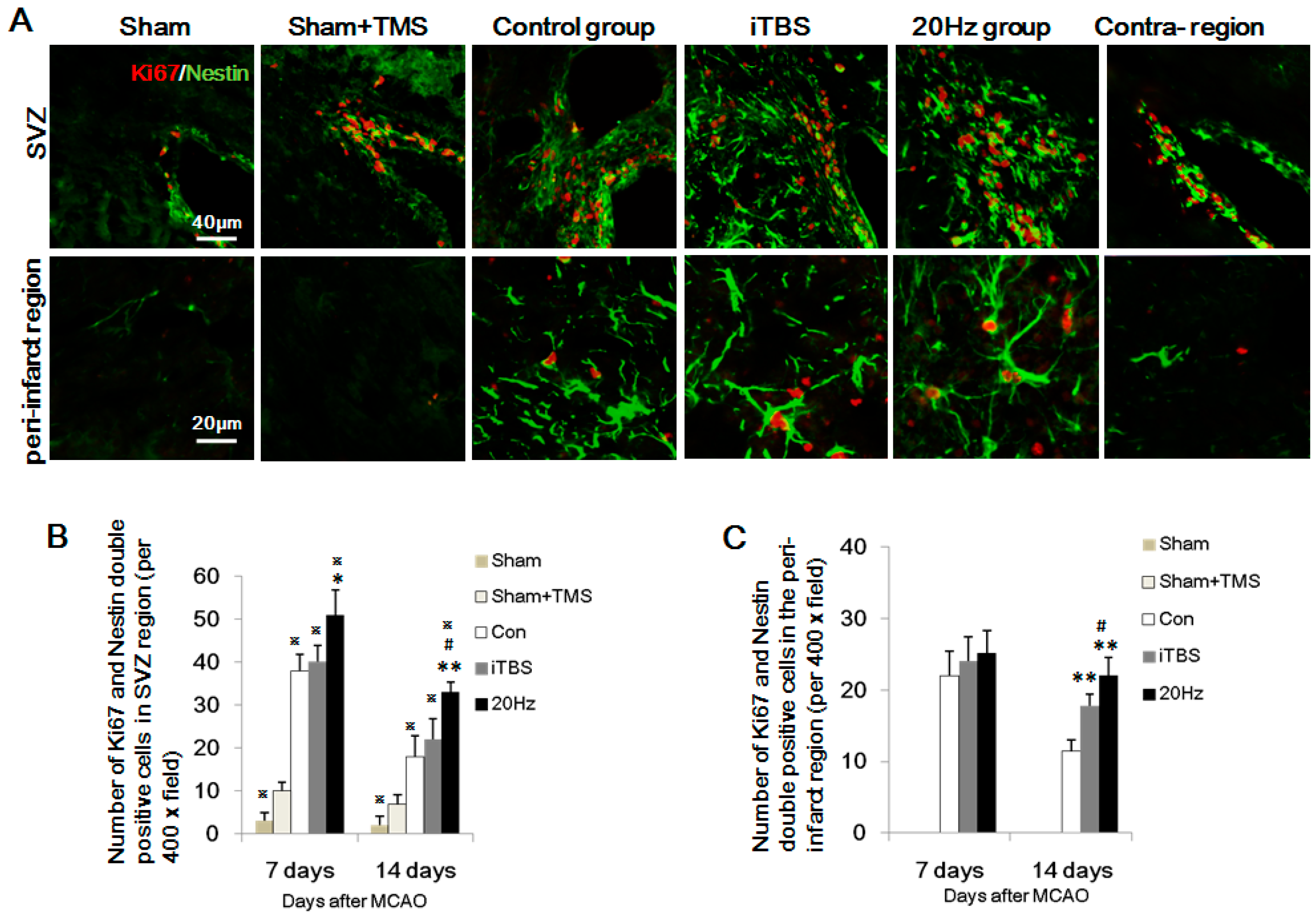
2.4. rTMS Increases NSC Proliferation in the SVZ and Peri-Infarct Striatum
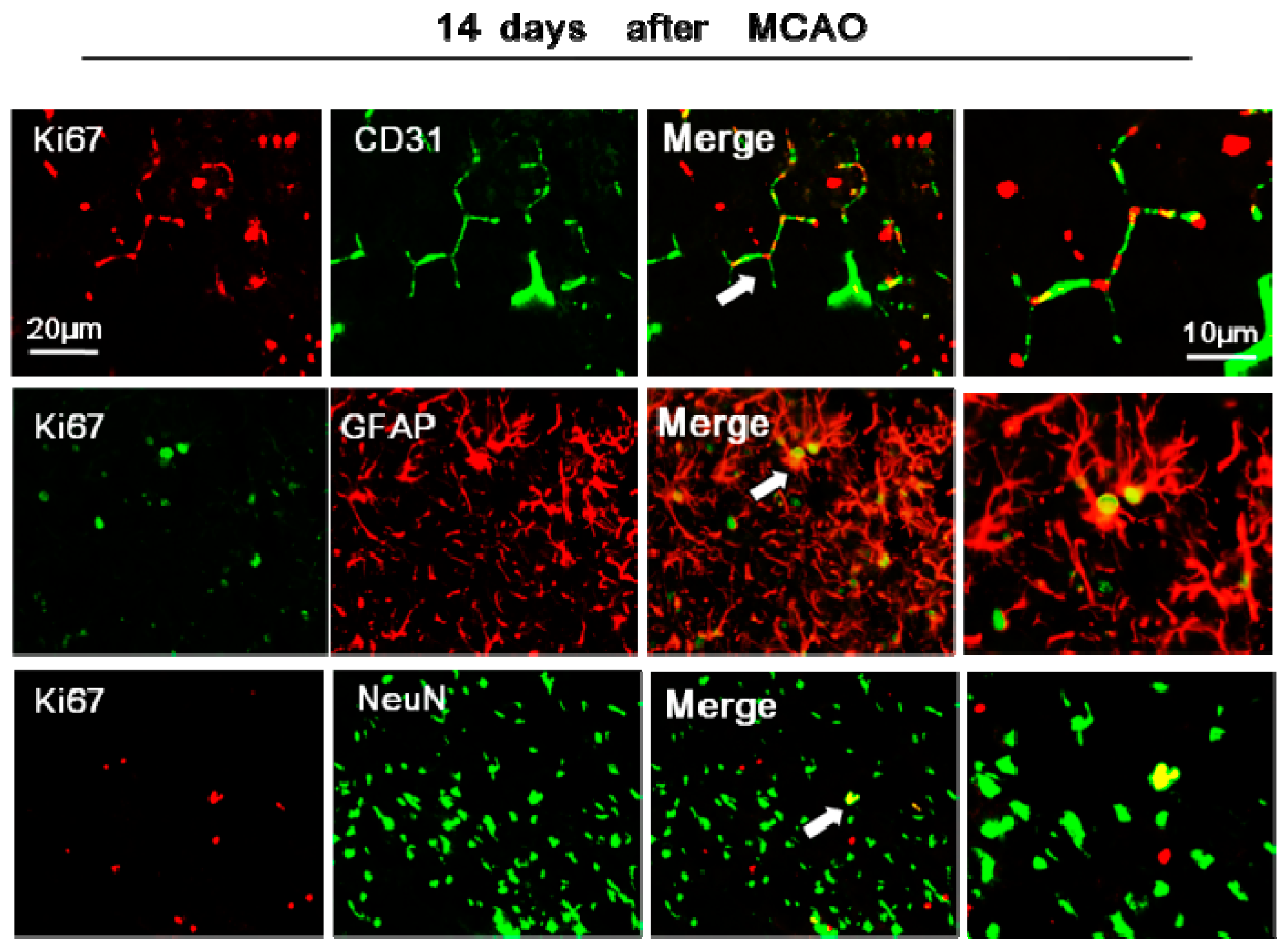
2.5. rTMS Promotes NSC Differentiation in the Peri-Infarct Striatum
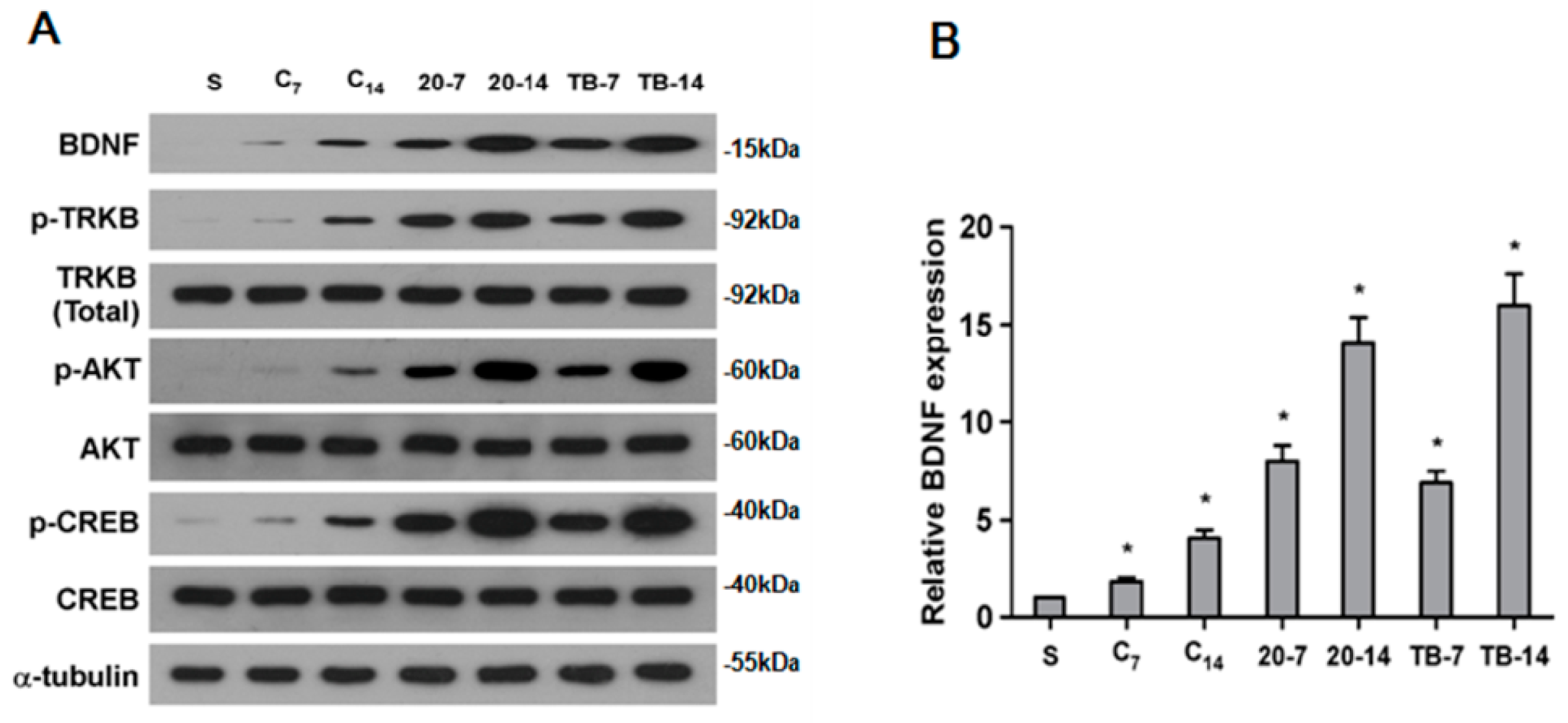
2.6. rTMS Activates BDNF/TrkB Signaling Pathway in Ischemic Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Neurobehavioral Evaluation
4.3. Repetitive Transcranial Magnetic Stimulation (rTMS)
4.4. Experimental Grouping
4.5. Tissue Preparation for Histochemistry
4.6. Infarct Volume Measurement
4.7. Immunofluorescence Analysis
4.8. Western Blotting
4.9. Data Analysis and Statistics
4.9.1. Image Analysis and Quantification
4.9.2. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| rTMS | repetitive transcranial magnetic stimulation |
| BDNF | brain-derived neurotrophic factor |
| TrkB | tropomyosin-related kinase B |
| MCAO | middle cerebral artery occlusion |
| DCX | doublecortin |
| GFAP | glial fibrillary acidic protein |
| NSC | neural stem cell |
| MNSS | modified neurological severity score |
| SGZ | subgranular zone |
| SVZ | subventricular zone |
| DG | dentate gyrus |
| iTBS | intermittent theta burst stimulation |
| LTP | long-term potentiation |
| LTD | long-term depression |
| RMT | resting motor threshold |
| MEP | motor evoked potentials |
| DAPI | 4′,6-diamidino-2-phenylindole |
| PBS | phosphate-buffered saline |
| TBST | Tris-buffered saline with Tween 20 |
| CREB | cAMP responsive element-binding protein |
| ANOVA | analysis of variance |
References
- Langhorne, P.; Coupar, F.; Pollock, A. Motor recovery after stroke: A systematic review. Lancet Neurol. 2009, 8, 741–754. [Google Scholar] [CrossRef]
- Thorsén, A.M.; Holmqvist, L.W.; de Pedro-Cuesta, J.; von Koch, L. A randomized controlled trial of early supported discharge and continued rehabilitation at home after stroke: Five-year follow-up of patient outcome. Stroke 2005, 36, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Bellenchi, G.C.; Volpicelli, F.; Piscopo, V.; Perrone-Capano, C.; di Porzio, U. Adult neural stem cells: An endogenous tool to repair brain injury? J. Neurochem. 2013, 124, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, A.; Collin, T.; Kirik, D.; Kokaia, Z.; Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 2002, 8, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Gage, F.H. Mammalian neural stem cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Thored, P.; Arvidsson, A.; Cacci, E.; Ahlenius, H.; Kallur, T.; Darsalia, V.; Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Persistent production of neurons from adult brain stem cells during recovery after stroke. Stem Cells 2006, 24, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Yozbatiran, N.; Alonso-Alonso, M.; See, J.; Demirtas-Tatlidede, A.; Luu, D.; Motiwala, R.R.; Pascual-Leone, A.; Cramer, S.C. Safety and behavioral effects of high-frequency repetitive transcranial magnetic stimulation in stroke. Stroke 2009, 40, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Houdayer, E.; Degardin, A.; Cassim, F.; Bocquillon, P.; Derambure, P.; Devanne, H. The effects of low- and high-frequency repetitive TMS on the input/output properties of the human corticospinal pathway. Exp. Brain Res. 2008, 187, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta burst stimulation of the human motor cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.W.; Hoy, K.E.; Fitzgerald, P.B. Theta-burst stimulation: A new form of TMS treatment for depression? Depress. Anxiety 2015, 32, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.D.; Boddington, L.J.; Igelström, K.M. Utility of intracerebral theta burst electrical stimulation to attenuate interhemispheric inhibition and to promote motor recovery after cortical injury in an animal model. Exp. Neurol. 2014, 261, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y. Repetitive transcranial magnetic stimulation (rTMS) for higher brain function deficits. Brain Nerve 2016, 68, 1459–1470. [Google Scholar] [PubMed]
- Guo, F.; Han, X.; Zhang, J.; Zhao, X.; Lou, J.; Chen, H.; Huang, X. Repetitive transcranial magnetic stimulation promotes neural stem cell proliferation via the regulation of miR-25 in a rat model of focal cerebral ischemia. PLoS ONE 2014, 10, e109267. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.B.; Toschi, N.; Kresse, A.E.; Post, A.; Keck, M.E. Long-term repetitive transcranial magnetic stimulation increases the expression of brain-derived neurotrophic factor and cholecystokinin mRNA, but not neuropeptide tyrosine mRNA in specific areas of rat brain. Neuropsychopharmacology 2000, 23, 205–215. [Google Scholar] [CrossRef]
- Berretta, A.; Tzeng, Y.C.; Clarkson, A.N. Post-stroke recovery: The role of activity-dependent release of brain-derived neurotrophic factor. Expert Rev. Neurother. 2014, 14, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Bath, K.G.; Lee, F.S. Neurotrophic factor control of adult SVZ neurogenesis. Dev. Neurobiol. 2010, 70, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Jansson, L.C.; Louhivuori, L.; Wigren, H.K. Brain-derived neurotrophic factor increases the motility of a particular N-methyl-d-aspartate/GABA-responsive subset of neural progenitor cells. Neuroscience 2012, 224, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Balthazart, J.; Ball, G.F. Endogenous versus exogenous markers of adult neurogenesis in canaries and other birds: Advantages and disadvantages. J. Comp. Neurol. 2014, 522, 4100–4120. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Xiang, A.P.; Mao, F.F. Nestin is required for the proper self-renewal of neural stem cells. Stem Cells 2010, 28, 2162–2171. [Google Scholar] [CrossRef] [PubMed]
- Li, W.L.; Yu, S.P.; Ogle, M.; Ding, X.S.; Wei, L. Enhanced neurogenesis and cell migration following focal ischemia and peripheral stimulation in mice. Dev. Neurobiol. 2008, 68, 1474–1486. [Google Scholar] [CrossRef] [PubMed]
- Christie, K.J.; Turnley, A.M. Regulation of endogenous neural stem/progenitor cells for neural repair—Factors that promote neurogenesis and gliogenesis in the normal and damaged brain. Front. Cell. Neurosci. 2013, 6, 70. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.; Qu, Y.; Tao, Y.; Zhu, S. Effects of repetitive transcranial magnetic stimulation on hand function recovery and excitability of the motor cortex after stroke: A meta-analysis. Am. J. Phys. Med. Rehabil. 2014, 93, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Lee, Y.T.; Han, T.R. Mechanism of functional recovery after repetitive transcranial magnetic stimulation (rTMS) in the subacute cerebral ischemic rat model: Neural plasticity or anti-apoptosis? Exp. Brain Res. 2011, 214, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Barreto, G.; White, R.E.; Ouyang, Y. Astrocytes: Targets for neuroprotection in stroke. Cent. Nerv. Syst. Agents Med. Chem. 2011, 11, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chopp, M. Astrocytes, therapeutic targets for neuroprotection and neurorestoration in ischemic stroke. Prog. Neurobiol. 2016, 144, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Bagley, J.A.; Belluscio, L. Dynamic imaging reveals that brain-derived neurotrophic factor can independently regulate motility and direction of neuroblasts within the rostral migratory stream. Neuroscience 2010, 169, 1449–1461. [Google Scholar] [CrossRef] [PubMed]
- Schäbitz, W.R.; Steigleder, T.; Cooper-Kuhn, C.M. Intravenous brain-derived neurotrophic factor enhances poststroke sensorimotor recovery and stimulates neurogenesis. Stroke 2007, 38, 2165–2172. [Google Scholar] [CrossRef] [PubMed]
- Chiaramello, S.; Dalmasso, G.; Bezin, L.; Marcel, D.; Jourdan, F.; Peretto, P.; Fasolo, A.; de Marchis, S. BDNF/TrkB interaction regulates migration of SVZ precursor cells via PI3-K and MAP-K signalling pathways. Eur. J. Neurosci. 2007, 26, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Young, C.C.; Brooks, K.J.; Buchan, A.M.; Szele, F.G. Cellular and molecular determinants of stroke-induced changes in subventricular zone cell migration. Antioxid. Redox Signal. 2011, 14, 1877–1888. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Zhang, R.G.; Xue, F.; Wang, H.N.; Chen, Y.C.; Hu, G.T.; Peng, Y.; Peng, Z.W.; Tan, Q.R. Quetiapine and repetitive transcranial magnetic stimulation ameliorate depression-like behaviors and up-regulate the proliferation of hippocampal-derived neural stem cells in a rat model of depression: The involvement of the BDNF/ERK signal pathway. Pharmacol. Biochem. Behav. 2015, 136, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Hu, X.Q.; Zhang, L.Y.; Li, L.L.; Zheng, H.Q.; Li, M.L.; Zhang, Q.J. Physical exercise regulates neural stem cells proliferation and migration via SDF-1α/CXCR4 pathway in rats after ischemic stroke. Neurosci. Lett. 2014, 578, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Chio, C.C.; Chang, C.P. Long course hyperbaric oxygen stimulates neurogenesis and attenuates inflammation after ischemic stroke. Mediators Inflamm. 2013, 2013, 512978. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.C.; Chopp, M.; Zhang, L. Thymosin β4 improves functional neurological outcome in a rat model of embolic stroke. Neuroscience 2010, 169, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.J.; Lee, N.; Choi, C. Therapeutic effect of BDNF-overexpressing human neural stem cells (HB1.F3.BDNF) in a rodent model of middle cerebral artery occlusion. Cell. Transplant. 2013, 22, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Ridding, M.C.; Rothwell, J.C. Is there a future for therapeutic use of transcranial magnetic stimulation? Nat. Rev. Neurosci. 2007, 8, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Richter, L.; Neumann, G.; Oung, S.; Schweikard, A.; Trillenberg, P. Optimal coil orientation for transcranial magnetic stimulation. PLoS ONE 2013, 8, e60358. [Google Scholar] [CrossRef] [PubMed]
- Ogiue-Ikeda, M.; Kawato, S.; Ueno, S. Acquisition of ischemic tolerance by repetitive transcranial magnetic stimulation in the rat hippocampus. Brain Res. 2005, 1037, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Morales, L.; Nowak, D.A.; Kammer, T.; Wolf, R.C.; Schönfeldt-Lecuona, C. Mechanisms and applications of theta-burst rTMS on the human motor cortex. Brain Topogr. 2010, 22, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Hoogendam, J.M.; Ramakers, G.M.; Di Lazzaro, V. Physiology of repetitive transcranial magnetic stimulation of the human brain. Brain Stimul. 2010, 3, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Benninger, D.H.; Iseki, K.; Kranick, S. Controlled study of 50-Hz repetitive transcranial magnetic stimulation for the treatment of Parkinson disease. Neurorehabil. Neural Repair 2012, 26, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Kanno, M.; Matsumoto, M.; Togashi, H. Effects of repetitive transcranial magnetic stimulation on behavioral and neurochemical changes in rats during an elevated plus-maze test. J. Neurol. Sci. 2003, 211, 5–14. [Google Scholar] [CrossRef]
- Swanson, R.A.; Morton, M.T.; Tsao-Wu, G.; Savalos, R.A.; Davidson, C.; Sharp, F.R. A semiautomated method for measuring brain infarct volume. J. Cereb. Blood Flow Metab. 1990, 10, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wen, H.; Wang, Q.; Zhang, C.; Jiang, L.; Dou, Z.; Luo, X.; Zeng, J. Exercise training inhibits the Nogo-A/NgR1/Rho-A signals in the cortical peri-infarct area in hypertensive stroke rats. Am. J. Phys. Med. Rehabil. 2015, 94, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Hu, X.Q.; Luo, J.; Li, L.L.; Chen, X.Y.; Huang, R.X.; Pei, Z. Physical exercise improves functional recovery through mitigation of autophagy, attenuation of apoptosis and enhancement of neurogenesis after MCAO in rats. BMC Neurosci. 2013, 14, 46. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Harland, J.; McCulloch, J.; Graham, D.I.; Brown, S.M.; Macrae, I.M. Specific expression of the cell cycle regulation proteins, GADD34 and PCNA, in the peri-infarct zone after focal cerebral ischemia in the rat. Eur. J. Neurosci. 2002, 15, 1929–1936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.S.; Li, J.J.; Xing, S.; Li, C.; Li, Y.; Dang, C.; Fan, Y.; Yu, J.; Pei, Z.; et al. Autophagosomes accumulation is associated with β-amyloid deposits and secondary damage in the thalamus after focal cortical infarction in hypertension rats. J. Neurochem. 2012, 120, 564–573. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Zheng, H.; Zhang, L.; Zhang, Q.; Li, L.; Pei, Z.; Hu, X. High-Frequency Repetitive Transcranial Magnetic Stimulation (rTMS) Improves Functional Recovery by Enhancing Neurogenesis and Activating BDNF/TrkB Signaling in Ischemic Rats. Int. J. Mol. Sci. 2017, 18, 455. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18020455
Luo J, Zheng H, Zhang L, Zhang Q, Li L, Pei Z, Hu X. High-Frequency Repetitive Transcranial Magnetic Stimulation (rTMS) Improves Functional Recovery by Enhancing Neurogenesis and Activating BDNF/TrkB Signaling in Ischemic Rats. International Journal of Molecular Sciences. 2017; 18(2):455. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18020455
Chicago/Turabian StyleLuo, Jing, Haiqing Zheng, Liying Zhang, Qingjie Zhang, Lili Li, Zhong Pei, and Xiquan Hu. 2017. "High-Frequency Repetitive Transcranial Magnetic Stimulation (rTMS) Improves Functional Recovery by Enhancing Neurogenesis and Activating BDNF/TrkB Signaling in Ischemic Rats" International Journal of Molecular Sciences 18, no. 2: 455. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18020455