
Effects of Ranibizumab and Aflibercept on Human Müller Cells and Photoreceptors under Stress Conditions

Abstract
:
1. Introduction
2. Results
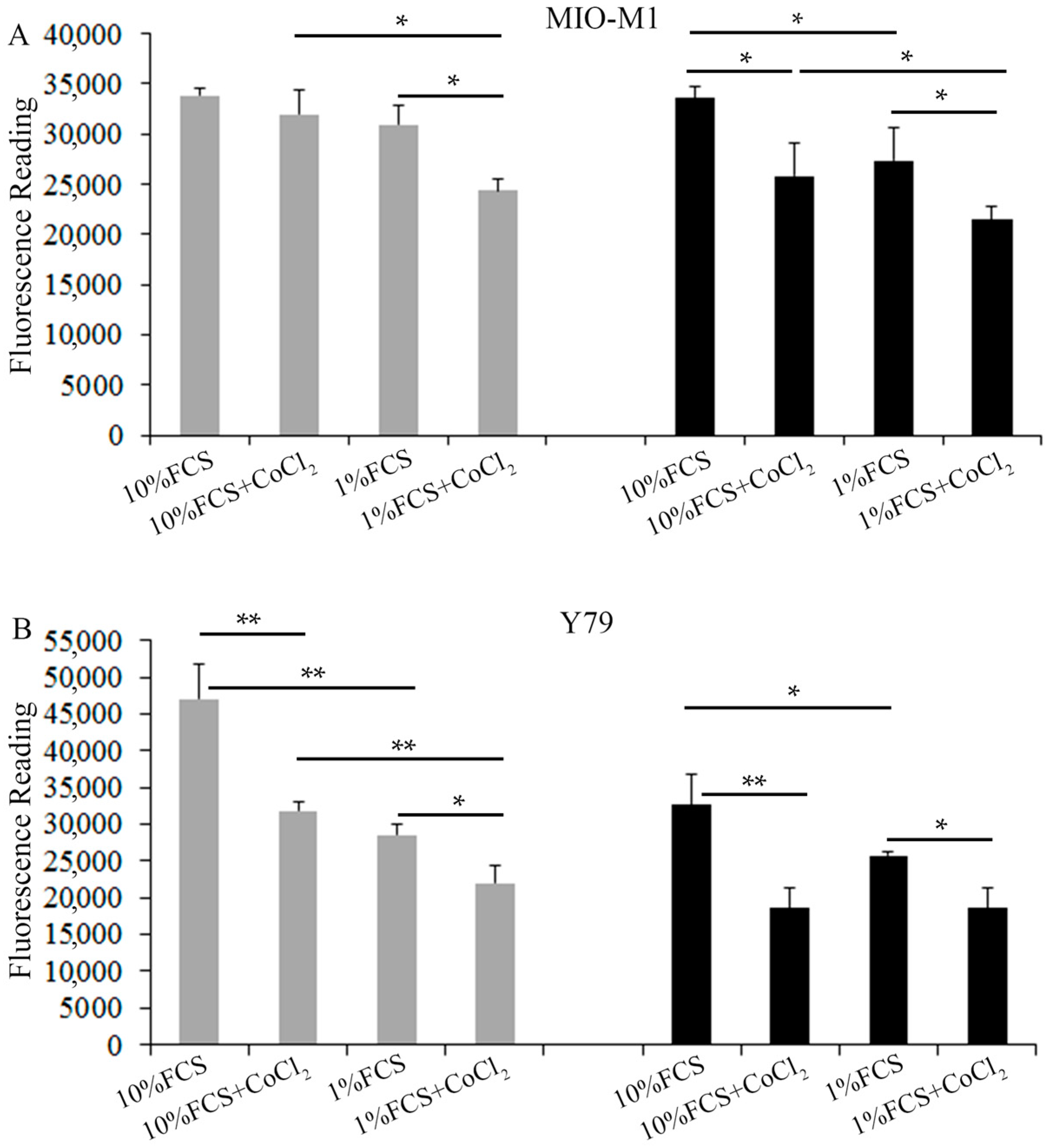
2.1. Effects of Hyperglycemia and Hypoxia on the Survival of Müller Cells and Photoreceptors
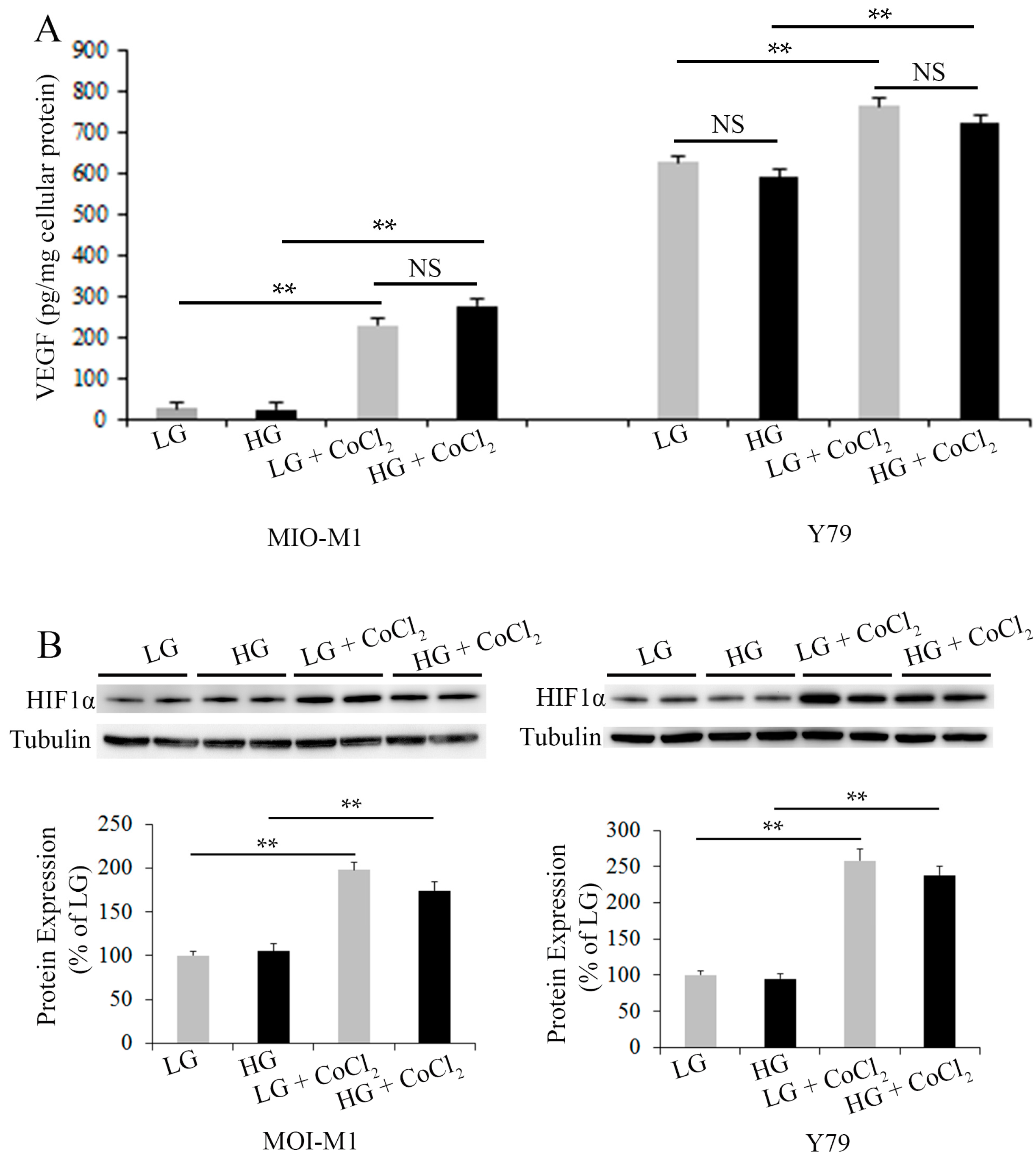
2.2. Effects of Cell Stress on Vascular Endothelial Growth Factor (VEGF)-A Production and Hypoxia-Inducible Factor 1α (HIF1α) Expression in Müller Cells and Photoreceptors
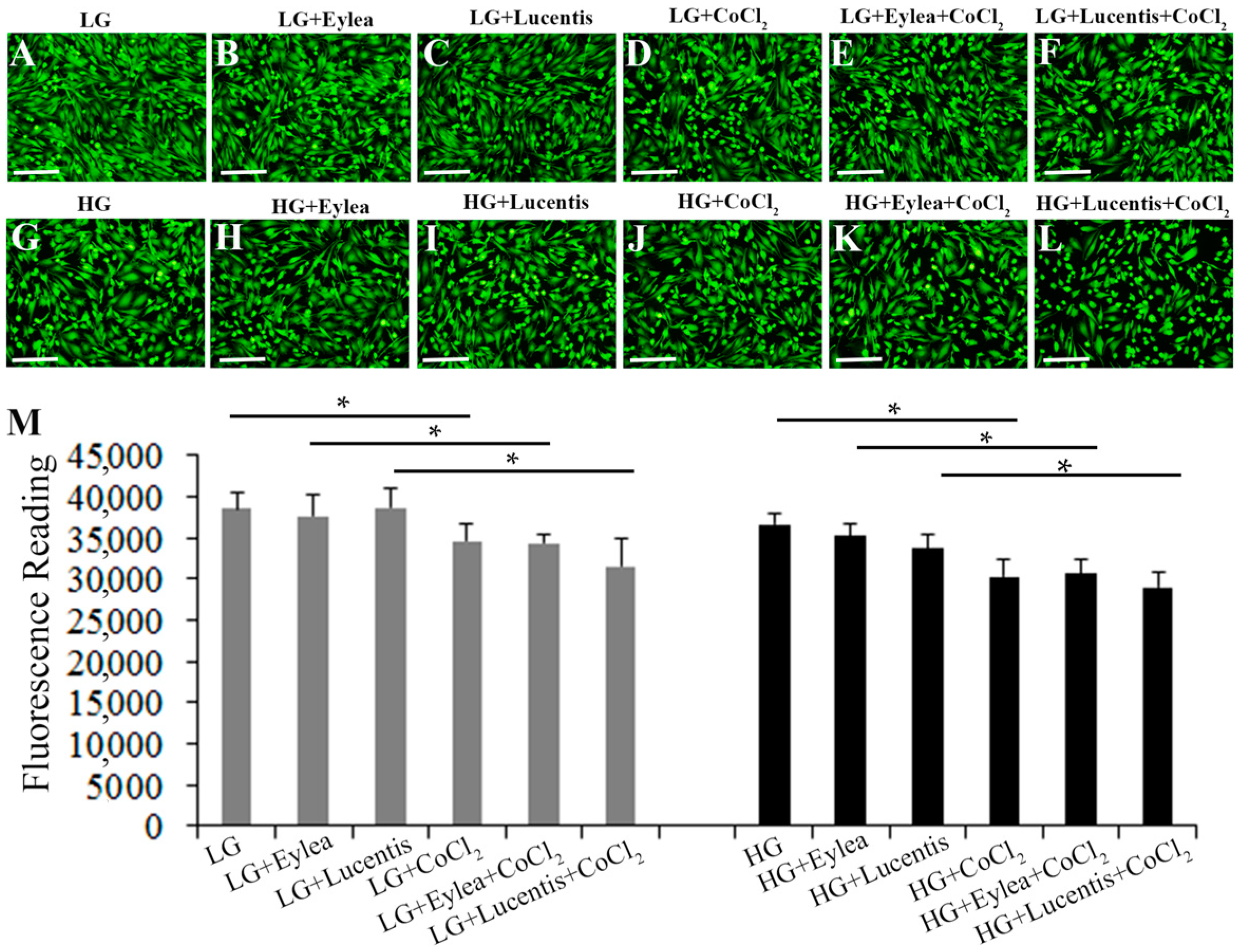
2.3. Effects of Aflibercept and Ranibizumab on Müller Cell Survival
2.4. Effects of Aflibercept and Ranibizumab on Photoreceptor Cell Viability
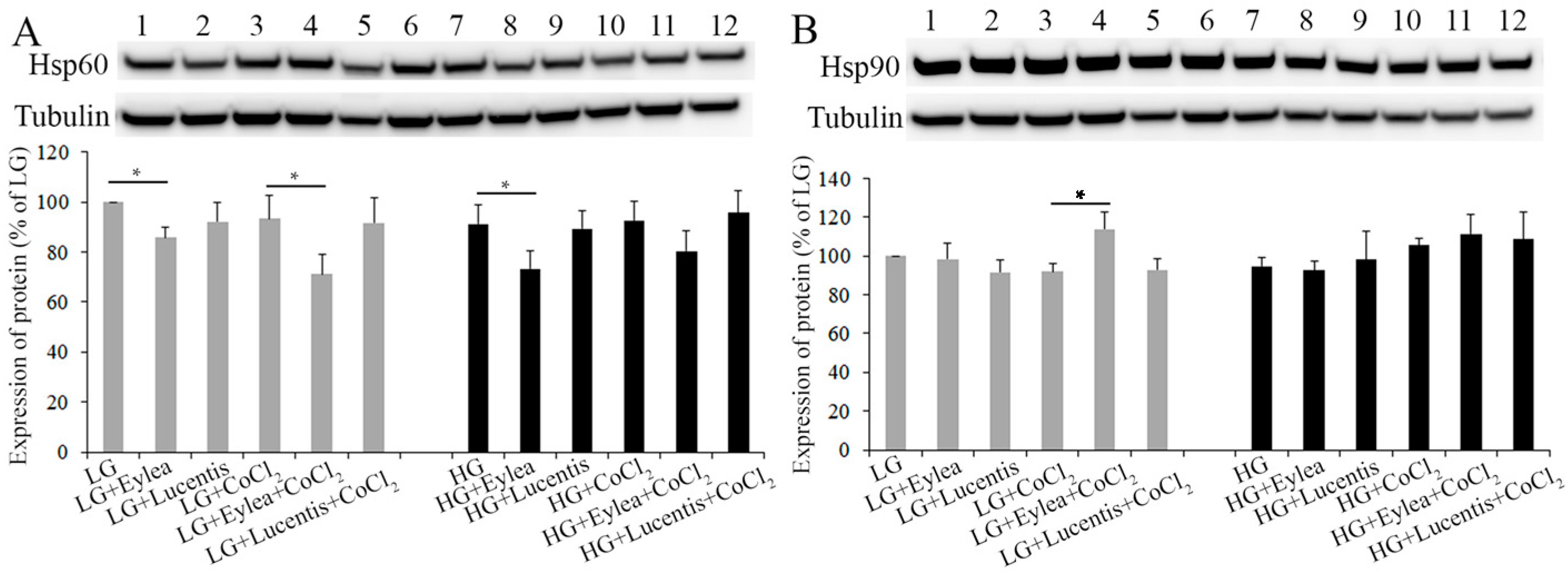
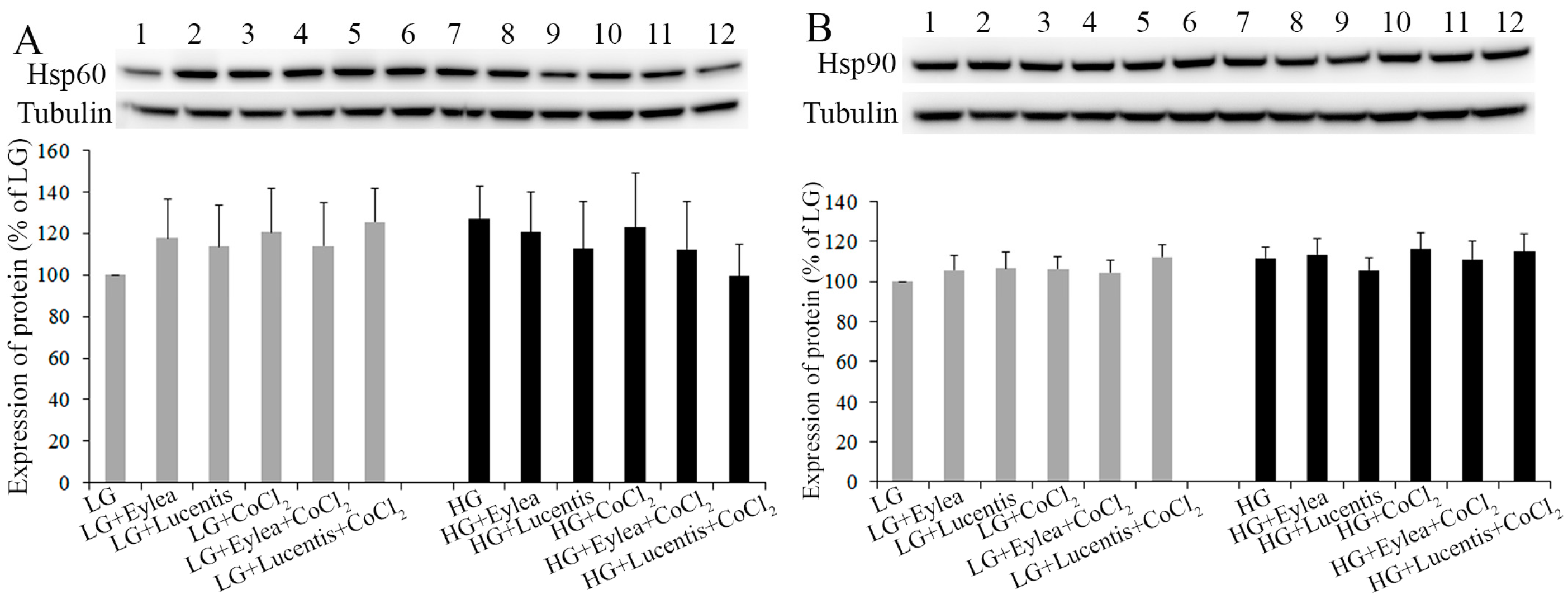
2.5. Effects of Aflibercept and Ranibizumab on Hsp60 and Hsp90 Expression in Müller Cells
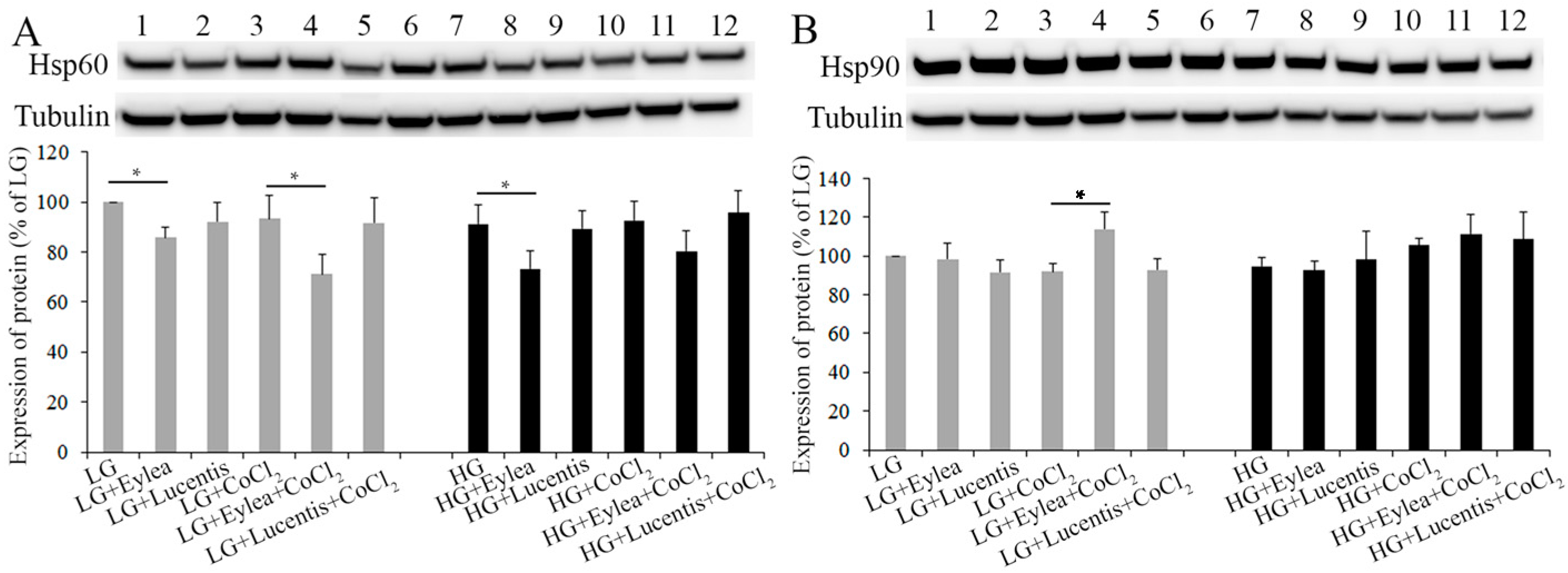
2.6. Effects of Aflibercept and Ranibizumab on Hsp60 and Hsp90 Expression in Photoreceptors
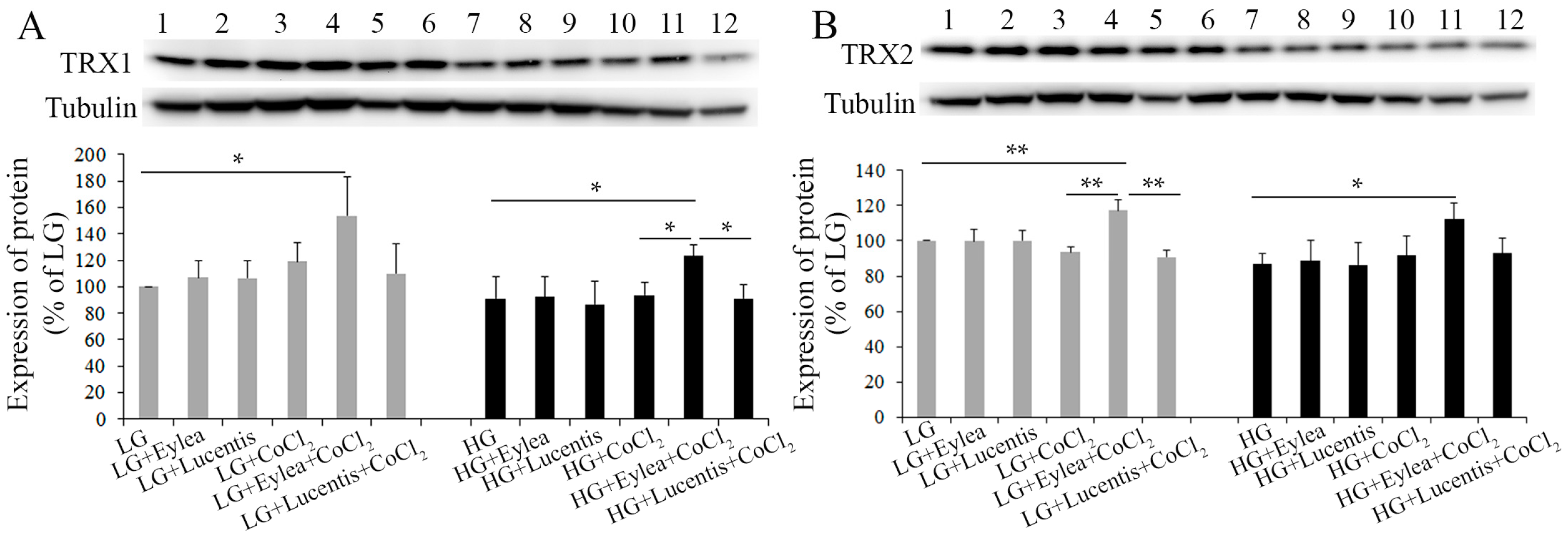
2.7. Effects of Aflibercept and Ranibizumab on Thioredoxin 1 and 2 (TRX1, TRX2) Expression in Photoreceptors
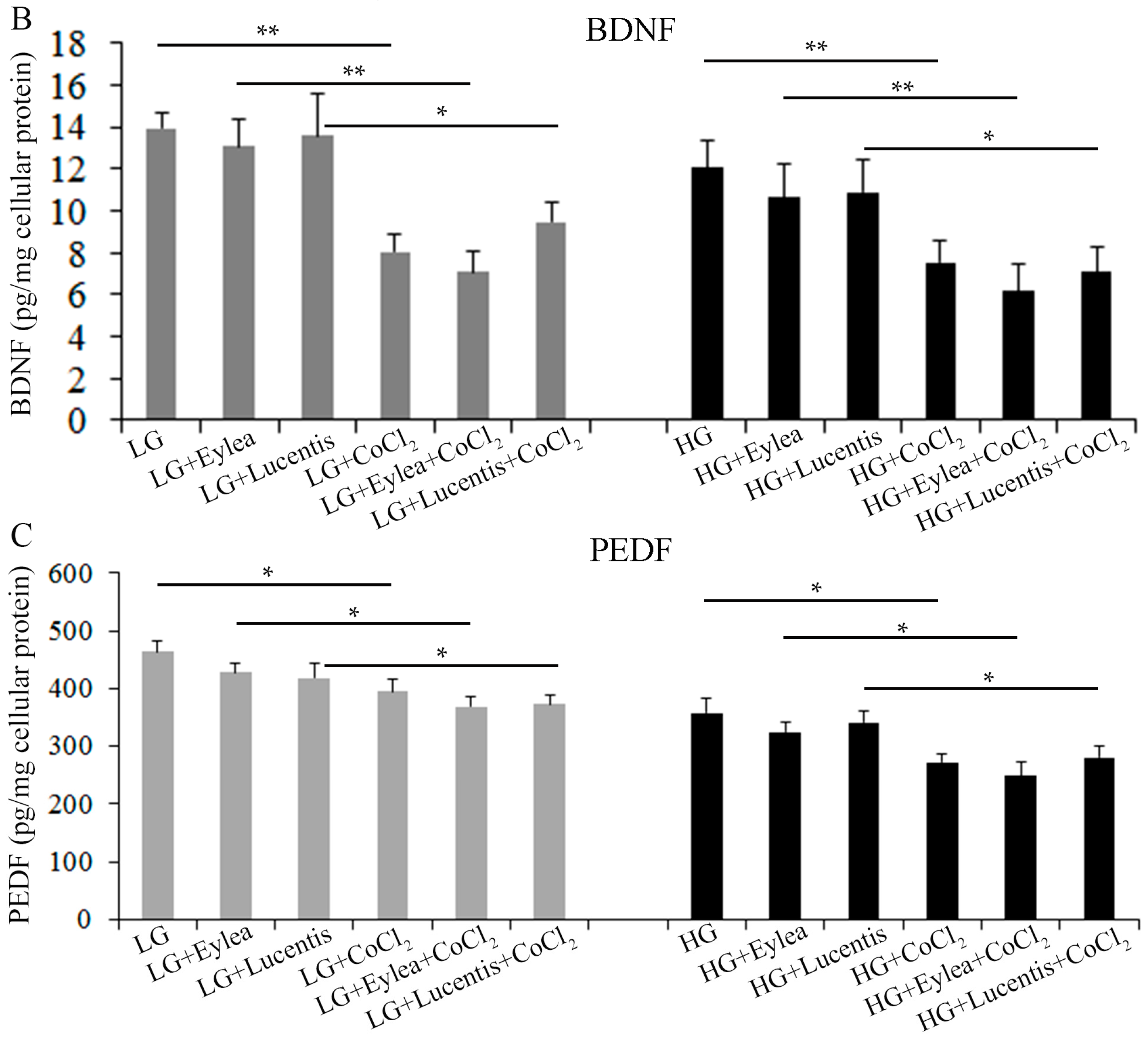
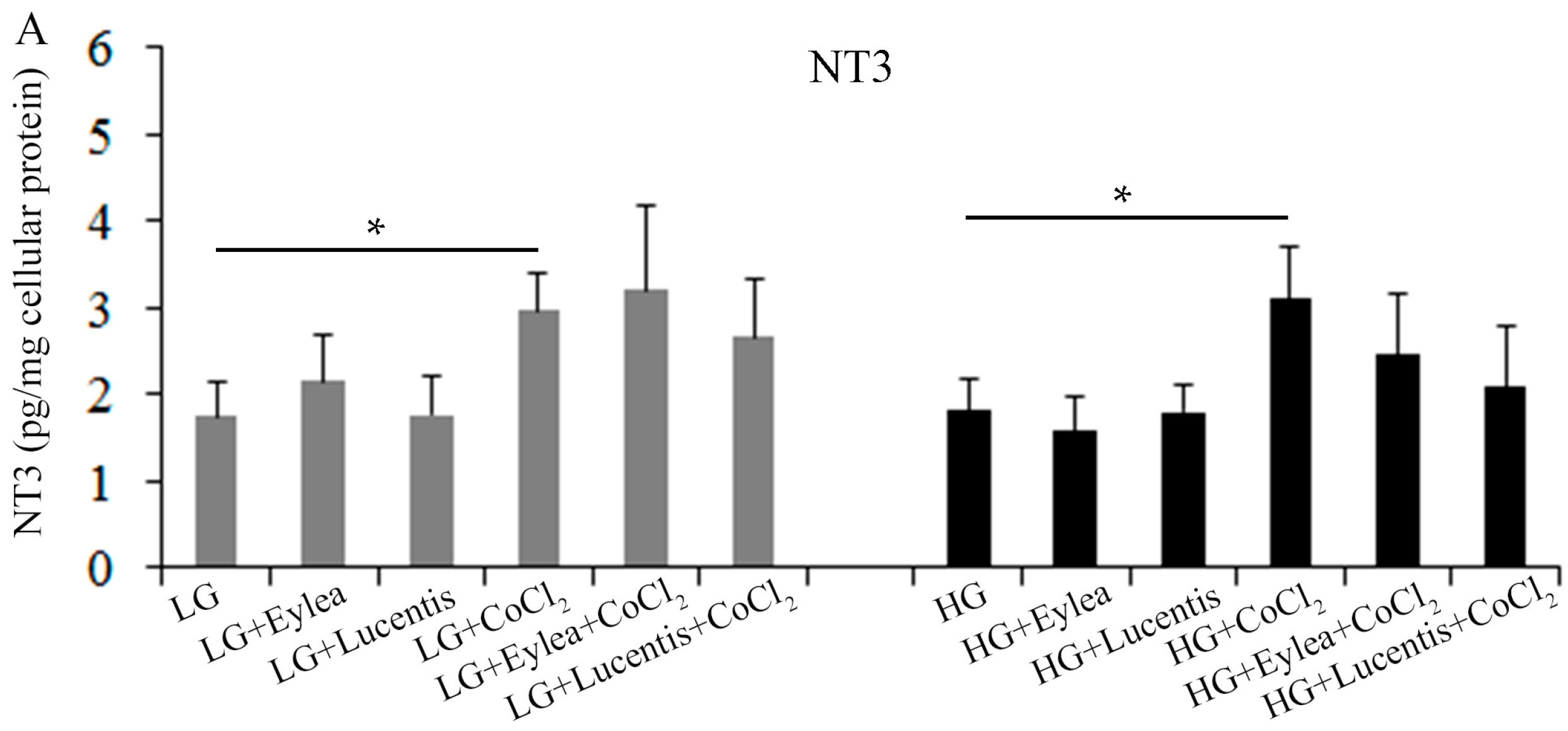
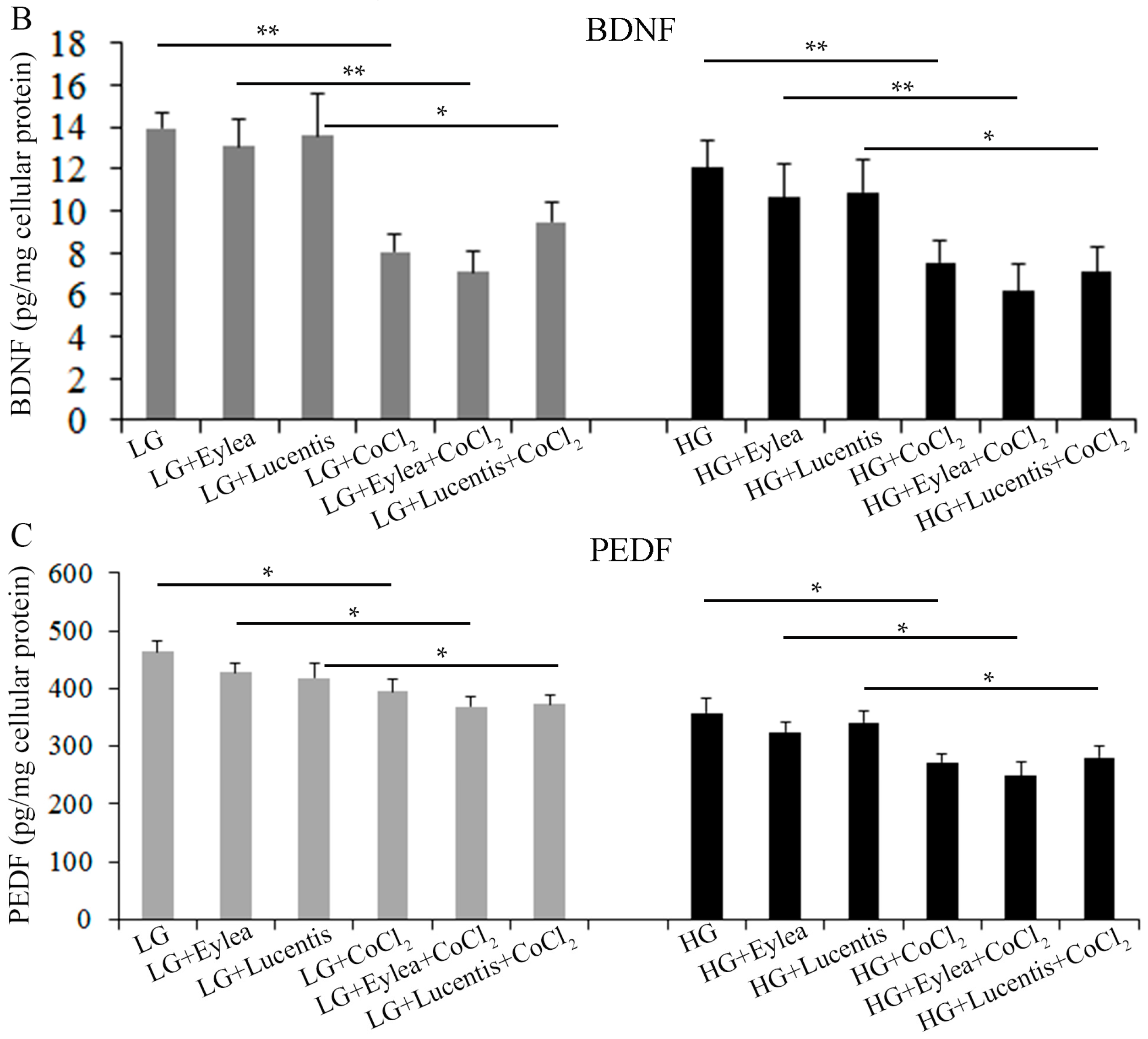
2.8. Effects of Aflibercept and Ranibizumab on Neurotrophic Factors in Müller Cells
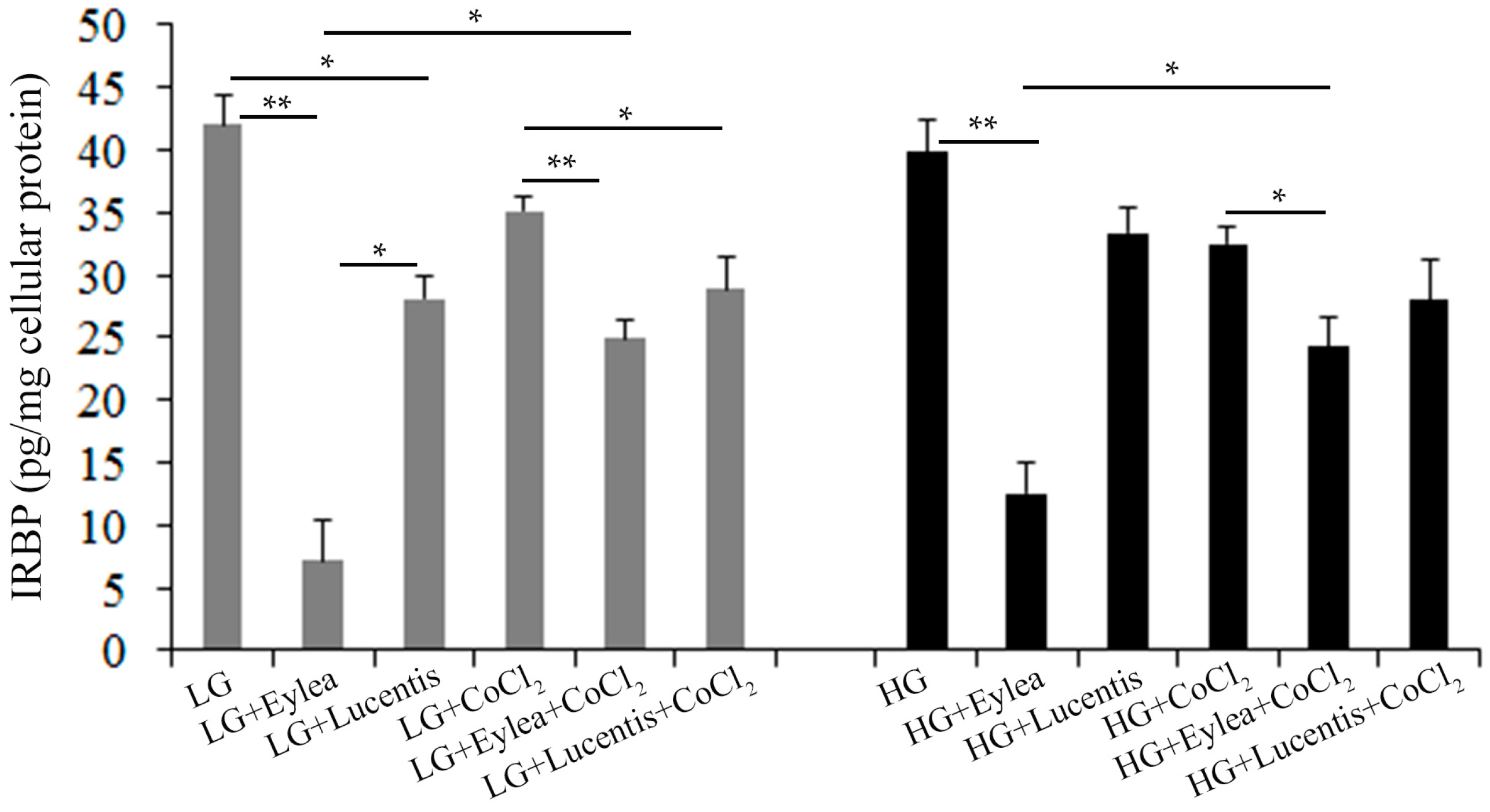
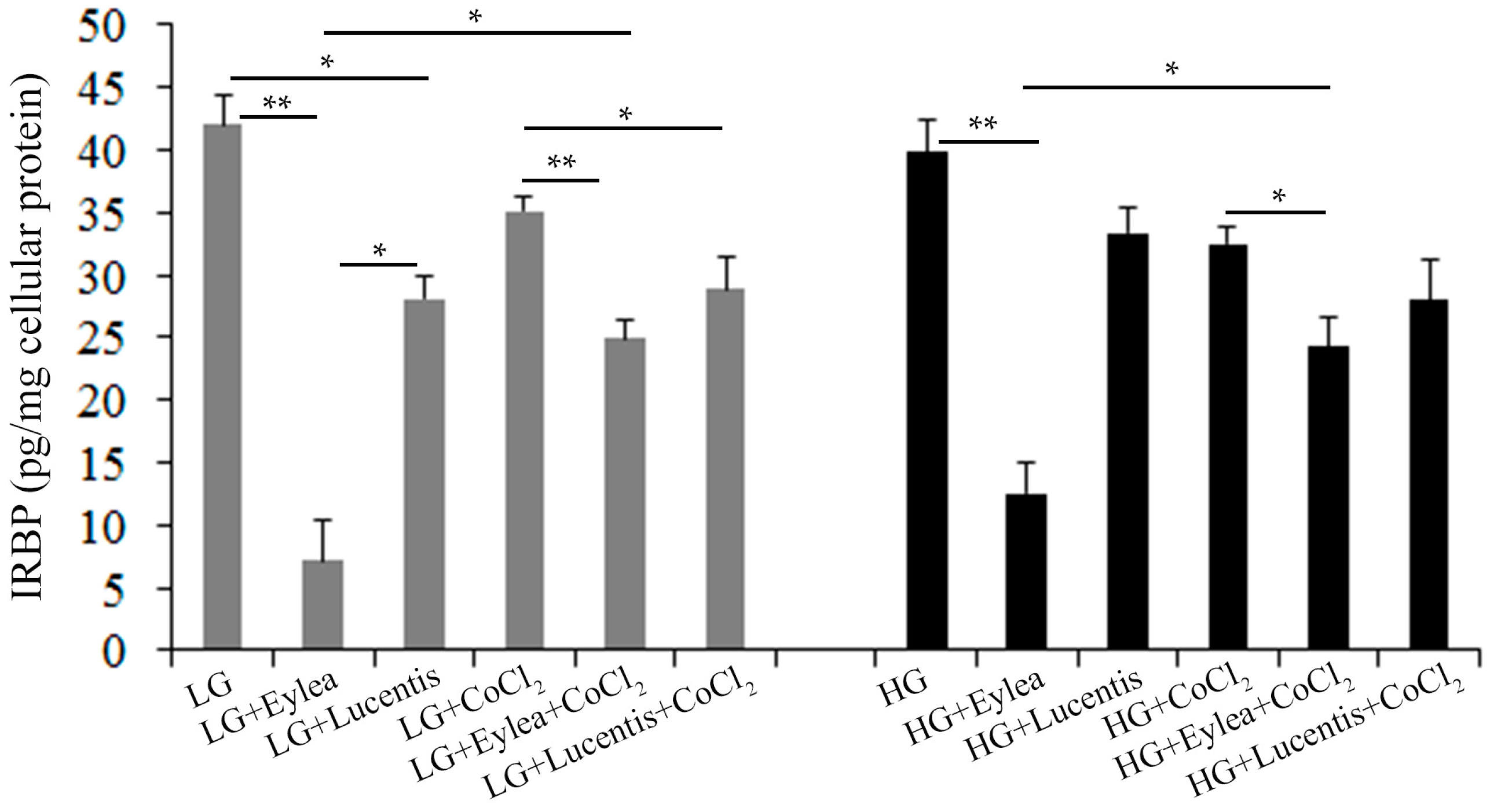
2.9. Effects of Aflibercept and Ranibizumab on Interphotoreceptor Retinoid-Binding Protein (IRBP) Production in Y79 Photoreceptors
3. Discussion
4. Materials and Methods
4.1. Culture of Human Müller Cells and Photoreceptors under Normal and Stressed Conditions
4.2. Anti-VEGF Treatments in Human Müller Cells and Photoreceptors
4.3. Calcein-AM Cell Viability Assay
4.4. Measurements of VEGF-A, NT3, BDNF, PEDF and IRBP in Conditioned Media Using Enzyme-Linked Immunosorbent Assays
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Sene, A.; Chin-Yee, D.; Apte, R.S. Seeing through VEGF: Innate and adaptive immunity in pathological angiogenesis in the eye. Trends Mol. Med. 2015, 21, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Jager, R.D.; Mieler, W.F.; Miller, J.W. Age-related macular degeneration. N. Engl. J. Med. 2008, 358, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Antonetti, D.A.; Klein, R.; Gardner, T.W. Diabetic retinopathy. N. Engl. J. Med. 2012, 366, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, F.; Morescalchi, F.; Duse, S.; Parmeggiani, F.; Gambicorti, E.; Costagliola, C. Aflibercept in wet AMD: Specific role and optimal use. Drug Des. Dev. Ther. 2013, 7, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Saint-Geniez, M.; Kurihara, T.; Sekiyama, E.; Maldonado, A.E.; D’Amore, P.A. An essential role for RPE-derived soluble VEGF in the maintenance of the choriocapillaris. Proc. Natl. Acad. Sci. USA 2009, 106, 18751–18756. [Google Scholar] [CrossRef] [PubMed]
- Marneros, A.G.; Fan, J.; Yokoyama, Y.; Gerber, H.P.; Ferrara, N.; Crouch, R.K.; Olsen, B.R. Vascular endothelial growth factor expression in the retinal pigment epithelium is essential for choriocapillaris development and visual function. Am. J. Pathol. 2005, 167, 1451–1459. [Google Scholar] [CrossRef]
- Byeon, S.H.; Lee, S.C.; Choi, S.H.; Lee, H.K.; Lee, J.H.; Chu, Y.K.; Kwon, O.W. Vascular endothelial growth factor as an autocrine survival factor for retinal pigment epithelial cells under oxidative stress via the VEGF-R2/PI3K/Akt. Investig. Ophthalmoil Vis. Sci. 2010, 51, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Foxton, R.H.; Finkelstein, A.; Vijay, S.; Dahlmann-Noor, A.; Khaw, P.T.; Morgan, J.E.; Shima, D.T.; Ng, Y.S. VEGF-A Is Necessary and sufficient for retinal neuroprotection in models of experimental glaucoma. Am. J. Pathol. 2013, 182, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Brar, V.S.; Sharma, R.K.; Murthy, R.K.; Chalam, K.V. Bevacizumab neutralizes the protective effect of vascular endothelial growth factor on retinal ganglion cells. Mol. Vis. 2010, 16, 1848–1853. [Google Scholar] [PubMed]
- Hombrebueno, J.R.; Ali, I.H.; Xu, H.; Chen, M. Sustained intraocular VEGF neutralization results in retinal neurodegeneration in the Ins2(Akita) diabetic mouse. Sci. Rep. 2015, 5, 18316. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Kim, J.H.; Park, C.K. Neuronal cell death in the inner retina and the influence of vascular endothelial growth factor inhibition in a diabetic rat model. Am. J. Pathol. 2014, 184, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Saint-Geniez, M.; Maharaj, A.S.; Walshe, T.E.; Tucker, B.A.; Sekiyama, E.; Kurihara, T.; Darland, D.C.; Young, M.J.; D’Amore, P.A. Endogenous VEGF is required for visual function: Evidence for a survival role on Müller cells and photoreceptors. PLoS ONE 2008, 3, e3554. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, T.; Westenskow, P.D.; Bravo, S.; Aguilar, E.; Friedlander, M. Targeted deletion of Vegfa in adult mice induces vision loss. J. Clin. Investig. 2012, 122, 4213–4217. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Müller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.M.; Yafai, Y.; Wiedemann, P.; Kuhrt, H.; Wang, Y.S.; Reichenbach, A.; Eichler, W. Hypoxia-induced upregulation of pigment epithelium-derived factor by retinal glial (Müller) cells. J. Neurosci. Res. 2012, 90, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Unterlauft, J.D.; Eichler, W.; Kuhne, K.; Yang, X.M.; Yafai, Y.; Wiedemann, P.; Reichenbach, A.; Claudepierre, T. Pigment epithelium-derived factor released by Müller glial cells exerts neuroprotective effects on retinal ganglion cells. Neurochem. Res. 2012, 37, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Zhu, L.; Lee, S.R.; Chung, S.H.; Gillies, M.C. Involvement of NT3 and P75(NTR) in photoreceptor degeneration following selective Müller cell ablation. J. Neuroinflam. 2013, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Vogler, S.; Hollborn, M.; Berk, B.A.; Pannicke, T.; Seeger, J.; Wiedemann, P.; Reichenbach, A.; Bringmann, A. Ischemic regulation of brain-derived neurotrophic factor-mediated cell volume and TrkB expression in glial (Müller) and bipolar cells of the rat retina. Graefes Arch. Clin. Exp. Ophthalmol. 2016, 254, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Malik, D.; Tarek, M.; Caceres del Carpio, J.; Ramirez, C.; Boyer, D.; Kenney, M.C.; Kuppermann, B.D. Safety profiles of anti-VEGF drugs: Bevacizumab, ranibizumab, aflibercept and ziv-aflibercept on human retinal pigment epithelium cells in culture. Br. J. Ophthalmol. 2014, 98 (Suppl. 1), i11–i16. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Mohle, F.; Roider, J. Intracellular bevacizumab reduces phagocytotic uptake in RPE cells. Graefes Arch. Clin. Exp. Ophthalmol. 2010, 248, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Adelman, R.A.; Rizzolo, L.J. Minimal effects of VEGF and anti-VEGF drugs on the permeability or selectivity of RPE tight junctions. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3216–3225. [Google Scholar] [CrossRef] [PubMed]
- Schnichels, S.; Hagemann, U.; Januschowski, K.; Hofmann, J.; Bartz-Schmidt, K.U.; Szurman, P.; Spitzer, M.S.; Aisenbrey, S. Comparative toxicity and proliferation testing of aflibercept, bevacizumab and ranibizumab on different ocular cells. Br. J. Ophthalmol. 2013, 97, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Ojala, J.; Kaarniranta, K.; Hiltunen, M.; Soininen, H. Hsp90 regulates tau pathology through co-chaperone complexes in Alzheimer’s disease. Prog. Neurobiol. 2011, 93, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Bross, P.; Magnoni, R.; Bie, A.S. Molecular chaperone disorders: Defective Hsp60 in neurodegeneration. Curr. Top. Med. Chem. 2012, 12, 2491–2503. [Google Scholar] [CrossRef] [PubMed]
- Vlamis-Gardikas, A.; Holmgren, A. Thioredoxin and glutaredoxin isoforms. Methods Enzymol. 2002, 347, 286–296. [Google Scholar] [PubMed]
- Seigel, G.M.; Notter, M.F. Differentiation of Y79 retinoblastoma cells induced by succinylated concanavalin A. Cell Growth Differ. 1993, 4, 1–7. [Google Scholar] [PubMed]
- Rehemtulla, A.; Warwar, R.; Kumar, R.; Ji, X.; Zack, D.J.; Swaroop, A. The basic motif-leucine zipper transcription factor Nrl can positively regulate rhodopsin gene expression. Proc. Natl. Acad. Sci. USA 1996, 93, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Craft, C.M.; Xu, J.; Slepak, V.Z.; Zhan-Poe, X.; Zhu, X.; Brown, B.; Lolley, R.N. PhLPs and PhLOPs in the phosducin family of Gβγ binding proteins. Biochemistry 1998, 37, 15758–15772. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Zhu, X.; Craft, C.M. Retinoic acid upregulates cone arrestin expression in retinoblastoma cells through a Cis element in the distal promoter region. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1375–1383. [Google Scholar]
- Garcia-Ramirez, M.; Hernandez, C.; Villarroel, M.; Canals, F.; Alonso, M.A.; Fortuny, R.; Masmiquel, L.; Navarro, A.; Garcia-Arumi, J.; Simo, R. Interphotoreceptor retinoid-binding protein (IRBP) is downregulated at early stages of diabetic retinopathy. Diabetologia 2009, 52, 2633–2641. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, H.; Yu, A.Y.; Della, N.; Ozaki, K.; Luna, J.D.; Yamada, H.; Hackett, S.F.; Okamoto, N.; Zack, D.J.; Semenza, G.L.; et al. Hypoxia inducible factor-1α is increased in ischemic retina: Temporal and spatial correlation with VEGF expression. Investig. Ophthalmol. Vis. Sci. 1999, 40, 182–189. [Google Scholar]
- Bai, Y.; Ma, J.X.; Guo, J.; Wang, J.; Zhu, M.; Chen, Y.; Le, Y.Z. Müller cell-derived VEGF is a significant contributor to retinal neovascularization. J. Pathol. 2009, 219, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Kvanta, A.; Steen, B.; Seregard, S. Expression of vascular endothelial growth factor (VEGF) in retinoblastoma but not in posterior uveal melanoma. Exp. Eye Res. 1996, 63, 511–518. [Google Scholar] [CrossRef] [PubMed]
- De Gooyer, T.E.; Stevenson, K.A.; Humphries, P.; Simpson, D.A.; Gardiner, T.A.; Stitt, A.W. Retinopathy is reduced during experimental diabetes in a mouse model of outer retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5561–5568. [Google Scholar] [CrossRef] [PubMed]
- Arden, G.B.; Sidman, R.L.; Arap, W.; Schlingemann, R.O. Spare the rod and spoil the eye. Br. J. Ophthalmol. 2005, 89, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Arden, G.B. The absence of diabetic retinopathy in patients with retinitis pigmentosa: Implications for pathophysiology and possible treatment. Br. J. Ophthalmol. 2001, 85, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Veenstra, A.; Palczewski, K.; Kern, T.S. Photoreceptor cells are major contributors to diabetes-induced oxidative stress and local inflammation in the retina. Proc. Natl. Acad. Sci. USA 2013, 110, 16586–16591. [Google Scholar] [CrossRef] [PubMed]
- Joyal, J.S.; Sun, Y.; Gantner, M.L.; Shao, Z.; Evans, L.P.; Saba, N.; Fredrick, T.; Burnim, S.; Kim, J.S.; Patel, G.; et al. Retinal lipid and glucose metabolism dictates angiogenesis through the lipid sensor Ffar1. Nat. Med. 2016, 22, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Puddu, A.; Sanguineti, R.; Traverso, C.E.; Viviani, G.L.; Nicolo, M. Response to anti-VEGF-A treatment of retinal pigment epithelial cells in vitro. Eur. J. Ophthalmol. 2016, 26, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, K.; Ng, Y.S.; Zhong, L.C.; Bradley, J.; Schubert, W.; Jo, N.; Akita, J.; Samuelsson, S.J.; Robinson, G.S.; Adamis, A.P.; et al. Vascular endothelial growth factor-A is a survival factor for retinal neurons and a critical neuroprotectant during the adaptive response to ischemic injury. Am. J. Pathol. 2007, 171, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Tahmaz, N.; Dithmer, M.; Richert, E.; Roider, J. Effects of aflibercept on primary RPE cells: Toxicity, wound healing, uptake and phagocytosis. Br. J. Ophthalmol. 2014, 98, 1448–1452. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.K.; Kruse, M.L.; Meyer, T.; Wesch, D.; Kabelitz, D.; Roider, J. Different properties of VEGF-antagonists: Bevacizumab but not Ranibizumab accumulates in RPE cells. Graefes Arch. Clin. Exp. Ophthalmol. 2009, 247, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Inan, U.U.; Avci, B.; Kusbeci, T.; Kaderli, B.; Avci, R.; Temel, S.G. Preclinical safety evaluation of intravitreal injection of full-length humanized vascular endothelial growth factor antibody in rabbit eyes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Avci, B.; Avci, R.; Inan, U.U.; Kaderli, B. Comparative evaluation of apoptotic activity in photoreceptor cells after intravitreal injection of bevacizumab and pegaptanib sodium in rabbits. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3438–3446. [Google Scholar] [CrossRef] [PubMed]
- Januschowski, K.; Schnichels, S.; Hagemann, U.; Koch, V.; Hofmann, J.; Spitzer, M.S.; Bartz-Schmidt, K.U.; Szurman, P.; Luke, M.; Aisenbrey, S. Electrophysiological toxicity testing of VEGF Trap-Eye in an isolated perfused vertebrate retina organ culture model. Acta Ophthalmol. 2014, 92, e305–e311. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Harada, C.; Kohsaka, S.; Wada, E.; Yoshida, K.; Ohno, S.; Mamada, H.; Tanaka, K.; Parada, L.F.; Wada, K. Microglia-Müller glia cell interactions control neurotrophic factor production during light-induced retinal degeneration. J. Neurosci. 2002, 22, 9228–9236. [Google Scholar] [PubMed]
- Hu, L.M.; Luo, Y.; Zhang, J.; Lei, X.; Shen, J.; Wu, Y.; Qin, M.; Unver, Y.B.; Zhong, Y.; Xu, G.T.; et al. EPO reduces reactive gliosis and stimulates neurotrophin expression in Müller cells. Front. Biosci. 2011, 3, 1541–1555. [Google Scholar]
- Fu, S.; Dong, S.; Zhu, M.; Sherry, D.M.; Wang, C.; You, Z.; Haigh, J.J.; Le, Y.Z. Müller glia are a major cellular source of survival signals for retinal neurons in diabetes. Diabetes 2015, 64, 3554–3563. [Google Scholar] [CrossRef] [PubMed]
- Unterlauft, J.D.; Claudepierre, T.; Schmidt, M.; Müller, K.; Yafai, Y.; Wiedemann, P.; Reichenbach, A.; Eichler, W. Enhanced survival of retinal ganglion cells is mediated by Müller glial cell-derived PEDF. Exp. Eye Res. 2014, 127, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Fernandez, F.; Landers, R.A.; Glazebrook, P.A.; Fong, S.L.; Liou, G.I.; Lam, D.M.; Bridges, C.D. An extracellular retinol-binding glycoprotein in the eyes of mutant rats with retinal dystrophy: Development, localization, and biosynthesis. J. Cell Biol. 1984, 99, 2092–2098. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Sawada, Y.; Yoshitomi, T. Structure and function of the interphotoreceptor matrix surrounding retinal photoreceptor cells. Exp. Eye Res. 2015, 133, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Den Hollander, A.I.; McGee, T.L.; Ziviello, C.; Banfi, S.; Dryja, T.P.; Gonzalez-Fernandez, F.; Ghosh, D.; Berson, E.L. A homozygous missense mutation in the IRBP gene (RBP3) associated with autosomal recessive retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1864–1872. [Google Scholar] [CrossRef] [PubMed]
- Sands, M.; Howell, K.; Costello, C.M.; McLoughlin, P. Placenta growth factor and vascular endothelial growth factor B expression in the hypoxic lung. Respir. Res. 2011, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Balaiya, S.; Ferguson, L.R.; Chalam, K.V. Evaluation of sirtuin role in neuroprotection of retinal ganglion cells in hypoxia. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4315–4322. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lafleur, J.; Mwaikambo, B.R.; Zhu, T.; Gagnon, C.; Chemtob, S.; Di Polo, A.; Hardy, P. The role of lysophosphatidic acid receptor (LPA1) in the oxygen-induced retinal ganglion cell degeneration. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Hilliard, G.; Ferguson, T.; Millhorn, D.E. Cobalt inhibits the interaction between hypoxia-inducible factor-α and von Hippel-Lindau protein by direct binding to hypoxia-inducible factor-α. J. Biol. Chem. 2003, 278, 15911–15916. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Li, S.; Chung, S.H.; Gillies, M.C. Retinal vascular changes after glial disruption in rats. J. Neurosci. Res. 2010, 88, 1485–1499. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Zhang, J.; Chung, S.H.; Hu, Y.; Ma, Z.; Gillies, M.C. Submacular DL-α-aminoadipic acid eradicates primate photoreceptors but does not affect luteal pigment or the retinal vasculature. Investig. Ophthalmol. Vis. Sci. 2011, 52, 119–127. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Glucose (LG/HG) | CoCl2 (200 µM) | Aflibercept (0.5 mg/mL) | Ranibizumab (0.125 mg/mL) |
|---|---|---|---|---|
| 1 | LG | − | − | − |
| 2 | LG | − | + | − |
| 3 | LG | − | − | + |
| 4 | LG | + | − | − |
| 5 | LG | + | + | − |
| 6 | LG | + | − | + |
| 7 | HG | − | − | − |
| 8 | HG | − | + | − |
| 9 | HG | − | − | + |
| 10 | HG | + | − | − |
| 11 | HG | + | + | − |
| 12 | HG | + | − | + |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, W.; Yau, B.; Lee, S.-R.; Zhu, L.; Yam, M.; Gillies, M.C. Effects of Ranibizumab and Aflibercept on Human Müller Cells and Photoreceptors under Stress Conditions. Int. J. Mol. Sci. 2017, 18, 533. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18030533
Shen W, Yau B, Lee S-R, Zhu L, Yam M, Gillies MC. Effects of Ranibizumab and Aflibercept on Human Müller Cells and Photoreceptors under Stress Conditions. International Journal of Molecular Sciences. 2017; 18(3):533. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18030533
Chicago/Turabian StyleShen, Weiyong, Belinda Yau, So-Ra Lee, Ling Zhu, Michelle Yam, and Mark C. Gillies. 2017. "Effects of Ranibizumab and Aflibercept on Human Müller Cells and Photoreceptors under Stress Conditions" International Journal of Molecular Sciences 18, no. 3: 533. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18030533