
Comparison of the In Vivo Biotransformation of Two Emerging Estrogenic Contaminants, BP2 and BPS, in Zebrafish Embryos and Adults


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Radioactivity Metabolic Balance
2.1.1. Exposure of Zebrafish Larvae to Bisphenol S (BPS) and Benzophenone-2 (BP2)
2.1.2. Exposure of Adult Zebrafish to BPS and BP2
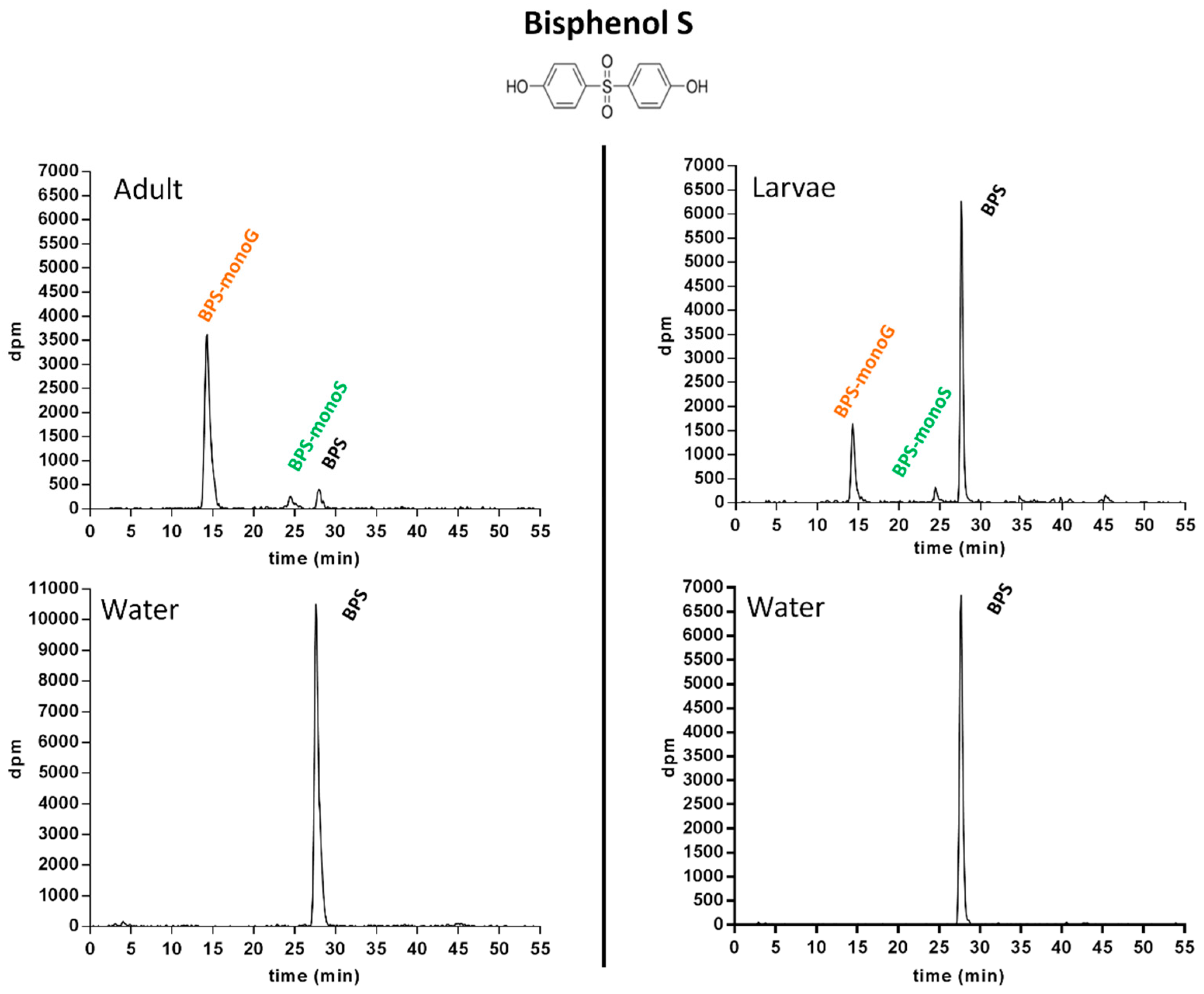
2.1.3. Metabolic Profiling in Zebrafish Adults and Larvae Exposed to BPS
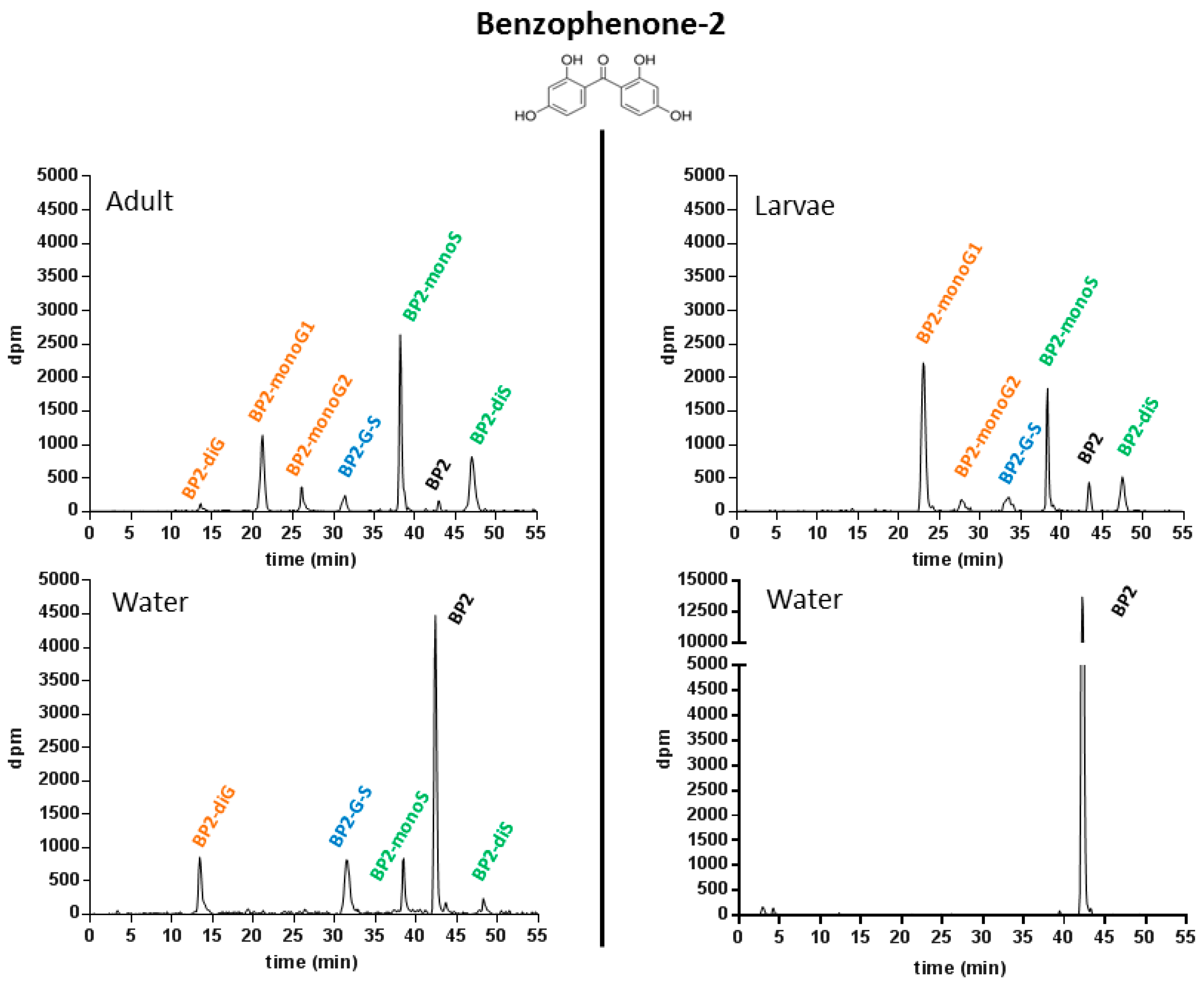
2.1.4. Metabolic Profiling in Zebrafish Adults and Larvae Exposed to BP2
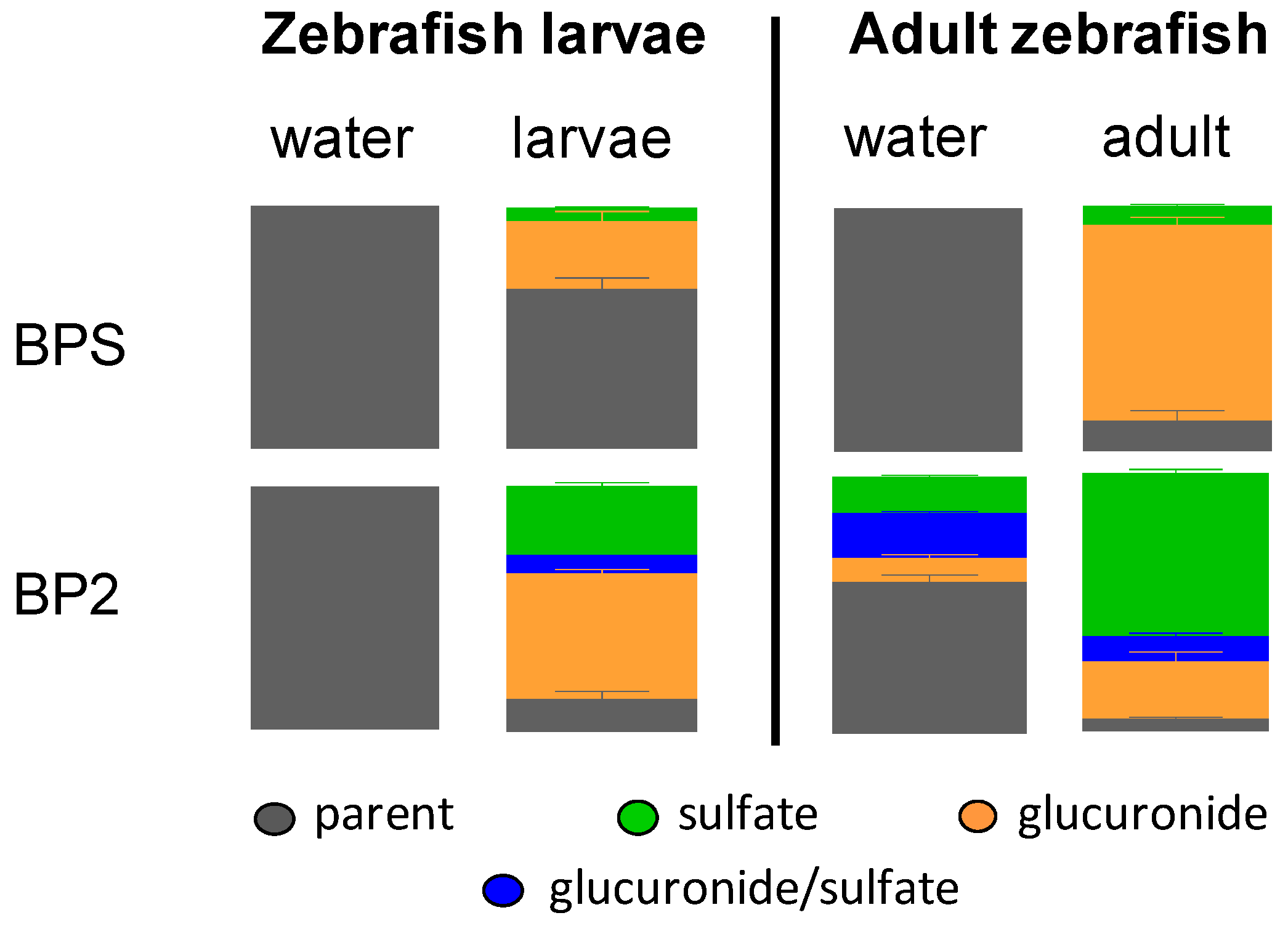
2.1.5. Pattern of BPS and BP2 Biotransformation in Zebrafish Adults and Larvae
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Zebrafish Housing
4.3. Biotransformation Studies in Zebrafish Larvae
4.4. Biotransformation Studies in Adult Zebrafish
4.4.1. Exposure
4.4.2. Samples Preparation for Radio-HPLC Profiling
4.5. Radio-HPLC Profiling
4.6. Enzymatic Hydrolyses
4.7. Radioactivity Quantification in Adult Zebrafish
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ankley, G.T.; Bencic, D.C.; Breen, M.S.; Collette, T.W.; Conolly, R.B.; Denslow, N.D.; Edwards, S.W.; Ekman, D.R.; Garcia-Reyero, N.; Jensen, K.M.; et al. Endocrine disrupting chemicals in fish: Developing exposure indicators and predictive models of effects based on mechanism of action. Aquat. Toxicol. 2009, 92, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Colborn, T.; vom Saal, F.S.; Soto, A.M. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspect. 1993, 101, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, A.K.; Rider, C.V.; Blystone, C.R.; Wilson, V.S.; Hartig, P.C.; Ankley, G.T.; Foster, P.M.; Gray, C.L.; Gray, L.E. Fifteen years after “Wingspread”—Environmental endocrine disrupters and human and wildlife health: Where we are today and where we need to go. Toxicol. Sci. 2008, 105, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Segner, H. Zebrafish (Danio rerio) as a model organism for investigating endocrine disruption. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Cosnefroy, A.; Brion, F.; Maillot-Marechal, E.; Porcher, J.M.; Pakdel, F.; Balaguer, P.; Ait-Aissa, S. Selective activation of zebrafish estrogen receptor subtypes by chemicals by using stable reporter gene assay developed in a zebrafish liver cell line. Toxicol. Sci. 2012, 125, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Brion, F.; le Page, Y.; Piccini, B.; Cardoso, O.; Tong, S.K.; Chung, B.C.; Kah, O. Screening estrogenic activities of chemicals or mixtures in vivo using transgenic (cyp19a1b-GFP) zebrafish embryos. PLoS ONE 2012, 7, e36069. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.N.; Laws, S.C.; Willett, K.; Schmieder, P.; Odum, J.; Bovee, T.F. In vitro metabolism and bioavailability tests for endocrine active substances: What is needed next for regulatory purposes? ALTEX 2013, 30, 331–351. [Google Scholar] [CrossRef] [PubMed]
- Le Fol, V.; Ait-Aissa, S.; Cabaton, N.; Dolo, L.; Grimaldi, M.; Balaguer, P.; Perdu, E.; Debrauwer, L.; Brion, F.; Zalko, D. Cell-specific biotransformation of benzophenone-2 and bisphenol-S in zebrafish and human in vitro models used for toxicity and estrogenicity screening. Environ. Sci. Technol. 2015, 49, 3860–3868. [Google Scholar] [CrossRef] [PubMed]
- Riu, A.; le Maire, A.; Grimaldi, M.; Audebert, M.; Hillenweck, A.; Bourguet, W.; Balaguer, P.; Zalko, D. Characterization of novel ligands of ER α, ER β, and PPAR γ: The case of halogenated bisphenol A and their conjugated metabolites. Toxicol. Sci. 2011, 122, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Zalko, D.; Soto, A.M.; Dolo, L.; Dorio, C.; Rathahao, E.; Debrauwer, L.; Faure, R.; Cravedi, J.P. Biotransformations of bisphenol A in a mammalian model: Answers and new questions raised by low-dose metabolic fate studies in pregnant CD1 mice. Environ. Health. Perspect. 2003, 111, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Cosnefroy, A.; Brion, F.; Guillet, B.; Laville, N.; Porcher, J.M.; Balaguer, P.; Ait-Aissa, S. A stable fish reporter cell line to study estrogen receptor transactivation by environmental (xeno)estrogens. Toxicol. In Vitro 2009, 23, 1450–1454. [Google Scholar] [CrossRef] [PubMed]
- Weisbrod, C.J.; Kunz, P.Y.; Zenker, A.K.; Fent, K. Effects of the UV filter benzophenone-2 on reproduction in fish. Toxicol. Appl. Pharmacol. 2007, 225, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.H.; Grantham, E.C.; Liu, B.; Macapagal, R.; Willingham, E.; Baskin, L.S. In utero exposure to benzophenone-2 causes hypospadias through an estrogen receptor dependent mechanism. J. Urol. 2007, 178, 1637–1642. [Google Scholar] [CrossRef] [PubMed]
- Molina-Molina, J.M.; Escande, A.; Pillon, A.; Gomez, E.; Pakdel, F.; Cavailles, V.; Olea, N.; Ait-Aissa, S.; Balaguer, P. Profiling of benzophenone derivatives using fish and human estrogen receptor-specific in vitro bioassays. Toxicol. Appl. Pharmacol. 2008, 232, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Hong, S.; Kho, Y.; Choi, K. Effects of bisphenol S exposure on endocrine functions and reproduction of zebrafish. Environ. Sci. Technol. 2013, 47, 8793–8800. [Google Scholar] [CrossRef] [PubMed]
- Molina-Molina, J.M.; Amaya, E.; Grimaldi, M.; Saenz, J.M.; Real, M.; Fernandez, M.F.; Balaguer, P.; Olea, N. In vitro study on the agonistic and antagonistic activities of bisphenol-S and other bisphenol-A congeners and derivatives via nuclear receptors. Toxicol. Appl. Pharmacol. 2013, 272, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Naderi, M.; Wong, M.Y.; Gholami, F. Developmental exposure of zebrafish (Danio rerio) to bisphenol-S impairs subsequent reproduction potential and hormonal balance in adults. Aquat. Toxicol. 2014, 148, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Embry, M.R.; Belanger, S.E.; Braunbeck, T.A.; Galay-Burgos, M.; Halder, M.; Hinton, D.E.; Leonard, M.A.; Lillicrap, A.; Norberg-King, T.; Whale, G. The fish embryo toxicity test as an animal alternative method in hazard and risk assessment and scientific research. Aquat. Toxicol. 2010, 97, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Strahle, U.; Scholz, S.; Geisler, R.; Greiner, P.; Hollert, H.; Rastegar, S.; Schumacher, A.; Selderslaghs, I.; Weiss, C.; Witters, H.; et al. Zebrafish embryos as an alternative to animal experiments—A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 2012, 33, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Arnot, J.A.; Mackay, D.; Parkerton, T.E.; Bonnell, M. A database of fish biotransformation rates for organic chemicals. Environ. Toxicol. Chem. 2008, 27, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Cravedi, J.P. Role of biotransformation in the fate and toxicity of chemicals: Consequences for the assessment of residues in fish. Rev. Med. Vet. 2002, 153, 419–424. [Google Scholar]
- Braunig, J.; Schiwy, S.; Broedel, O.; Muller, Y.; Frohme, M.; Hollert, H.; Keiter, S.H. Time-dependent expression and activity of cytochrome P450 1s in early life-stages of the zebrafish (Danio rerio). Environ. Sci. Poll. Res. 2015, 22, 16319–16328. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J.V.; McArthur, A.G.; Kubota, A.; Zanette, J.; Parente, T.; Jonsson, M.E.; Nelson, D.R.; Stegeman, J.J. Identification and developmental expression of the full complement of cytochrome P450 genes in zebrafish. BMC Genom. 2010, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.; Cavanaugh, K.; Verbueken, E.; Pype, C.; Casteleyn, C.; van Ginneken, C.; van Cruchten, S. Xenobiotic metabolism in the zebrafish: A review of the spatiotemporal distribution, modulation and activity of cytochrome P450 families 1 to 3. J. Toxicol. Sci. 2016, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Verbueken, E.; Alsop, D.; Saad, M.A.; Pype, C.; van Peer, E.M.; Casteleyn, C.R.; van Ginneken, C.J.; Wilson, J.; van Cruchten, S.J. In vitro biotransformation of two human CYP3A probe substrates and their inhibition during early zebrafish development. Int. J. Mol. Sci. 2017, 18, 217. [Google Scholar] [CrossRef] [PubMed]
- Brox, S.; Seiwert, B.; Haase, N.; Kuster, E.; Reemtsma, T. Metabolism of clofibric acid in zebrafish embryos (Danio rerio) as determined by liquid chromatography-high resolution-mass spectrometry. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 185–186, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Brox, S.; Seiwert, B.; Kuster, E.; Reemtsma, T. Toxicokinetics of polar chemicals in zebrafish embryo (Danio rerio): Influence of physicochemical properties and of biological processes. Environ. Sci. Technol. 2016, 50, 10264–10272. [Google Scholar] [CrossRef] [PubMed]
- Gibert, Y.; Sassi-Messai, S.; Fini, J.B.; Bernard, L.; Zalko, D.; Cravedi, J.P.; Balaguer, P.; Andersson-Lendahl, M.; Demeneix, B.; Laudet, V. Bisphenol A induces otolith malformations during vertebrate embryogenesis. BMC Dev. Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.N.; de Jong-Curtain, T.A.; Mawdsley, D.J.; White, S.J.; Shin, J.; Appel, B.; Dong, P.D.; Stainier, D.Y.; Heath, J.K. Formation of the digestive system in zebrafish: III. Intestinal epithelium morphogenesis. Dev. Biol. 2005, 286, 114–135. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, H.; Hill, A. Admetox in zebrafish. Drug Discov. Today Dis. Models 2013, 10, e31–e35. [Google Scholar] [CrossRef]
- Kais, B.; Schneider, K.E.; Keiter, S.; Henn, K.; Ackermann, C.; Braunbeck, T. DMSO modifies the permeability of the zebrafish (Danio rerio) chorion-implications for the fish embryo test (FET). Aquat. Toxicol. 2013, 140–141, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Schlecht, C.; Klammer, H.; Frauendorf, H.; Wuttke, W.; Jarry, H. Pharmacokinetics and metabolism of benzophenone 2 in the rat. Toxicology 2008, 245, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Alderton, W.; Berghmans, S.; Butler, P.; Chassaing, H.; Fleming, A.; Golder, Z.; Richards, F.; Gardner, I. Accumulation and metabolism of drugs and CYP probe substrates in zebrafish larvae. Xenobiotica 2010, 40, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, H.; Wu, Q. Characterization of the zebrafish ugt repertoire reveals a new class of drug-metabolizing UDP glucuronosyltransferases. Mol. Pharmacol. 2014, 86, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Kurogi, K.; Liu, T.A.; Sakakibara, Y.; Suiko, M.; Liu, M.C. The use of zebrafish as a model system for investigating the role of the SULTS in the metabolism of endogenous compounds and xenobiotics. Drug Metab. Rev. 2013, 45, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.A.; Bhuiyan, S.; Liu, M.Y.; Sugahara, T.; Sakakibara, Y.; Suiko, M.; Yasuda, S.; Kakuta, Y.; Kimura, M.; Williams, F.E.; et al. Zebrafish as a model for the study of the phase II cytosolic sulfotransferases. Curr. Drug Metab. 2010, 11, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Kumar, A.P.; Liu, M.Y.; Sakakibara, Y.; Suiko, M.; Chen, L.; Liu, M.C. Identification of a novel thyroid hormone-sulfating cytosolic sulfotransferase, SULT1 ST5, from zebrafish. FEBS J. 2005, 272, 3828–3837. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Liu, M.Y.; Yang, Y.S.; Snow, R.; Takahashi, S.; Liu, M.C. Identification of novel hydroxysteroid-sulfating cytosolic SULTS, SULT2 ST2 and SULT2 ST3, from zebrafish: Cloning, expression, characterization, and developmental expression. Arch. Biochem. Biophys. 2006, 455, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Yasuda, S.; Williams, F.E.; Liu, M.Y.; Sakakibara, Y.; Bhuiyan, S.; Snow, R.; Carter, G.; Liu, M.C. Characterization and ontogenic study of novel steroid-sulfating SULT3 sulfotransferases from zebrafish. Mol. Cell. Endocrinol. 2008, 294, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Busquet, F.; Strecker, R.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T.; Carr, G.J.; Cenijn, P.; Fochtman, P.; Gourmelon, A.; Hubler, N.; et al. OECD validation study to assess intra- and inter-laboratory reproducibility of the zebrafish embryo toxicity test for acute aquatic toxicity testing. Regul. Toxicol. Pharmacol. 2014, 69, 496–511. [Google Scholar] [CrossRef] [PubMed]
- Massei, R.; Vogs, C.; Renner, P.; Altenburger, R.; Scholz, S. Differential sensitivity in embryonic stages of the zebrafish (Danio rerio): The role of toxicokinetics for stage-specific susceptibility for azinphos-methyl lethal effects. Aquat. Toxicol. 2015, 166, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Le Fol, V.; Sonavane, M.; Ait-Aissa, S.; Piccini, B.; Maillot-Marechal, E.; Palluel, O.; Zalko, D.; Brion, F. An integrated in vitro and in vivo approach to assess the estrogenic activity of bisphenol A, bisphenol S and bisphenol F in zebrafish. Ecotoxicol. Environ. Saf. 2017. submitted. [Google Scholar]
- Cosnefroy, A. In Vitro and In Vivo Integrated Assessment of Estrogenic Effects of Environmental Substances in Zebrafish. Ph.D. Thesis, Museum d’Histoire Naturelle, Paris, France, 2010; p. 155. [Google Scholar]
- Pery, A.R.; Devillers, J.; Brochot, C.; Mombelli, E.; Palluel, O.; Piccini, B.; Brion, F.; Beaudouin, R. A physiologically based toxicokinetic model for the zebrafish Danio rerio. Environ. Sci. Technol. 2014, 48, 781–790. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Fol, V.; Brion, F.; Hillenweck, A.; Perdu, E.; Bruel, S.; Aït-Aïssa, S.; Cravedi, J.-P.; Zalko, D. Comparison of the In Vivo Biotransformation of Two Emerging Estrogenic Contaminants, BP2 and BPS, in Zebrafish Embryos and Adults. Int. J. Mol. Sci. 2017, 18, 704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040704
Le Fol V, Brion F, Hillenweck A, Perdu E, Bruel S, Aït-Aïssa S, Cravedi J-P, Zalko D. Comparison of the In Vivo Biotransformation of Two Emerging Estrogenic Contaminants, BP2 and BPS, in Zebrafish Embryos and Adults. International Journal of Molecular Sciences. 2017; 18(4):704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040704
Chicago/Turabian StyleLe Fol, Vincent, François Brion, Anne Hillenweck, Elisabeth Perdu, Sandrine Bruel, Selim Aït-Aïssa, Jean-Pierre Cravedi, and Daniel Zalko. 2017. "Comparison of the In Vivo Biotransformation of Two Emerging Estrogenic Contaminants, BP2 and BPS, in Zebrafish Embryos and Adults" International Journal of Molecular Sciences 18, no. 4: 704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040704