
Chemopreventive Strategies for Inflammation-Related Carcinogenesis: Current Status and Future Direction


Abstract
:1. Introduction
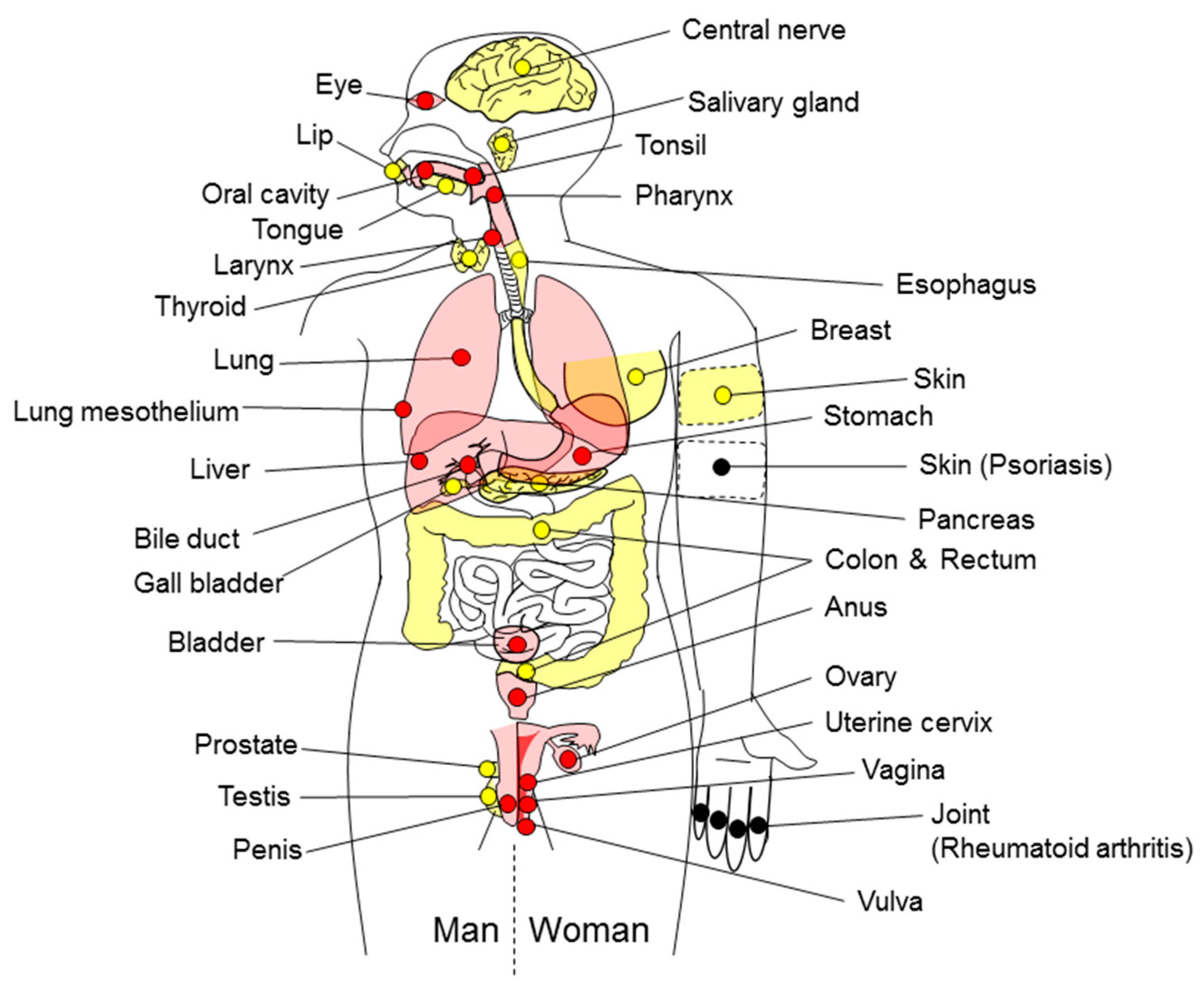
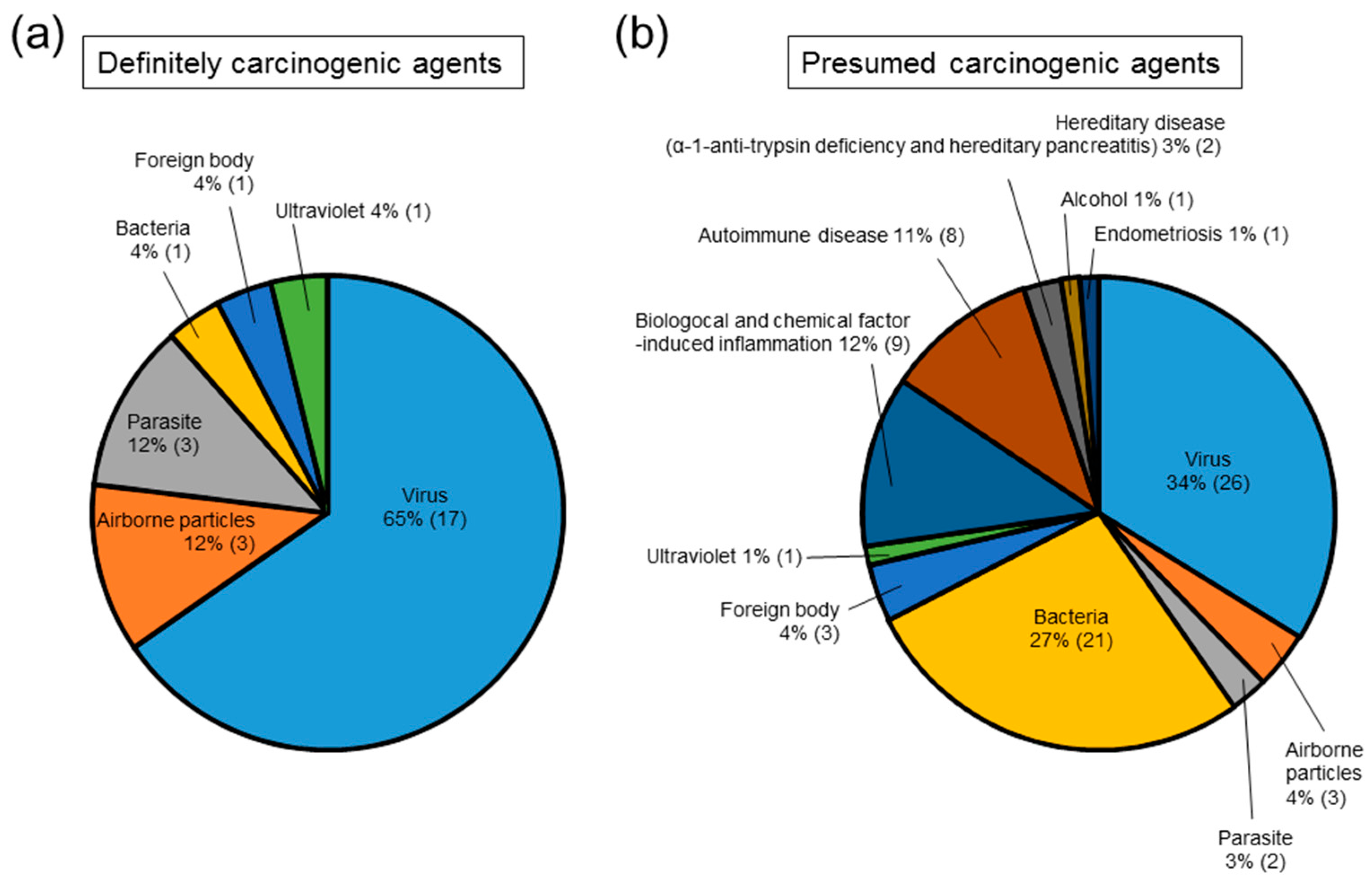
2. Causes of Inflammation-Related Carcinogenesis
Inhaled Foreign Body-Induced Carcinogenesis
3. Animal Models for Inflammation-Related Cancer Chemoprevention Studies
3.1. Esophageal Cancer
3.2. Gastric Cancer
3.3. Colon Cancer
3.4. Hepatocellular Carcinoma
3.5. Cholangiocarcinoma
3.6. Biliary Tract Cancer
3.7. Pancreatic Ductal Adenocarcinoma
3.8. Skin Cancer
3.9. Experimental Models of Foreign Body-Induced Carcinogenesis
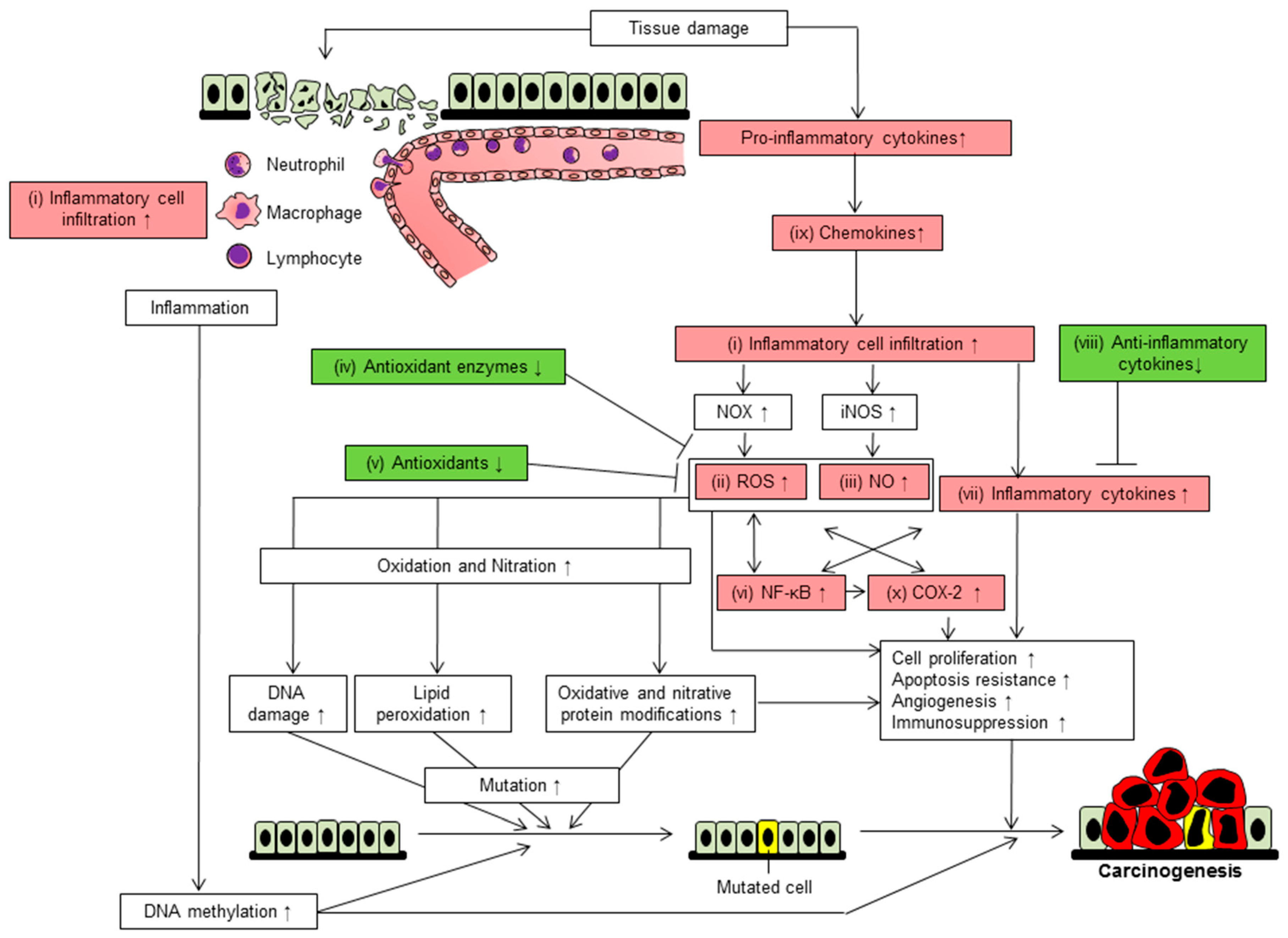
4. Ten Mechanisms Involved in Inflammation-Related Carcinogenesis-Based Chemoprevention
4.1. Inflammatory Cell Infiltration
4.2. Reactive Oxygen Species
4.3. Nitric Oxide
4.4. Reduction of Antioxidant Enzymes
4.5. Reduction of Antioxidant
4.6. Activation of NF-κB
4.7. Upregulation of Pro-Inflammatory Cytokines
4.8. Downregulation of Anti-Inflammatory Cytokines
4.9. Elevation of Chemokines
4.10. Induction of Cyclooxygenase-2
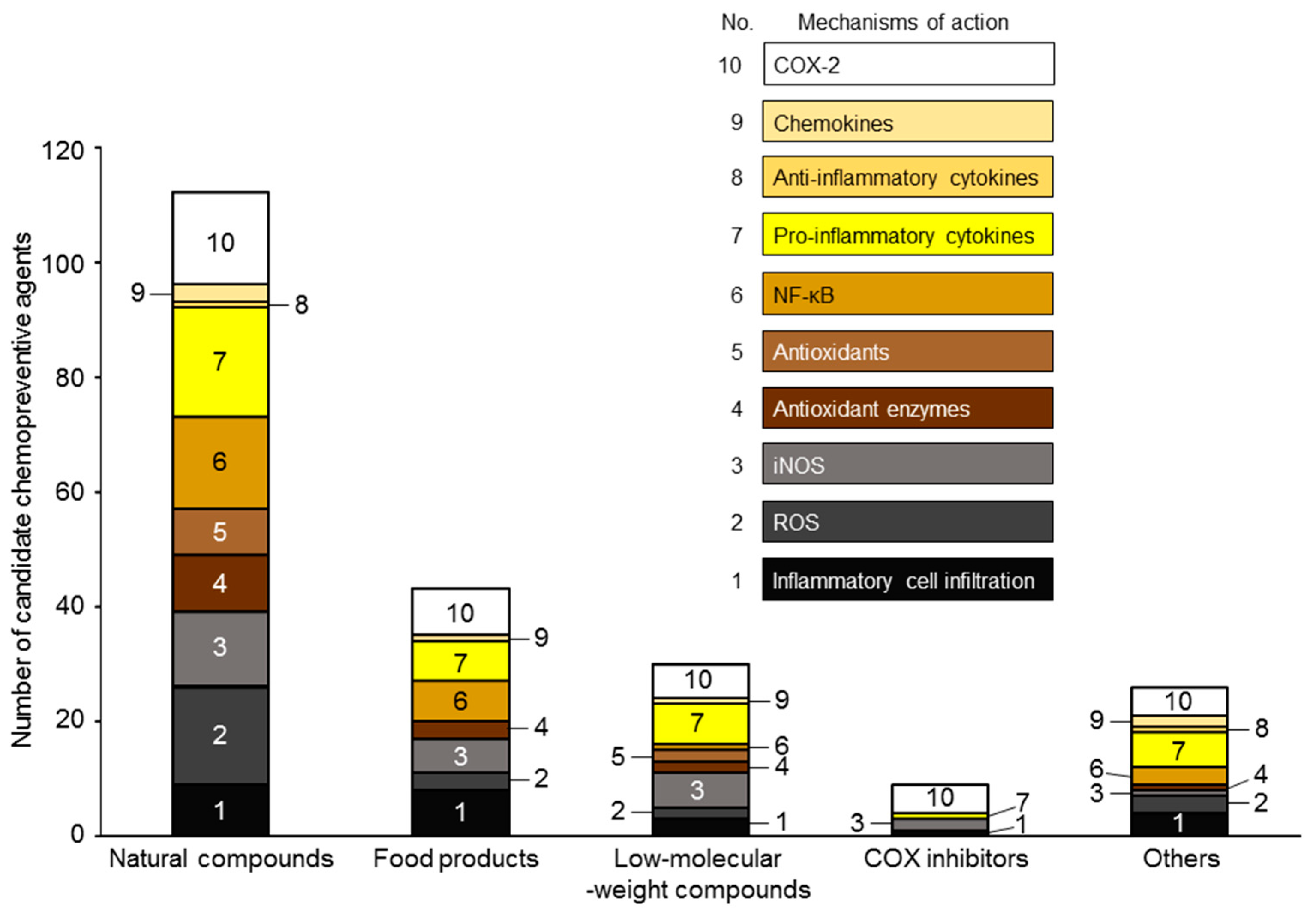
5. Candidate Chemopreventive Agents against Inflammation-Related Carcinogenesis
6. Future Prospects
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Asian dust |
| AOM | Azoxymethane |
| Apc | Adenomatous polyposis coli |
| ATL | Adult T-cell leukemia |
| CCL | C-C motif chemokine ligand |
| CCl4 | Carbon tetra chloride |
| COPD | Chronic obstructive pulmonary disease |
| COX | Cyclooxygenase |
| CXCL | Chemokine (C-X-C motif) ligand |
| DBM | Dibenzoylmethane |
| DEN | Diethylnitrosamine |
| DLBC | Diffuse large B-cell |
| DMBA | 7,12-Dimethylbenz[a]-anthracene |
| DMH | Dimethylhydrazine |
| DSS | Dextran sulfate sodium |
| EAPP | Ethanol extracts from the aerial parts of A. princeps Pampanini cv. Sajabal |
| EBV | Epstein-Barr virus |
| EGF | Epidermal growth factor |
| EVOO | Extra virgin olive oil |
| FAP | Familial adenomatous polyposis |
| FBE | Fruiting body extract |
| FBRA | Fermented brown rice and rice bran with Aspergillus oryzae |
| GOFA/β-CD | 3-(4′-Geranyloxy-3′-methoxyphenyl)-2-trans propenoic acid/β-cyclodextrin |
| GOFA-L-NAME | 4′-Geranyloxyferulic acid-N(omega)-nitro-l-arginine methyl ester |
| H. felis | Helicobacter felis |
| H. pylori | Helicobacter pylori |
| HBV | Hepatitis B virus |
| HCC | Hepatocellular carcinoma |
| HCV | Hepatitis C virus |
| HDV | Hepatitis D virus |
| HERV-K | Human endogenous retrovirus type K |
| HIV | Human immunodeficiency virus |
| HPV | Human papillomavirus |
| HTLV-1 | Human T-cell lymphotropic virus type 1 |
| IARC | International Agency for Research on Cancer |
| IBD | Inflammatory bowel disease |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| JCV | JC virus |
| KSHV | Kaposi sarcoma herpes virus |
| MALT | Mucosa-associated lymphoid tissue |
| MCV | Molluscum contagiosum virus |
| MDA | Malondialdehydes |
| ME | Mycelia extract |
| MEFA | Methanol extracts of the fruit of A. communis |
| MELA | Methanol extract of the leaf of A. communis |
| MiR | MicroRNA |
| MNNG | N-Methyl-N′-nitro-N-nitrosoguanidine |
| Mn-SOD | Manganese superoxide dismutase |
| NDMA | N-Nitrosodimethylamine |
| NO | Nitric oxide |
| NSAID | Nonsteroidal anti-inflammatory drug |
| O. viverrini | Opisthorchis viverrini |
| PAG | Processed Aloe vera gel |
| PBM | Pancreaticobiliary maljunction |
| PDAC | Pancreatic ductal adenocarcinomas |
| PEITC | Phenethyl isothiocyanate |
| PGE | Prostaglandin E |
| PhIP | 2-Amino-1-methyl-6-phenylimidazo[4,5-b] pyridine |
| PSK | Polysaccharide K |
| ROS | Reactive oxygen species |
| SCC | Squamous cell carcinoma |
| TNF | Tumor necrosis factor |
| TPA | 12-O-Tetradecanoylphorbol-13-acetate |
| UDCA | Ursodeoxycholic acid |
| UV | Ultraviolet |
| γ-TmT | γ-Tocopherol-rich mixture of tocopherols |
| 4-HNE | 4-Hydroxynonenal |
| 5-OH-HxMF | 5-Hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone |
| 13-HOA | (±)-13-Hydroxy-10-oxo-trans-11-octadecenoic acid |
References
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Parkin, D.M. The global health burden of infection-associated cancers in the year 2002. Int. J. Cancer 2006, 118, 3030–3044. [Google Scholar] [CrossRef] [PubMed]
- Belpomme, D.; Irigaray, P.; Hardell, L.; Clapp, R.; Montagnier, L.; Epstein, S.; Sasco, A.J. The multitude and diversity of environmental carcinogens. Environ. Res. 2007, 105, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Chan, R.L.; Luo, X.M.; Wu, W.K.; Shin, V.Y.; Cho, C.H. Animal models of gastrointestinal inflammation and cancer. Life Sci. 2014, 108, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Omata, M. Inflammation and cancer: Role of nuclear factor-κB activation. Cancer Sci. 2008, 99, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Cogliano, V.J.; Baan, R.; Straif, K.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; et al. Preventable exposures associated with human cancers. J. Natl. Cancer Inst. 2011, 103, 1827–1839. [Google Scholar] [CrossRef] [PubMed]
- Elinav, E.; Nowarski, R.; Thaiss, C.A.; Hu, B.; Jin, C.; Flavell, R.A. Inflammation-induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat. Rev. Cancer 2013, 13, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Miyoshi, N.; Tomono, S. Infection, Inflammation, and Cancer: Overview. In Cancer and Inflammation Mechanisms; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 1–7. [Google Scholar]
- Wen, B.W.; Tsai, C.S.; Lin, C.L.; Chang, Y.J.; Lee, C.F.; Hsu, C.H.; Kao, C.H. Cancer risk among gingivitis and periodontitis patients: A nationwide cohort study. QJM 2014, 107, 283–290. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Biological agents. Volume 100 B. A review of human carcinogens. IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100, 1–441. [Google Scholar]
- Siirala, U. Tongue cancer. Acta Otolaryngol. 1973, 75, 309. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, Y.; Choi, J.W.; Kim, Y.S. The association between papillary thyroid carcinoma and histologically proven Hashimoto’s thyroiditis: A meta-analysis. Eur. J. Endocrinol. 2013, 168, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Muto, M.; Hitomi, Y.; Ohtsu, A.; Shimada, H.; Kashiwase, Y.; Sasaki, H.; Yoshida, S.; Esumi, H. Acetaldehyde production by non-pathogenic Neisseria in human oral microflora: Implications for carcinogenesis in upper aerodigestive tract. Int. J. Cancer 2000, 88, 342–350. [Google Scholar] [CrossRef]
- Loomis, D.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Baan, R.; Mattock, H.; Straif, K.; et al. The carcinogenicity of outdoor air pollution. Lancet Oncol. 2013, 14, 1262–1263. [Google Scholar] [CrossRef]
- Santillan, A.A.; Camargo, C.A., Jr.; Colditz, G.A. A meta-analysis of asthma and risk of lung cancer (United States). Cancer Causes Control 2003, 14, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A.M. Mechanistic links between COPD and lung cancer. Nat. Rev. Cancer 2013, 13, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, H.; Tanaka, S.; Saiki, Y.; Hara, M.; Nakata, K.; Tanimura, S.; Banba, J. Lung cancer associated with usual interstitial pneumonia. Pathol. Int. 1995, 45, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Odaka, M.; Hosoda, Y.; Iwai, K.; Tachibana, T. Excess death of lung cancer among sarcoidosis patients. Sarcoidosis 1991, 8, 51–55. [Google Scholar] [PubMed]
- Wu, C.Y.; Hu, H.Y.; Pu, C.Y.; Huang, N.; Shen, H.C.; Li, C.P.; Chou, Y.J. Pulmonary tuberculosis increases the risk of lung cancer: A population-based cohort study. Cancer 2011, 117, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Suo, L.J.; Qian, Q.; Shen, X.K.; Qiu, L.X.; Yu, L.K.; Song, Y. Chlamydia pneumoniae infection and lung cancer risk: A meta-analysis. Eur. J. Cancer 2011, 47, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Ragin, C.; Obikoya-Malomo, M.; Kim, S.; Chen, Z.; Flores-Obando, R.; Gibbs, D.; Koriyama, C.; Aguayo, F.; Koshiol, J.; Caporaso, N.E.; et al. HPV-associated lung cancers: An international pooled analysis. Carcinogenesis 2014, 35, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Kirk, G.D.; Merlo, C.; O’Driscoll, P.; Mehta, S.H.; Galai, N.; Vlahov, D.; Samet, J.; Engels, E.A. HIV infection is associated with an increased risk for lung cancer, independent of smoking. Clin. Infect. Dis. 2007, 45, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Steenland, K.; Stayner, L. Silica, asbestos, man-made mineral fibers, and cancer. Cancer Causes Control 1997, 8, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Salmons, B.; Lawson, J.S.; Gunzburg, W.H. Recent developments linking retroviruses to human breast cancer: Infectious agent, enemy within or both? J. Gen. Virol. 2014, 95, 2589–2593. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.; Wedemeyer, H. Hepatitis delta: Immunopathogenesis and clinical challenges. Dig. Dis. 2010, 28, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, N.M. Recent insights on risk factors of hepatocellular carcinoma. World J. Hepatol. 2009, 1, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Talwalkar, J.A. Primary sclerosing cholangitis: Diagnosis, prognosis, and management. Clin. Gastroenterol. Hepatol. 2013, 11, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Kamisawa, T.; Kuruma, S.; Chiba, K.; Tabata, T.; Koizumi, S.; Kikuyama, M. Biliary carcinogenesis in pancreaticobiliary maljunction. J. Gastroenterol. 2017, 52, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Scanu, T.; Spaapen, R.M.; Bakker, J.M.; Pratap, C.B.; Wu, L.E.; Hofland, I.; Broeks, A.; Shukla, V.K.; Kumar, M.; Janssen, H.; et al. Salmonella manipulation of host signaling pathways provokes cellular transformation associated with gallbladder carcinoma. Cell Host Microbe 2015, 17, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Weiss, F.U. Pancreatic cancer risk in hereditary pancreatitis. Front. Physiol. 2014, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; McLaughlin, J.K.; Gridley, G.; Bjelke, E.; Schuman, L.M.; Silverman, D.T.; Wacholder, S.; Co-Chien, H.T.; Blot, W.J.; Fraumeni, J.F., Jr. A cohort study of smoking, alcohol consumption, and dietary factors for pancreatic cancer (United States). Cancer Causes Control 1993, 4, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Hartnett, L.; Egan, L.J. Inflammation, DNA methylation and colitis-associated cancer. Carcinogenesis 2012, 33, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.; Hogan, A.M.; Winter, D.C. Microbial and viral pathogens in colorectal cancer. Lancet Oncol. 2011, 12, 504–512. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial driver-passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Boleij, A.; van Gelder, M.M.; Swinkels, D.W.; Tjalsma, H. Clinical importance of streptococcus gallolyticus infection among colorectal cancer patients: Systematic review and meta-analysis. Clin. Infect. Dis. 2011, 53, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.E.; Bassett, M.T.; Sigola, L.B.; Taylor, P. Relationship between bladder cancer incidence, Schistosoma haematobium infection, and geographical region in Zimbabwe. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 551–553. [Google Scholar] [CrossRef]
- West, D.A.; Cummings, J.M.; Longo, W.E.; Virgo, K.S.; Johnson, F.E.; Parra, R.O. Role of chronic catheterization in the development of bladder cancer in patients with spinal cord injury. Urology 1999, 53, 292–297. [Google Scholar] [CrossRef]
- Ky, A.; Sohn, N.; Weinstein, M.A.; Korelitz, B.I. Carcinoma arising in anorectal fistulas of Crohn’s disease. Dis. Colon Rectum. 1998, 41, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Akre, O.; Lipworth, L.; Tretli, S.; Linde, A.; Engstrand, L.; Adami, H.O.; Melbye, M.; Andersen, A.; Ekbom, A. Epstein-Barr virus and cytomegalovirus in relation to testicular-cancer risk: A nested case-control study. Int. J. Cancer 1999, 82, 1–5. [Google Scholar] [CrossRef]
- De Marzo, A.M.; Platz, E.A.; Sutcliffe, S.; Xu, J.; Gronberg, H.; Drake, C.G.; Nakai, Y.; Isaacs, W.B.; Nelson, W.G. Inflammation in prostate carcinogenesis. Nat. Rev. Cancer 2007, 7, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Dennis, L.K.; Dawson, D.V. Meta-analysis of measures of sexual activity and prostate cancer. Epidemiology 2002, 13, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.R.; Judson, G.; Alderete, J.F.; Mundodi, V.; Kucknoor, A.S.; Giovannucci, E.L.; Platz, E.A.; Sutcliffe, S.; Fall, K.; Kurth, T.; et al. Prospective study of Trichomonas vaginalis infection and prostate cancer incidence and mortality: Physicians’ health study. J. Natl. Cancer Inst. 2009, 101, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Kralickova, M.; Vetvicka, V. Endometriosis and ovarian cancer. World J. Clin. Oncol. 2014, 5, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, M.C.; Visser, P.J.; Overbeek, L.I.; van Beurden, M.; Berkhof, J. Lichen sclerosus: Incidence and risk of vulvar squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2016, 25, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Hejna, W.F. Squamous-cell carcinoma developing in the chronic draining sinuses of osteomyelitis. Cancer 1965, 18, 128–132. [Google Scholar] [CrossRef]
- Arron, S.T.; Jennings, L.; Nindl, I.; Rosl, F.; Bouwes Bavinck, J.N.; Seckin, D.; Trakatelli, M.; Murphy, G.M. Viral Working Group of the International Transplant Skin Cancer Collaborative (ITSCC); Care in Organ Transplant Patients, Europe (SCOPE). Viral oncogenesis and its role in nonmelanoma skin cancer. Br. J. Dermatol. 2011, 164, 1201–1213. [Google Scholar] [CrossRef] [PubMed]
- Reiss, K.; Khalili, K. Viruses and cancer: Lessons from the human polyomavirus, JCV. Oncogene 2003, 22, 6517–6523. [Google Scholar] [CrossRef] [PubMed]
- Dehio, C. Bartonella-host-cell interactions and vascular tumour formation. Nat. Rev. Microbiol. 2005, 3, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Grulich, A.E.; van Leeuwen, M.T.; Falster, M.O.; Vajdic, C.M. Incidence of cancers in people with HIV/AIDS compared with immunosuppressed transplant recipients: A meta-analysis. Lancet 2007, 370, 59–67. [Google Scholar] [CrossRef]
- Jeong, S.H. HBV infection as a risk factor for non-Hodgkin lymphoma. Lancet Oncol. 2010, 11, 806. [Google Scholar] [CrossRef]
- Colebunders, R.; de Vuyst, H.; Verstraeten, T.; Schroyens, W.; van Marck, E. A non-Hodgkin’s lymphoma in a patient with HIV-2 infection. Genitourin. Med. 1995, 71, 129. [Google Scholar] [CrossRef] [PubMed]
- Green, P.H.; Fleischauer, A.T.; Bhagat, G.; Goyal, R.; Jabri, B.; Neugut, A.I. Risk of malignancy in patients with celiac disease. Am. J. Med. 2003, 115, 191–195. [Google Scholar] [CrossRef]
- Galun, E.; Ilan, Y.; Livni, N.; Ketzinel, M.; Nahor, O.; Pizov, G.; Nagler, A.; Eid, A.; Rivkind, A.; Laster, M.; et al. Hepatitis B virus infection associated with hematopoietic tumors. Am. J. Pathol. 1994, 145, 1001–1007. [Google Scholar] [PubMed]
- Coffin, J.M. The discovery of HTLV-1, the first pathogenic human retrovirus. Proc. Natl. Acad. Sci. USA 2015, 112, 15525–15529. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, A.J.; Guidoboni, M.; Ponzoni, M.; de Conciliis, C.; Dell’Oro, S.; Fleischhauer, K.; Caggiari, L.; Lettini, A.A.; Dal Cin, E.; Ieri, R.; et al. Evidence for an association between Chlamydia psittaci and ocular adnexal lymphomas. J. Natl. Cancer Inst. 2004, 96, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Aozasa, K. Hashimoto’s thyroiditis as a risk factor of thyroid lymphoma. Acta Pathol. Jpn. 1990, 40, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Molinie, V.; Pouchot, J.; Navratil, E.; Aubert, F.; Vinceneux, P.; Barge, J. Primary Epstein-Barr virus-related non-Hodgkin’s lymphoma of the pleural cavity following long-standing tuberculous empyema. Arch. Pathol. Lab. Med. 1996, 120, 288–291. [Google Scholar] [PubMed]
- Aozasa, K.; Takakuwa, T.; Nakatsuka, S. Pyothorax-associated lymphoma: A lymphoma developing in chronic inflammation. Adv. Anat. Pathol. 2005, 12, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Lecuit, M.; Abachin, E.; Martin, A.; Poyart, C.; Pochart, P.; Suarez, F.; Bengoufa, D.; Feuillard, J.; Lavergne, A.; Gordon, J.I.; et al. Immunoproliferative small intestinal disease associated with Campylobacter jejuni. N. Engl. J. Med. 2004, 350, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Goodlad, J.R.; Davidson, M.M.; Hollowood, K.; Ling, C.; MacKenzie, C.; Christie, I.; Batstone, P.J.; Ho-Yen, D.O. Primary cutaneous B-cell lymphoma and Borrelia burgdorferi infection in patients from the Highlands of Scotland. Am. J. Surg. Pathol. 2000, 24, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, J.; Matsuoka, M. Molecular mechanisms of HTLV-1 infection and pathogenesis. Int. J. Hematol. 2011, 94, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Piccaluga, P.P.; Gazzola, A.; Agostinelli, C.; Bacci, F.; Sabattini, E.; Pileri, S.A. Pathobiology of Epstein-Barr virus-driven peripheral T-cell lymphomas. Semin. Diagn. Pathol. 2011, 28, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Rochford, R.; Moormann, A.M. Burkitt’s Lymphoma. Curr. Top. Microbiol. Immunol. 2015, 390, 267–285. [Google Scholar] [PubMed]
- Grywalska, E.; Rolinski, J. Epstein-Barr virus-associated lymphomas. Semin. Oncol. 2015, 42, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.W.; Damania, B. Kaposi sarcoma-associated herpesvirus (KSHV): Molecular biology and oncogenesis. Cancer Lett. 2010, 289, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Samaras, V.; Rafailidis, P.I.; Mourtzoukou, E.G.; Peppas, G.; Falagas, M.E. Chronic bacterial and parasitic infections and cancer: A review. J. Infect. Dev. Ctries 2010, 4, 267–281. [Google Scholar] [PubMed]
- Mesri, E.A.; Feitelson, M.A.; Munger, K. Human viral oncogenesis: A cancer hallmarks analysis. Cell Host Microbe 2014, 15, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Meng, W.; Wang, B.; Qiao, L. Helicobacter pylori-induced gastric inflammation and gastric cancer. Cancer Lett. 2014, 345, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Okada, F. Beyond foreign-body-induced carcinogenesis: Impact of reactive oxygen species derived from inflammatory cells in tumorigenic conversion and tumor progression. Int. J. Cancer 2007, 121, 2364–2372. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Toyokuni, S. Biopersistent fiber-induced inflammation and carcinogenesis: Lessons learned from asbestos toward safety of fibrous nanomaterials. Arch. Biochem. Biophys. 2010, 502, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.H.; Toyokuni, S. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Gudima, A.; Riabov, V.; Dollinger, C.; Lavalle, P.; Vrana, N.E. Macrophage responses to implants: Prospects for personalized medicine. J. Leukoc. Biol. 2015, 98, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Toyokuni, S. Differences and similarities between carbon nanotubes and asbestos fibers during mesothelial carcinogenesis: Shedding light on fiber entry mechanism. Cancer Sci. 2012, 103, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.F.; Xu, Y.H.; Shi, M.H.; Lian, Y.X. The impact of PM2.5 on the human respiratory system. J. Thorac. Dis. 2016, 8, E69–E74. [Google Scholar] [PubMed]
- Watanabe, M.; Noma, H.; Kurai, J.; Sano, H.; Saito, R.; Abe, S.; Kimura, Y.; Aiba, S.; Oshimura, M.; Yamasaki, A.; et al. Decreased pulmonary function in school children in Western Japan after exposures to Asian desert dusts and its association with interleukin-8. BioMed Res. Int. 2015, 2015, 583293. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Noma, H.; Kurai, J.; Sano, H.; Kitano, H.; Saito, R.; Kimura, Y.; Aiba, S.; Oshimura, M.; Shimizu, E. Variation in the effect of particulate matter on pulmonary function in schoolchildren in western japan and its relation with interleukin-8. Int. J. Environ. Res. Public Health 2015, 12, 14229–14243. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Noma, H.; Kurai, J.; Sano, H.; Ueda, Y.; Mikami, M.; Yamamoto, H.; Tokuyasu, H.; Kato, K.; Konishi, T.; et al. Differences in the effects of Asian dust on pulmonary function between adult patients with asthma and those with asthma-chronic obstructive pulmonary disease overlap syndrome. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.C.; Krewski, D.; Pope, C.A., III; Chen, Y.; Gapstur, S.M.; Thun, M.J. Long-term ambient fine particulate matter air pollution and lung cancer in a large cohort of never-smokers. Am. J. Respir. Crit. Care Med. 2011, 184, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- King, P.T. Inflammation in chronic obstructive pulmonary disease and its role in cardiovascular disease and lung cancer. Clin. Transl. Med. 2015, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Young, R.P.; Hopkins, R.J.; Christmas, T.; Black, P.N.; Metcalf, P.; Gamble, G.D. COPD prevalence is increased in lung cancer, independent of age, sex and smoking history. Eur. Respir. J. 2009, 34, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.K.; Sporn, M.B. Recent advances in chemoprevention of cancer. Science 1997, 278, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Miwa, K.; Sahara, H.; Segawa, M.; Kinami, S.; Sato, T.; Miyazaki, I.; Hattori, T. Reflux of duodenal or gastro-duodenal contents induces esophageal carcinoma in rats. Int. J. Cancer 1996, 67, 269–274. [Google Scholar] [CrossRef]
- Tatematsu, M.; Yamamoto, M.; Shimizu, N.; Yoshikawa, A.; Fukami, H.; Kaminishi, M.; Oohara, T.; Sugiyama, A.; Ikeno, T. Induction of glandular stomach cancers in Helicobacter pylori-sensitive Mongolian gerbils treated with N-methyl-N-nitrosourea and N-methyl-N′-nitro-N-nitrosoguanidine in drinking water. Jpn. J. Cancer Res. 1998, 89, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Clapper, M.L.; Cooper, H.S.; Chang, W.C. Dextran sulfate sodium-induced colitis-associated neoplasia: A promising model for the development of chemopreventive interventions. Acta Pharmacol. Sin. 2007, 28, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Animal models of carcinogenesis in inflamed colorectum: Potential use in chemoprevention study. Curr. Drug Targets 2012, 13, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Dunsford, H.A.; Sell, S.; Chisari, F.V. Hepatocarcinogenesis due to chronic liver cell injury in hepatitis B virus transgenic mice. Cancer Res. 1990, 50, 3400–3407. [Google Scholar] [PubMed]
- Schiffer, E.; Housset, C.; Cacheux, W.; Wendum, D.; Desbois-Mouthon, C.; Rey, C.; Clergue, F.; Poupon, R.; Barbu, V.; Rosmorduc, O. Gefitinib, an EGFR inhibitor, prevents hepatocellular carcinoma development in the rat liver with cirrhosis. Hepatology 2005, 41, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Uehara, T.; Ainslie, G.R.; Kutanzi, K.; Pogribny, I.P.; Muskhelishvili, L.; Izawa, T.; Yamate, J.; Kosyk, O.; Shymonyak, S.; Bradford, B.U.; et al. Molecular mechanisms of fibrosis-associated promotion of liver carcinogenesis. Toxicol. Sci. 2013, 132, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Thamavit, W.; Bhamarapravati, N.; Sahaphong, S.; Vajrasthira, S.; Angsubhakorn, S. Effects of dimethylnitrosamine on induction of cholangiocarcinoma in Opisthorchis viverrini-infected Syrian golden hamsters. Cancer Res. 1978, 38, 4634–4639. [Google Scholar] [PubMed]
- Tajima, Y.; Eto, T.; Tsunoda, T.; Tomioka, T.; Inoue, K.; Fukahori, T.; Kanematsu, T. Induction of extrahepatic biliary carcinoma by N-nitrosobis(2-oxopropyl)amine in hamsters given cholecystoduodenostomy with dissection of the common duct. Jpn. J. Cancer Res. 1994, 85, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.; Schuhmacher, A.J.; Canamero, M.; Grippo, P.J.; Verdaguer, L.; Perez-Gallego, L.; Dubus, P.; Sandgren, E.P.; Barbacid, M. Chronic pancreatitis is essential for induction of pancreatic ductal adenocarcinoma by K-Ras oncogenes in adult mice. Cancer Cell 2007, 11, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Munzel, P.A.; Braeuning, A. Non-melanoma skin cancer in mouse and man. Arch. Toxicol. 2013, 87, 783–978. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, H.; Lohani, K.R.; Lee, T.H.; Agrawal, D.K.; Mittal, S.K. Animal models of Barrett’s esophagus and esophageal adenocarcinoma—Past, present, and future. Clin. Transl. Sci. 2015, 8, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Genta, R.M.; Spechler, S.J.; Souza, R.F.; Wang, D.H. Development and characterization of a surgical mouse model of reflux esophagitis and Barrett’s esophagus. J. Gastrointest. Surg. 2014, 18, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Buskens, C.J.; Hulscher, J.B.; van Gulik, T.M.; Ten Kate, F.J.; van Lanschot, J.J. Histopathologic evaluation of an animal model for Barrett’s esophagus and adenocarcinoma of the distal esophagus. J. Surg. Res. 2006, 135, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.G.; Wang, T.C.; Rogers, A.B.; Poutahidis, T.; Ge, Z.; Taylor, N.; Dangler, C.A.; Israel, D.A.; Krishna, U.; Gaus, K.; et al. Host and microbial constituents influence Helicobacter pylori-induced cancer in a murine model of hypergastrinemia. Gastroenterology 2003, 124, 1879–1890. [Google Scholar] [CrossRef]
- Rogers, A.B.; Fox, J.G. Inflammation and Cancer. I. Rodent models of infectious gastrointestinal and liver cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G361–G366. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Toyoda, T.; Mizoshita, T.; Tatematsu, M. Helicobacter pylori infection and gastric carcinogenesis in rodent models. Semin. Immunopathol. 2013, 35, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Redline, R.; Nedrud, J.; Czinn, S. Role of the host in pathogenesis of Helicobacter-associated gastritis: H. felis infection of inbred and congenic mouse strains. Infect. Immun. 1996, 64, 238–245. [Google Scholar] [PubMed]
- Nakamura, Y.; Sakagami, T.; Yamamoto, N.; Yokota, Y.; Koizuka, H.; Hori, K.; Fukuda, Y.; Tanida, N.; Kobayashi, T.; Shimoyama, T. Helicobacter pylori does not promote N-methyl-N-nitrosourea-induced gastric carcinogenesis in SPF C57BL/6 mice. Jpn. J. Cancer Res. 2002, 93, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Perse, M.; Cerar, A. Dextran sodium sulphate colitis mouse model: Traps and tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef] [PubMed]
- Melgar, S.; Karlsson, A.; Michaelsson, E. Acute colitis induced by dextran sulfate sodium progresses to chronicity in C57BL/6 but not in BALB/c mice: Correlation between symptoms and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Hursting, S.D.; Slaga, T.J.; Fischer, S.M.; DiGiovanni, J.; Phang, J.M. Mechanism-based cancer prevention approaches: Targets, examples, and the use of transgenic mice. J. Natl. Cancer Inst. 1999, 91, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, H.L.; Kettunen, A.S.; Rautonen, N.E. Intestinal immune responses in wild-type and Apcmin/+ mouse, a model for colon cancer. Cancer Res. 2003, 63, 5136–5142. [Google Scholar] [PubMed]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Hata, K.; Sugie, S.; Niho, N.; Sakano, K.; Takahashi, M.; Wakabayashi, K. Dextran sodium sulfate strongly promotes colorectal carcinogenesis in ApcMin/+ mice: Inflammatory stimuli by dextran sodium sulfate results in development of multiple colonic neoplasms. Int. J. Cancer 2006, 118, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Bakiri, L.; Wagner, E.F. Mouse models for liver cancer. Mol. Oncol. 2013, 7, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Babinet, C.; Farza, H.; Morello, D.; Hadchouel, M.; Pourcel, C. Specific expression of hepatitis B surface antigen (HBsAg) in transgenic mice. Science 1985, 230, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- McGivern, D.R.; Lemon, S.M. Virus-specific mechanisms of carcinogenesis in hepatitis C virus associated liver cancer. Oncogene 2011, 30, 1969–1983. [Google Scholar] [CrossRef] [PubMed]
- Rajewsky, M.F.; Dauber, W.; Frankenberg, H. Liver carcinogenesis by diethylnitrosamine in the rat. Science 1966, 152, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Thamavit, W.; Kongkanuntn, R.; Tiwawech, D.; Moore, M.A. Level of Opisthorchis infestation and carcinogen dose-dependence of cholangiocarcinoma induction in Syrian golden hamsters. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1987, 54, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Ohshima, H.; Srivatanakul, P.; Vatanasapt, V. Cholangiocarcinoma: Epidemiology, mechanisms of carcinogenesis and prevention. Cancer Epidemiol. Biomark. Prev. 1993, 2, 537–544. [Google Scholar]
- Ames, B.N.; Gold, L.S. Chemical carcinogenesis: Too many rodent carcinogens. Proc. Natl. Acad. Sci. USA 1990, 87, 7772–7776. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Ellwein, L.B. Cell proliferation in carcinogenesis. Science 1990, 249, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, S.A.; Gordon, L.I. Inflammation and cancer: Role of phagocyte-generated oxidants in carcinogenesis. Blood 1990, 76, 655–663. [Google Scholar] [PubMed]
- Li, X.; Wang, S.; Zhu, R.; Li, H.; Han, Q.; Zhao, R.C. Lung tumor exosomes induce a pro-inflammatory phenotype in mesenchymal stem cells via NF-κB-TLR signaling pathway. J. Hematol. Oncol. 2016, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Chaiyadet, S.; Sotillo, J.; Smout, M.; Cantacessi, C.; Jones, M.K.; Johnson, M.S.; Turnbull, L.; Whitchurch, C.B.; Potriquet, J.; Laohaviroj, M.; et al. Carcinogenic liver fluke secretes extracellular vesicles that promote cholangiocytes to adopt a tumorigenic phenotype. J. Infect. Dis. 2015, 212, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, A.; Itoi, T. Carcinogenesis and chemoprevention of biliary tract cancer in pancreaticobiliary maljunction. World J. Gastrointest. Oncol. 2010, 2, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Tajima, Y.; Kitajima, T.; Tomioka, T.; Eto, T.; Inoue, K.; Fukahori, T.; Sasaki, M.; Tsunoda, T. Hamster Models of Biliary Carcinoma. In Hepatobiliary and Pancreatic Carcinogenesis in the Hamster; Springer: Heidelberg, Germany; Dordrecht, The Netherlands, 2009; pp. 29–68. [Google Scholar]
- Tsuchida, A.; Itoi, T.; Kasuya, K.; Endo, M.; Katsumata, K.; Aoki, T.; Suzuki, M.; Aoki, T. Inhibitory effect of meloxicam, a cyclooxygenase-2 inhibitor, on N-nitrosobis (2-oxopropyl) amine induced biliary carcinogenesis in Syrian hamsters. Carcinogenesis 2005, 26, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.L. ras Oncogenes in human cancer: A review. Cancer Res. 1989, 49, 4682–4689. [Google Scholar] [PubMed]
- Almoguera, C.; Shibata, D.; Forrester, K.; Martin, J.; Arnheim, N.; Perucho, M. Most human carcinomas of the exocrine pancreas contain mutant c-K-ras genes. Cell 1988, 53, 549–554. [Google Scholar] [CrossRef]
- Boone, C.W.; Steele, V.E.; Kelloff, G.J. Screening for chemopreventive (anticarcinogenic) compounds in rodents. Mutat. Res. 1992, 267, 251–255. [Google Scholar] [CrossRef]
- Hennings, H.; Glick, A.B.; Lowry, D.T.; Krsmanovic, L.S.; Sly, L.M.; Yuspa, S.H. FVB/N mice: An inbred strain sensitive to the chemical induction of squamous cell carcinomas in the skin. Carcinogenesis 1993, 14, 2353–2358. [Google Scholar] [CrossRef] [PubMed]
- Nassar, D.; Latil, M.; Boeckx, B.; Lambrechts, D.; Blanpain, C. Genomic landscape of carcinogen-induced and genetically induced mouse skin squamous cell carcinoma. Nat. Med. 2015, 21, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, H.; Tatemichi, M.; Sawa, T.; Gilibert, I.; Ma, N.; Hiraku, Y.; Donehower, L.A.; Ohgaki, H.; Kawanishi, S.; Ohshima, H. Oxidative and nitrative stress caused by subcutaneous implantation of a foreign body accelerates sarcoma development in Trp53+/− mice. Carcinogenesis 2007, 28, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Jennings, T.A.; Peterson, L.; Axiotis, C.A.; Friedlaender, G.E.; Cooke, R.A.; Rosai, J. Angiosarcoma associated with foreign body material. A report of three cases. Cancer 1988, 62, 2436–2444. [Google Scholar] [CrossRef]
- Okada, F.; Kawaguchi, T.; Habelhah, H.; Kobayashi, T.; Tazawa, H.; Takeichi, N.; Kitagawa, T.; Hosokawa, M. Conversion of human colonic adenoma cells to adenocarcinoma cells through inflammation in nude mice. Lab. Investig. 2000, 80, 1617–1628. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, H.; Kawaguchi, T.; Kobayashi, T.; Kuramitsu, Y.; Wada, S.; Satomi, Y.; Nishino, H.; Kobayashi, M.; Kanda, Y.; Osaki, M.; et al. Chronic inflammation-derived nitric oxide causes conversion of human colonic adenoma cells into adenocarcinoma cells. Exp. Cell Res. 2013, 319, 2835–2844. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y.; Kawaguchi, T.; Kuramitsu, Y.; Kitagawa, T.; Kobayashi, T.; Takahashi, N.; Tazawa, H.; Habelhah, H.; Hamada, J.; Kobayashi, M.; et al. Fascin regulates chronic inflammation-related human colon carcinogenesis by inhibiting cell anoikis. Proteomics 2014, 14, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Hosokawa, M.; Hamada, J.I.; Hasegawa, J.; Kato, M.; Mizutani, M.; Ren, J.; Takeichi, N.; Kobayashi, H. Malignant progression of a mouse fibrosarcoma by host cells reactive to a foreign body (gelatin sponge). Br. J. Cancer 1992, 66, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, H.; Okada, F.; Kobayashi, T.; Tada, M.; Mori, Y.; Une, Y.; Sendo, F.; Kobayashi, M.; Hosokawa, M. Infiltration of neutrophils is required for acquisition of metastatic phenotype of benign murine fibrosarcoma cells: Implication of inflammation-associated carcinogenesis and tumor progression. Am. J. Pathol. 2003, 163, 2221–2232. [Google Scholar] [CrossRef]
- Okada, F. Inflammation-related carcinogenesis: Current findings in epidemiological trends, causes and mechanisms. Yonago Acta Med. 2014, 57, 65–72. [Google Scholar] [PubMed]
- Hamilton, J.A.; Achuthan, A. Colony stimulating factors and myeloid cell biology in health and disease. Trends Immunol. 2013, 34, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y. Tumor-associated macrophages: From basic research to clinical application. J. Hematol. Oncol. 2017, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.; Rachidi, S.; Hong, F.; Sun, S.; Ouyang, X.; Wallace, C.; Zhang, Y.; Garret-Mayer, E.; Wu, J.; Liu, B.; et al. Immune chaperone gp96 drives the contributions of macrophages to inflammatory colon tumorigenesis. Cancer Res. 2014, 74, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, X.; Chen, X.; Li, Y.; Ke, Z.; Tang, T.; Chai, H.; Guo, A.M.; Chen, H.; Yang, J. Isoliquiritigenin, a flavonoid from licorice, blocks M2 macrophage polarization in colitis-associated tumorigenesis through downregulating PGE2 and IL-6. Toxicol. Appl. Pharmacol. 2014, 279, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Roncucci, L.; Mora, E.; Mariani, F.; Bursi, S.; Pezzi, A.; Rossi, G.; Pedroni, M.; Luppi, D.; Santoro, L.; Monni, S.; et al. Myeloperoxidase-positive cell infiltration in colorectal carcinogenesis as indicator of colorectal cancer risk. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2291–2297. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Popivanova, B.K.; Kostadinova, F.I.; Furuichi, K.; Shamekh, M.M.; Kondo, T.; Wada, T.; Egashira, K.; Mukaida, N. Blockade of a chemokine, CCL2, reduces chronic colitis-associated carcinogenesis in mice. Cancer Res. 2009, 69, 7884–7892. [Google Scholar] [CrossRef] [PubMed]
- Horemans, T.; Boulet, G.; van Kerckhoven, M.; Bogers, J.; Thys, S.; Vervaet, C.; Vervaeck, A.; Delputte, P.; Maes, L.; Cos, P. In Vivo evaluation of apocynin for prevention of Helicobacter pylori-induced gastric carcinogenesis. Eur. J. Cancer Prev. 2017, 26, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.H.; Niu, Y.B.; Sun, Y.; Guo, Z.J.; Li, Q.; Li, C.; Feng, J.; Cao, S.S.; Mei, Q.B. An apple oligogalactan prevents against inflammation and carcinogenesis by targeting LPS/TLR4/NF-κB pathway in a mouse model of colitis-associated colon cancer. Carcinogenesis 2010, 31, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Onuma, K.; Kanda, Y.; Suzuki Ikeda, S.; Sakaki, R.; Nonomura, T.; Kobayashi, M.; Osaki, M.; Shikanai, M.; Kobayashi, H.; Okada, F. Fermented brown rice and rice bran with Aspergillus oryzae (FBRA) prevents inflammation-related carcinogenesis in mice, through inhibition of inflammatory cell infiltration. Nutrients 2015, 7, 10237–10250. [Google Scholar] [CrossRef] [PubMed]
- Sliva, D.; Loganathan, J.; Jiang, J.; Jedinak, A.; Lamb, J.G.; Terry, C.; Baldridge, L.A.; Adamec, J.; Sandusky, G.E.; Dudhgaonkar, S. Mushroom Ganoderma lucidum prevents colitis-associated carcinogenesis in mice. PLoS ONE 2012, 7, e47873. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.A.; Chen, H.C.; Yen, G.C. The preventive role of breadfruit against inflammation-associated epithelial carcinogenesis in mice. Mol. Nutr. Food Res. 2014, 58, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Im, S.A.; Kim, J.W.; Kim, H.S.; Park, C.S.; Shin, E.; Do, S.G.; Park, Y.I.; Lee, C.K. Prevention of azoxymethane/dextran sodium sulfate-induced mouse colon carcinogenesis by processed Aloe vera gel. Int. Immunopharmacol. 2016, 40, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Hao, X.; Lee, M.J.; Lambert, J.D.; Lu, G.; Xiao, H.; Newmark, H.L.; Yang, C.S. A γ-tocopherol-rich mixture of tocopherols inhibits colon inflammation and carcinogenesis in azoxymethane and dextran sulfate sodium-treated mice. Cancer Prev. Res. 2009, 2, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Onuma, K.; Suenaga, Y.; Sakaki, R.; Yoshitome, S.; Sato, Y.; Ogawara, S.; Suzuki, S.; Kuramitsu, Y.; Yokoyama, H.; Murakami, A.; et al. Development of a quantitative bioassay to assess preventive compounds against inflammation-based carcinogenesis. Nitric Oxide 2011, 25, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Tsukamoto, T.; Seki, T.; Tanaka, H.; Morimura, S.; Cao, L.; Mizoshita, T.; Ban, H.; Toyoda, T.; Maeda, H.; et al. 4-Vinyl-2,6-dimethoxyphenol (canolol) suppresses oxidative stress and gastric carcinogenesis in Helicobacter pylori-infected carcinogen-treated Mongolian gerbils. Int. J. Cancer 2008, 122, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Wang, Y.; Liu, C.; Wang, H.; Fan, H.; Li, Y.; Wang, J.; Zhang, X.; Lu, J.; Ji, H.; et al. Chemopreventive activity of GEN-27, a genistein derivative, in colitis-associated cancer is mediated by p65-CDX2-β-catenin axis. Oncotarget 2016, 7, 17870–17884. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.C.; Siddiqui, M.S.; Athar, M.; Alam, M.S. Geraniol inhibits murine skin tumorigenesis by modulating COX-2 expression, Ras-ERK1/2 signaling pathway and apoptosis. J. Appl. Toxicol. 2013, 33, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Lai, C.S.; Tsai, C.Y.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Inotilone suppresses phorbol ester-induced inflammation and tumor promotion in mouse skin. Mol. Nutr. Food Res. 2012, 56, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Viennois, E.; Xiao, B.; Ayyadurai, S.; Wang, L.; Wang, P.G.; Zhang, Q.; Chen, Y.; Merlin, D. Micheliolide, a new sesquiterpene lactone that inhibits intestinal inflammation and colitis-associated cancer. Lab. Investig. 2014, 94, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Nakamura, Y.; Torikai, K.; Tanaka, T.; Koshiba, T.; Koshimizu, K.; Kuwahara, S.; Takahashi, Y.; Ogawa, K.; Yano, M.; et al. Inhibitory effect of citrus nobiletin on phorbol ester-induced skin inflammation, oxidative stress, and tumor promotion in mice. Cancer Res. 2000, 60, 5059–5066. [Google Scholar] [PubMed]
- Meeker, S.; Seamons, A.; Paik, J.; Treuting, P.M.; Brabb, T.; Grady, W.M.; Maggio-Price, L. Increased dietary vitamin D suppresses MAPK signaling, colitis, and colon cancer. Cancer Res. 2014, 74, 4398–4408. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Seril, D.N.; Yang, A.L.; Lu, G.G.; Yang, G.Y. Inhibition of chronic ulcerative colitis associated adenocarcinoma development in mice by inositol compounds. Carcinogenesis 2007, 28, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.T.; Lin, Y.T.; Tseng, R.Y.; Shun, C.T.; Lin, Y.C.; Wu, M.S.; Fang, J.M.; Chen, C.C. Prevention of colitis and colitis-associated colorectal cancer by a novel polypharmacological histone deacetylase inhibitor. Clin Cancer Res. 2016, 22, 4158–4169. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.P.; Jena, G.B.; Tikoo, K.B.; Kumar, V. Melatonin modulated autophagy and Nrf2 signaling pathways in mice with colitis-associated colon carcinogenesis. Mol. Carcinog. 2016, 55, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, N.; Wang, S.; Hong, J.; Fang, M.; Yousselfson, J.; Yang, P.; Newman, R.A.; Lubet, R.A.; Yang, C.S. Aberrant arachidonic acid metabolism in esophageal adenocarcinogenesis, and the effects of sulindac, nordihydroguaiaretic acid, and α-difluoromethylornithine on tumorigenesis in a rat surgical model. Carcinogenesis 2002, 23, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Doulberis, M.; Angelopoulou, K.; Kaldrymidou, E.; Tsingotjidou, A.; Abas, Z.; Erdman, S.E.; Poutahidis, T. Cholera-toxin suppresses carcinogenesis in a mouse model of inflammation-driven sporadic colon cancer. Carcinogenesis 2015, 36, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, H.; Kinjo, T.; Klinman, D.M. Effect of suppressive oligodeoxynucleotides on the development of inflammation-induced papillomas. Cancer Prev. Res. 2011, 4, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Nishizawa, T.; Ohigashi, H.; Tanaka, T.; Hou, D.X.; Colburn, N.H.; Murakami, A. Linoleic acid metabolite suppresses skin inflammation and tumor promotion in mice: Possible roles of programmed cell death 4 induction. Carcinogenesis 2009, 30, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Fujii, H.; Kimura, F.; Mishima, T.; Murata, J.; Tazawa, K.; Fujimaki, M.; Okada, F.; Hosokawa, M.; Saiki, I. Inhibitory effect of a traditional Chinese medicine, Juzen-taiho-to, on progressive growth of weakly malignant clone cells derived from murine fibrosarcoma. Jpn. J. Cancer Res. 1996, 87, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Yum, H.W.; Zhong, X.; Park, J.; Na, H.K.; Kim, N.; Lee, H.S.; Surh, Y.J. Oligonol inhibits dextran sulfate sodium-induced colitis and colonic adenoma formation in mice. Antioxid. Redox Signal. 2013, 19, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gu, X.; Robbins, D.; Li, G.; Shi, R.; McCord, J.M.; Zhao, Y. Protandim, a fundamentally new antioxidant approach in chemoprevention using mouse two-stage skin carcinogenesis as a model. PLoS ONE 2009, 4, e5284. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Takabayashi, S.; Osawa, T.; Nakamura, Y. Benzyl isothiocyanate inhibits excessive superoxide generation in inflammatory leukocytes: Implication for prevention against inflammation-related carcinogenesis. Carcinogenesis 2004, 25, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Y.; Li, R.H.; Huang, K.; Tan, G.; Li, C.; Zhi, F.C. Increased expression and possible role of chitinase 3-like-1 in a colitis-associated carcinoma model. World J. Gastroenterol. 2014, 20, 15736–15744. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Tung, N.H.; Shoyama, Y.; Sugie, S.; Mori, T.; Tanaka, T. Dietary crocin inhibits colitis and colitis-associated colorectal carcinogenesis in male ICR mice. Evid. Based Complement. Altern. Med. eCAM 2012, 2012, 820415. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.L.; Khor, T.O.; Huang, M.T.; Kong, A.N. Differential in vivo mechanism of chemoprevention of tumor formation in azoxymethane/dextran sodium sulfate mice by PEITC and DBM. Carcinogenesis 2010, 31, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cai, X.; Yang, J.; Sun, X.; Hu, C.; Yan, Z.; Xu, X.; Lu, W.; Wang, X.; Cao, P. Chemoprevention of dietary digitoflavone on colitis-associated colon tumorigenesis through inducing Nrf2 signaling pathway and inhibition of inflammation. Mol. Cancer 2014, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; de Azevedo, M.B.; Duran, N.; Alderete, J.B.; Epifano, F.; Genovese, S.; Tanaka, M.; Tanaka, T.; Curini, M. Colorectal cancer chemoprevention by 2β-cyclodextrin inclusion compounds of auraptene and 4’-geranyloxyferulic acid. Int. J. Cancer 2010, 126, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Shen, C.; Tao, Y.; Wang, S.; Wei, Z.; Cao, Y.; Wu, H.; Fan, F.; Lin, C.; Shan, Y.; et al. Chemopreventive efficacy of menthol on carcinogen-induced cutaneous carcinoma through inhibition of inflammation and oxidative stress in mice. Food Chem. Toxicol. 2015, 82, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Kuno, T.; Hatano, Y.; Tomita, H.; Hara, A.; Hirose, Y.; Hirata, A.; Mori, H.; Terasaki, M.; Masuda, S.; Tanaka, T. Organomagnesium suppresses inflammation-associated colon carcinogenesis in male Crj: CD-1 mice. Carcinogenesis 2013, 34, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Shionoya, H.; Kobayashi, M.; Kobayashi, T.; Tazawa, H.; Onuma, K.; Iuchi, Y.; Matsubara, N.; Ijichi, T.; Dugas, B.; et al. Prevention of inflammation-mediated acquisition of metastatic properties of benign mouse fibrosarcoma cells by administration of an orally available superoxide dismutase. Br. J. Cancer 2006, 94, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Habelhah, H.; Okada, F.; Nakai, K.; Choi, S.K.; Hamada, J.; Kobayashi, M.; Hosokawa, M. Polysaccharide K induces Mn superoxide dismutase (Mn-SOD) in tumor tissues and inhibits malignant progression of QR-32 tumor cells: Possible roles of interferon α, tumor necrosis factor α and transforming growth factor β in Mn-SOD induction by polysaccharide K. Cancer Immunol. Immunother. 1998, 46, 338–344. [Google Scholar] [PubMed]
- Khan, A.Q.; Khan, R.; Tahir, M.; Rehman, M.U.; Lateef, A.; Ali, F.; Hamiza, O.O.; Hasan, S.K.; Sultana, S. Silibinin inhibits tumor promotional triggers and tumorigenesis against chemically induced two-stage skin carcinogenesis in Swiss albino mice: Possible role of oxidative stress and inflammation. Nutr. Cancer 2014, 66, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Jiang, Z.; Hall, Y.J.; Jang, Y.; Snyder, P.W.; Bain, C.; Huang, J.; Jannasch, A.; Cooper, B.; Wang, Y.; et al. γ-Tocopherol attenuates moderate but not severe colitis and suppresses moderate colitis-promoted colon tumorigenesis in mice. Free Radic. Biol. Med. 2013, 65, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kwon, H.S.; Kim, D.H.; Shin, E.K.; Kang, Y.H.; Park, J.H.; Shin, H.K.; Kim, J.K. 3,3′-diindolylmethane attenuates colonic inflammation and tumorigenesis in mice. Inflamm. Bowel Dis. 2009, 15, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Xi, M.Y.; Jia, J.M.; Sun, H.P.; Sun, Z.Y.; Jiang, J.W.; Wang, Y.J.; Zhang, M.Y.; Zhu, J.F.; Xu, L.L.; Jiang, Z.Y.; et al. 3-aroylmethylene-2,3,6,7-tetrahydro-1H-pyrazino[2,1-a]isoquinolin-4(11bH)-ones as potent Nrf2/ARE inducers in human cancer cells and AOM-DSS treated mice. J. Med. Chem. 2013, 56, 7925–7938. [Google Scholar] [CrossRef] [PubMed]
- Hamada, J.; Nakata, D.; Nakae, D.; Kobayashi, Y.; Akai, H.; Konishi, Y.; Okada, F.; Shibata, T.; Hosokawa, M.; Moriuchi, T. Increased oxidative DNA damage in mammary tumor cells by continuous epidermal growth factor stimulation. J. Natl. Cancer Inst. 2001, 93, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Fidalgo, S.; Villegas, I.; Cardeno, A.; Talero, E.; Sanchez-Hidalgo, M.; Motilva, V.; Alarcon de la Lastra, C. Extra-virgin olive oil-enriched diet modulates DSS-colitis-associated colon carcinogenesis in mice. Clin. Nutr. 2010, 29, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Phutthaphadoong, S.; Yamada, Y.; Hirata, A.; Tomita, H.; Hara, A.; Limtrakul, P.; Iwasaki, T.; Kobayashi, H.; Mori, H. Chemopreventive effect of fermented brown rice and rice bran (FBRA) on the inflammation-related colorectal carcinogenesis in ApcMin/+ mice. Oncol. Rep. 2010, 23, 53–59. [Google Scholar] [PubMed]
- Yasui, Y.; Hosokawa, M.; Mikami, N.; Miyashita, K.; Tanaka, T. Dietary astaxanthin inhibits colitis and colitis-associated colon carcinogenesis in mice via modulation of the inflammatory cytokines. Chem. Biol. Interact. 2011, 193, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.Z.; Liu, C.H.; Wei, B.; Qiao, J.; Lu, T.; Wei, H.C.; Chen, H.D.; He, C.D. Baicalein inhibits DMBA/TPA-induced skin tumorigenesis in mice by modulating proliferation, apoptosis, and inflammation. Inflammation 2013, 36, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Sung, B.; Kang, Y.J.; Jang, J.Y.; Hwang, S.Y.; Lee, Y.; Kim, M.; Im, E.; Yoon, J.H.; Kim, C.M.; et al. Anti-inflammatory effects of betaine on AOM/DSS induced colon tumorigenesis in ICR male mice. Int. J. Oncol. 2014, 45, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Prakobwong, S.; Khoontawad, J.; Yongvanit, P.; Pairojkul, C.; Hiraku, Y.; Sithithaworn, P.; Pinlaor, P.; Aggarwal, B.B.; Pinlaor, S. Curcumin decreases cholangiocarcinogenesis in hamsters by suppressing inflammation-mediated molecular events related to multistep carcinogenesis. Int. J. Cancer 2011, 129, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.L.; Lai, C.S.; Chang, Y.H.; Chen, W.J.; Ho, C.T.; Pan, M.H. Pterostilbene, a natural analogue of resveratrol, potently inhibits 7,12-dimethylbenz[a]anthracene (DMBA)/12-O-tetradecanoylphorbol-13-acetate (TPA)-induced mouse skin carcinogenesis. Food Funct. 2012, 3, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Kohno, H.; Suzuki, R.; Yasui, Y.; Miyamoto, S.; Wakabayashi, K.; Tanaka, T. Ursodeoxycholic acid versus sulfasalazine in colitis-related colon carcinogenesis in mice. Clin. Cancer Res. 2007, 13, 2519–2525. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Li, S.; Chai, C.Y.; Lo, C.Y.; Ho, C.T.; Wang, Y.J.; Pan, M.H. Inhibitory effect of citrus 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone on 12-O-tetradecanoylphorbol 13-acetate-induced skin inflammation and tumor promotion in mice. Carcinogenesis 2007, 28, 2581–2588. [Google Scholar] [CrossRef] [PubMed]
- Kohno, H.; Suzuki, R.; Sugie, S.; Tanaka, T. Suppression of colitis-related mouse colon carcinogenesis by a COX-2 inhibitor and PPAR ligands. BMC Cancer 2005, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Kochi, T.; Shirakami, Y.; Genovese, S.; Epifano, F.; Fiorito, S.; Mori, T.; Tanaka, T.; Moriwaki, H. A newly synthesized compound, 4′-geranyloxyferulic acid-N(omega)-nitro-l-arginine methyl ester suppresses inflammation-associated colorectal carcinogenesis in male mice. Int. J. Cancer 2014, 135, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, J.S.; Hong, K.S.; Chung, J.W.; Kim, J.H.; Hahm, K.B. Novel application of proton pump inhibitor for the prevention of colitis-induced colorectal carcinogenesis beyond acid suppression. Cancer Prev. Res. 2010, 3, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Mishima, T.; Tajima, Y.; Kuroki, T.; Kosaka, T.; Adachi, T.; Kitasato, A.; Tsuneoka, N.; Kitajima, T.; Kanematsu, T. Chemopreventative effect of an inducible nitric oxide synthase inhibitor, ONO-1714, on inflammation-associated biliary carcinogenesis in hamsters. Carcinogenesis 2009, 30, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Y.; Zhang, C.; Su, Z.Y.; Li, W.; Huang, M.T.; Kong, A.N. The epigenetic effects of aspirin: The modification of histone H3 lysine 27 acetylation in the prevention of colon carcinogenesis in azoxymethane- and dextran sulfate sodium-treated CF-1 mice. Carcinogenesis 2016, 37, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wang, K.; Wang, Z.; Li, N.; Ji, G. Chemopreventive effect of dietary glutamine on colitis-associated colon tumorigenesis in mice. Carcinogenesis 2013, 34, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.S.; Choi, H.E.; Shin, J.S.; Cho, E.J.; Cho, Y.W.; Choi, J.H.; Baek, N.I.; Lee, K.T. Chemopreventive effects of standardized ethanol extract from the aerial parts of Artemisia princeps Pampanini cv. Sajabal via NF-κB inactivation on colitis-associated colon tumorigenesis in mice. Food Chem. Toxicol. 2015, 75, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Lavi, I.; Nimri, L.; Levinson, D.; Peri, I.; Hadar, Y.; Schwartz, B. Glucans from the edible mushroom Pleurotus pulmonarius inhibit colitis-associated colon carcinogenesis in mice. J. Gastroenterol. 2012, 47, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.; Yasui, Y.; Sugie, S.; Koketsu, M.; Watanabe, K.; Tanaka, T. Dietary tricin suppresses inflammation-related colon carcinogenesis in male Crj: CD-1 mice. Cancer Prev. Res. 2009, 2, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Kangwan, N.; Kim, Y.J.; Han, Y.M.; Jeong, M.; Park, J.M.; Go, E.J.; Hahm, K.B. Sonic hedgehog inhibitors prevent colitis-associated cancer via orchestrated mechanisms of IL-6/gp130 inhibition, 15-PGDH induction, Bcl-2 abrogation, and tumorsphere inhibition. Oncotarget 2016, 7, 7667–7682. [Google Scholar] [PubMed]
- Polytarchou, C.; Hommes, D.W.; Palumbo, T.; Hatziapostolou, M.; Koutsioumpa, M.; Koukos, G.; van der Meulen-de Jong, A.E.; Oikonomopoulos, A.; van Deen, W.K.; Vorvis, C.; et al. MicroRNA214 is associated with progression of ulcerative colitis, and inhibition reduces development of colitis and colitis-associated cancer in mice. Gastroenterology 2015, 149, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, F.; Wang, Y.; Cai, M.; Wang, Q.; Guo, Q.; Li, Z.; Hu, R. Oroxylin A inhibits colitis-associated carcinogenesis through modulating the IL-6/STAT3 signaling pathway. Inflamm. Bowel Dis. 2013, 19, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, H.; Xu, R.; Mei, Q.; Fan, D. Triptolide downregulates Rac1 and the JAK/STAT3 pathway and inhibits colitis-related colon cancer progression. Exp. Mol. Med. 2009, 41, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Altamemi, I.; Murphy, E.A.; Catroppo, J.F.; Zumbrun, E.E.; Zhang, J.; McClellan, J.L.; Singh, U.P.; Nagarkatti, P.S.; Nagarkatti, M. Role of microRNAs in resveratrol-mediated mitigation of colitis-associated tumorigenesis in ApcMin/+ mice. J. Pharmacol. Exp. Ther. 2014, 350, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, J.; Jackson, A.L.; Clark, L.N.; Kilgore, J.; Guo, H.; Livingston, N.; Batchelor, K.; Yin, Y.; Gilliam, T.P.; et al. NT1014, a novel biguanide, inhibits ovarian cancer growth in vitro and in vivo. J. Hematol. Oncol. 2016, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Toyoda, T.; Tsukamoto, T.; Mori, A.; Tatematsu, M.; Ushijima, T. Prevention of Helicobacter pylori-induced gastric cancers in gerbils by a DNA demethylating agent. Cancer Prev. Res. 2013, 6, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Takai, S.; Hosono, A.; Seki, T. Bovine milk-derived α-lactalbumin inhibits colon inflammation and carcinogenesis in azoxymethane and dextran sodium sulfate-treated mice. Biosci. Biotechnol. Biochem. 2014, 78, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Piazzi, G.; D’Argenio, G.; Prossomariti, A.; Lembo, V.; Mazzone, G.; Candela, M.; Biagi, E.; Brigidi, P.; Vitaglione, P.; Fogliano, V.; et al. Eicosapentaenoic acid free fatty acid prevents and suppresses colonic neoplasia in colitis-associated colorectal cancer acting on Notch signaling and gut microbiota. Int. J. Cancer 2014, 135, 2004–2013. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Hu, H.M.; Tsai, P.Y.; Wu, I.C.; Yang, S.F.; Chang, L.L.; Wang, J.Y.; Jan, C.M.; Wang, W.M.; Wu, D.C. Short-term celecoxib intervention is a safe and effective chemopreventive for gastric carcinogenesis based on a Mongolian gerbil model. World J. Gastroenterol. 2009, 15, 4907–4914. [Google Scholar] [CrossRef] [PubMed]
- Buttar, N.S.; Wang, K.K.; Leontovich, O.; Westcott, J.Y.; Pacifico, R.J.; Anderson, M.A.; Krishnadath, K.K.; Lutzke, L.S.; Burgart, L.J. Chemoprevention of esophageal adenocarcinoma by COX-2 inhibitors in an animal model of Barrett’s esophagus. Gastroenterology 2002, 122, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. Rate of generation of oxidative stress-related damage and animal longevity. Free Radic. Biol. Med. 2002, 33, 1167–1172. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.J. Emerging avenues linking inflammation and cancer. Free Radic. Biol. Med. 2012, 52, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008, 275, 3249–3277. [Google Scholar] [CrossRef] [PubMed]
- Davtyan, T.K.; Manukyan, H.M.; Hakopyan, G.S.; Mkrtchyan, N.R.; Avetisyan, S.A.; Galoyan, A.A. Hypothalamic proline-rich polypeptide is an oxidative burst regulator. Neurochem. Res. 2005, 30, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Kobayashi, M.; Tanaka, H.; Kobayashi, T.; Tazawa, H.; Iuchi, Y.; Onuma, K.; Hosokawa, M.; Dinauer, M.C.; Hunt, N.H. The role of nicotinamide adenine dinucleotide phosphate oxidase-derived reactive oxygen species in the acquisition of metastatic ability of tumor cells. Am. J. Pathol. 2006, 169, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.; Gansauge, F.; Beger, H.; Dolara, P.; Winde, G.; Bartsch, H. Increased etheno-DNA adducts in affected tissues of patients suffering from Crohn’s disease, ulcerative colitis, and chronic pancreatitis. Antioxid. Redox Signal. 2006, 8, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Ambs, S.; Merriam, W.G.; Bennett, W.P.; Felley-Bosco, E.; Ogunfusika, M.O.; Oser, S.M.; Klein, S.; Shields, P.G.; Billiar, T.R.; Harris, C.C. Frequent nitric oxide synthase-2 expression in human colon adenomas: Implication for tumor angiogenesis and colon cancer progression. Cancer Res. 1998, 58, 334–341. [Google Scholar] [PubMed]
- Wilson, K.T.; Fu, S.; Ramanujam, K.S.; Meltzer, S.J. Increased expression of inducible nitric oxide synthase and cyclooxygenase-2 in Barrett’s esophagus and associated adenocarcinomas. Cancer Res. 1998, 58, 2929–2934. [Google Scholar] [PubMed]
- Sawa, T.; Ohshima, H. Nitrative DNA damage in inflammation and its possible role in carcinogenesis. Nitric Oxide 2006, 14, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Nakai, K.; Kobayashi, T.; Shibata, T.; Tagami, S.; Kawakami, Y.; Kitazawa, T.; Kominami, R.; Yoshimura, S.; Suzuki, K.; et al. Inflammatory cell-mediated tumour progression and minisatellite mutation correlate with the decrease of antioxidative enzymes in murine fibrosarcoma cells. Br. J. Cancer 1999, 79, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Korantzopoulos, P.; Kolettis, T.M.; Kountouris, E.; Dimitroula, V.; Karanikis, P.; Pappa, E.; Siogas, K.; Goudevenos, J.A. Oral vitamin C administration reduces early recurrence rates after electrical cardioversion of persistent atrial fibrillation and attenuates associated inflammation. Int. J. Cardiol. 2005, 102, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lykkesfeldt, J.; Shigenaga, M.K.; Shigeno, E.T.; Christen, S.; Ames, B.N. γ-tocopherol supplementation inhibits protein nitration and ascorbate oxidation in rats with inflammation. Free Radic. Biol. Med. 2002, 33, 1534–1542. [Google Scholar] [CrossRef]
- Christen, S.; Woodall, A.A.; Shigenaga, M.K.; Southwell-Keely, P.T.; Duncan, M.W.; Ames, B.N. γ-Tocopherol traps mutagenic electrophiles such as NOX and complements α-tocopherol: Physiological implications. Proc. Natl. Acad. Sci. USA 1997, 94, 3217–3222. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A.; Xu, Z.M. Minimizing rancidity in muscle foods. Food Technol. 1998, 52, 54–59. [Google Scholar]
- Kim, H.J.; Kim, M.K.; Chang, W.K.; Choi, H.S.; Choi, B.Y.; Lee, S.S. Effect of nutrient intake and Helicobacter pylori infection on gastric cancer in Korea: A case-control study. Nutr. Cancer 2005, 52, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Tazawa, H.; Sylla, B.S.; Sawa, T. Prevention of human cancer by modulation of chronic inflammatory processes. Mutat. Res. 2005, 591, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Ouyang, W.; Huang, C. Inflammation, a key event in cancer development. Mol. Cancer Res. 2006, 4, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Schuliga, M. NF-κB signaling in chronic inflammatory airway disease. Biomolecules 2015, 5, 1266–1283. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Solt, L.A.; May, M.J. The IκB kinase complex: Master regulator of NF-κB signaling. Immunol. Res. 2008, 42, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS crosstalk in inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Kagoya, Y.; Yoshimi, A.; Kataoka, K.; Nakagawa, M.; Kumano, K.; Arai, S.; Kobayashi, H.; Saito, T.; Iwakura, Y.; Kurokawa, M. Positive feedback between NF-κB and TNF-α promotes leukemia-initiating cell capacity. J. Clin. Investig. 2014, 124, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.F.; Hu, T.L.; Soutto, M.; Belkhiri, A.; El-Rifai, W. Loss of glutathione peroxidase 7 promotes TNF-α-induced NF-κB activation in Barrett’s carcinogenesis. Carcinogenesis 2014, 35, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Khatami, M.; Baglole, C.J.; Sun, J.; Harris, S.A.; Moon, E.Y.; Al-Mulla, F.; Al-Temaimi, R.; Brown, D.G.; Colacci, A.; et al. Environmental immune disruptors, inflammation and cancer risk. Carcinogenesis 2015, 36 (Suppl. S1), S232–S253. [Google Scholar] [CrossRef] [PubMed]
- Schetter, A.J.; Heegaard, N.H.; Harris, C.C. Inflammation and cancer: Interweaving microRNA, free radical, cytokine and p53 pathways. Carcinogenesis 2010, 31, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Riva, F.; Bonavita, E.; Barbati, E.; Muzio, M.; Mantovani, A.; Garlanda, C. TIR8/SIGIRR is an interleukin-1 receptor/toll like receptor family member with regulatory functions in inflammation and immunity. Front. Immunol. 2012, 3, 322. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.J.; Owens, D.M.; Stamp, G.; Arnott, C.; Burke, F.; East, N.; Holdsworth, H.; Turner, L.; Rollins, B.; Pasparakis, M.; et al. Mice deficient in tumor necrosis factor-α are resistant to skin carcinogenesis. Nat. Med. 1999, 5, 828–831. [Google Scholar] [PubMed]
- Yoshida, G.J. Therapeutic strategies of drug repositioning targeting autophagy to induce cancer cell death: From pathophysiology to treatment. J. Hematol. Oncol. 2017, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fu, H.; Nanayakkara, G.; Li, Y.; Shao, Y.; Johnson, C.; Cheng, J.; Yang, W.Y.; Yang, F.; Lavallee, M.; et al. Novel extracellular and nuclear caspase-1 and inflammasomes propagate inflammation and regulate gene expression: A comprehensive database mining study. J. Hematol. Oncol. 2016, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zhang, J. Inflammasomes in inflammation-induced cancer. Front. Immunol. 2017, 8, 271. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, M.; Casolaro, V.; Pinto, A.; Sorrentino, R. Inflammasome: Cancer’s friend or foe? Pharmacol. Ther. 2014, 143, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Michopoulou, A.; Rousselle, P. How do epidermal matrix metalloproteinases support re-epithelialization during skin healing? Eur. J. Dermatol. 2015, 25, 33–42. [Google Scholar] [PubMed]
- Knight, D. Epithelium-fibroblast interactions in response to airway inflammation. Immunol. Cell Biol. 2001, 79, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Hopkinson-Woolley, J.; McCluskey, J. Growth factors and cutaneous wound repair. Prog. Growth Factor Res. 1992, 4, 25–44. [Google Scholar] [CrossRef]
- Nagayasu, H.; Hamada, J.; Nakata, D.; Shibata, T.; Kobayashi, M.; Hosokawa, M.; Takeichi, N. Reversible and irreversible tumor progression of a weakly malignant rat mammary carcinoma cell line by in vitro exposure to epidermal growth factor. Int. J. Oncol. 1998, 12, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Madan, R.; Karp, C.L.; Braciale, T.J. Effector T cells control lung inflammation during acute influenza virus infection by producing IL-10. Nat. Med. 2009, 15, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Schottelius, A.J.; Mayo, M.W.; Sartor, R.B.; Baldwin, A.S., Jr. Interleukin-10 signaling blocks inhibitor of κB kinase activity and nuclear factor κB DNA binding. J. Biol. Chem. 1999, 274, 31868–31874. [Google Scholar] [CrossRef] [PubMed]
- Sturlan, S.; Oberhuber, G.; Beinhauer, B.G.; Tichy, B.; Kappel, S.; Wang, J.; Rogy, M.A. Interleukin-10-deficient mice and inflammatory bowel disease associated cancer development. Carcinogenesis 2001, 22, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Ryu, K.J.; Hong, M.; Ko, Y.H.; Kim, W.S. The serum CXCL13 level is associated with the Glasgow Prognostic Score in extranodal NK/T-cell lymphoma patients. J. Hematol. Oncol. 2015, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Reinecker, H.C.; Loh, E.Y.; Ringler, D.J.; Mehta, A.; Rombeau, J.L.; MacDermott, R.P. Monocyte-chemoattractant protein 1 gene expression in intestinal epithelial cells and inflammatory bowel disease mucosa. Gastroenterology 1995, 108, 40–50. [Google Scholar] [CrossRef]
- Nasrallah, R.; Hassouneh, R.; Hebert, R.L. PGE2, kidney disease, and cardiovascular risk: Beyond hypertension and diabetes. J. Am. Soc. Nephrol. 2016, 27, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Miyaura, C.; Inada, M.; Matsumoto, C.; Ohshiba, T.; Uozumi, N.; Shimizu, T.; Ito, A. An essential role of cytosolic phospholipase A2α in prostaglandin E2-mediated bone resorption associated with inflammation. J. Exp. Med. 2003, 197, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Sahin, M.; Sahin, E.; Gumuslu, S. Cyclooxygenase-2 in cancer and angiogenesis. Angiology 2009, 60, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.W.; Luo, F.; Cheng, H.; Zhao, J.J.; Liu, P. Chemopreventive effects of aspirin at a glance. Biochim. Biophys. Acta 2015, 1855, 254–263. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Cancer-Preventive Agents; International Agency for Research on Cancer. Non-Steroidal Anti-Inflammatory Drugs; International Agency for Research on Cancer: Lyon, France, 1997. [Google Scholar]
- Flossmann, E.; Rothwell, P.M. British Doctors Aspirin Trial and the UK-TIA Aspirin Trial. Effect of aspirin on long-term risk of colorectal cancer: Consistent evidence from randomised and observational studies. Lancet 2007, 369, 1603–1613. [Google Scholar] [CrossRef]
- Cuzick, J.; Otto, F.; Baron, J.A.; Brown, P.H.; Burn, J.; Greenwald, P.; Jankowski, J.; La Vecchia, C.; Meyskens, F.; Senn, H.J.; et al. Aspirin and non-steroidal anti-inflammatory drugs for cancer prevention: An international consensus statement. Lancet Oncol. 2009, 10, 501–507. [Google Scholar] [CrossRef]
- Harris, R.E.; Casto, B.C.; Harris, Z.M. Cyclooxygenase-2 and the inflammogenesis of breast cancer. World J. Clin. Oncol. 2014, 5, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Knaus, E.E. Evolution of nonsteroidal anti-inflammatory drugs (NSAIDs): Cyclooxygenase (COX) inhibition and beyond. J. Pharm. Pharm. Sci. 2008, 11, 81s–110s. [Google Scholar] [CrossRef] [PubMed]
- Fowler, T.O.; Durham, C.O.; Planton, J.; Edlund, B.J. Use of nonsteroidal anti-inflammatory drugs in the older adult. J. Am. Assoc. Nurse Pract. 2014, 26, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.M.; Richardson, P.; El-Serag, H.B. Medications (NSAIDs, statins, proton pump inhibitors) and the risk of esophageal adenocarcinoma in patients with Barrett’s esophagus. Gastroenterology 2010, 138, 2260–2266. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.; Collado, M.; Navas, C.; Schuhmacher, A.J.; Hernandez-Porras, I.; Canamero, M.; Rodriguez-Justo, M.; Serrano, M.; Barbacid, M. Pancreatitis-induced inflammation contributes to pancreatic cancer by inhibiting oncogene-induced senescence. Cancer Cell 2011, 19, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.J.; Ollila, C.A.; Kumar, A.; Borresen, E.C.; Raina, K.; Agarwal, R.; Ryan, E.P. Chemopreventive properties of dietary rice bran: Current status and future prospects. Adv. Nutr. 2012, 3, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Kelloff, G.J.; Crowell, J.A.; Steele, V.E.; Lubet, R.A.; Malone, W.A.; Boone, C.W.; Kopelovich, L.; Hawk, E.T.; Lieberman, R.; Lawrence, J.A.; et al. Progress in cancer chemoprevention: Development of diet-derived chemopreventive agents. J. Nutr. 2000, 130 (Suppl. S2), S467–S471. [Google Scholar] [CrossRef]
- Thun, M.J.; DeLancey, J.O.; Center, M.M.; Jemal, A.; Ward, E.M. The global burden of cancer: Priorities for prevention. Carcinogenesis 2010, 31, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zheng, Z.; Ruan, J.; Li, Z.; Tzeng, C.M. Chronic inflammation links cancer and Parkinson’s disease. Front. Aging Neurosci. 2016, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Levy Nogueira, M.; da Veiga Moreira, J.; Baronzio, G.F.; Dubois, B.; Steyaert, J.M.; Schwartz, L. Mechanical stress as the common denominator between chronic inflammation, cancer, and Alzheimer’s disease. Front. Oncol. 2015, 5, 197. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Goldeck, D.; Derhovanessian, E. Inflammation, ageing and chronic disease. Curr. Opin. Immunol. 2014, 29, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.E. Inflammation, depression and dementia: Are they connected? Neurochem. Res. 2007, 32, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Stange, E.F.; Wehkamp, J. Recent advances in understanding and managing Crohn’s disease. F1000Res. 2016, 5, 2896. [Google Scholar] [CrossRef] [PubMed]
- Hajebrahimi, B.; Kiamanesh, A.; Asgharnejad Farid, A.A.; Asadikaram, G. Type 2 diabetes and mental disorders; a plausible link with inflammation. Cell. Mol. Biol. 2016, 62, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Fougere, B.; Boulanger, E.; Nourhashemi, F.; Guyonnet, S.; Cesari, M. Chronic inflammation: Accelerator of biological aging. J. Gerontol. A Biol. Sci. Med. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Viola, J.; Soehnlein, O. Atherosclerosis—A matter of unresolved inflammation. Semin. Immunol. 2015, 27, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoogen, P.; van den Akker, F.; Deddens, J.C.; Sluijter, J.P. Heart failure in chronic myocarditis: A role for microRNAs? Curr. Genom. 2015, 16, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Podolska, M.J.; Biermann, M.H.; Maueroder, C.; Hahn, J.; Herrmann, M. Inflammatory etiopathogenesis of systemic lupus erythematosus: An update. J. Inflamm. Res. 2015, 8, 161–171. [Google Scholar] [PubMed]
- De Souza, A.W.; de Carvalho, J.F. Diagnostic and classification criteria of Takayasu arteritis. J. Autoimmun. 2014, 48–49, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Zhernakova, A.; Withoff, S.; Wijmenga, C. Clinical implications of shared genetics and pathogenesis in autoimmune diseases. Nat. Rev. Endocrinol. 2013, 9, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef] [PubMed]
- David, J.; Ansell, B.M.; Woo, P. Polyarteritis nodosa associated with streptococcus. Arch. Dis. Child. 1993, 69, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Hirose, N.; Arai, Y.; Gondoh, Y.; Nakazawa, S.; Takayama, M.; Ebihara, Y.; Shimizu, K.; Inagaki, H.; Masui, Y.; Kitagawa, K.; et al. Tokyo centenarian study: Aging inflammation hypothesis. Geriatr. Gerontol. Int. 2004, 4, S182–S185. [Google Scholar] [CrossRef]
- Kamp, D.W.; Shacter, E.; Weitzman, S.A. Chronic inflammation and cancer: The role of the mitochondria. Oncology 2011, 25, 400–413. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites of Inflammation-Related Carcinogenesis | Causes of Inflammation/Pathological Condition | ||
|---|---|---|---|
| Definitely Carcinogenic Agents (Group 1) | Presumed Carcinogenic Agents (Group 2A to 3 and the Others) | References | |
| Eye | HIV type 1 | [8] | |
| UV-associated skin inflammation | [8] | ||
| Lip | UV-associated skin inflammation | [8] | |
| Oral cavity | HPV type 16 | [8] | |
| HPV type 18 | [8] | ||
| Gingivitis | [9] | ||
| Lichen planus | [9] | ||
| Leukoplakia | [10] | ||
| Periodontitis | [11] | ||
| Salivary gland | Sialadenitis | [9] | |
| Tongue | HPV | [12] | |
| Caries | [13] | ||
| Tonsil | HPV type 16 | [8,12] | |
| Nasopharynx | EBV | [8,10,12] | |
| Pharynx | HPV type 16 | [8] | |
| Asbestos | [8] | ||
| Oropharynx | HPV | [12] | |
| Larynx | Asbestos | [8] | |
| HPV type 16 | [8] | ||
| Thyroid | Chronic lymphocytic thyroiditis | [14] | |
| Hashimoto’s thyroiditis | [14] | ||
| Esophagus | Gastric reflux, esophagitis | [9,10] | |
| Barrett’s esophagus | [10] | ||
| Barrett’s metaplasia | [9] | ||
| Neisseria mucosa | [15] | ||
| Neisseria sicca | [15] | ||
| Neisseria subflava | [15] | ||
| Lung | Asbestos | [8] | |
| Coal gasification | [8] | ||
| Outdoor air pollution | [8,10,16] | ||
| Tobacco smoke/smoking | [8,10] | ||
| Asthma | [17] | ||
| Bronchitis | [9] | ||
| COPD | [18] | ||
| Interstitial pneumonia | [19] | ||
| Sarcoidosis | [20] | ||
| Silicosis | [9] | ||
| Tuberculosis | [21] | ||
| Chlamydia pneumoniae | [22] | ||
| HPV type 16 | [23] | ||
| HIV type 1 | [24] | ||
| Lung mesothelium | Asbestos | [8,10] | |
| Silicosis | [25] | ||
| Breast | HERV-K | [26] | |
| Inflammatory breast cancer | [10] | ||
| Stomach | Helicobacter pylori | [8,10,12] | |
| Asbestos | [8] | ||
| EBV | [8,10] | ||
| Chronic atrophic gastritis | [10] | ||
| Liver | HBV | [8,10,12] | |
| HCV | [8,10,12] | ||
| Clonorchis sinensis | [8,10] | ||
| Opisthorchis viverrini | [8,10] | ||
| Cirrhosis | [10] | ||
| HDV | [27] | ||
| HIV type 1 | [8] | ||
| Schistosoma japonicum | [8,10] | ||
| Hemochromatosis | [28] | ||
| α-1-anti-trypsin deficiency | [28] | ||
| Alcohol | [28] | ||
| Bile duct | Clonorchis sinensis | [12] | |
| Opisthorchis viverrini | [12] | ||
| Primary sclerosing cholangitis | [29] | ||
| Bile acids-associated cholangitis | [9] | ||
| Gall bladder | Gall bladder stone-associated cholecystitis | [9,10] | |
| Primary sclerosing cholangitis | [29] | ||
| Pancreaticobiliary maljunction | [30] | ||
| Salmonella typhimurium | [10] | ||
| Salmonella enterica serovar Typhi | [31] | ||
| Pancreas | Chronic pancreatitis | [10] | |
| Alcoholism-associated pancreatitis | [9] | ||
| Hereditary pancreatitis | [32] | ||
| Alcohol | [33] | ||
| Colon and Rectum | Bile acids-associated coloproctitis | [9] | |
| Inflammatory bowel diseases | [9,10,34] | ||
| Cytomegalovirus | [35] | ||
| EBV | [35] | ||
| HPV | [35] | ||
| JCV | [35] | ||
| Bacteroides | [35] | ||
| Clostridium septicum | [36] | ||
| Escherichia coli | [35] | ||
| Helicobacter pylori | [35] | ||
| Streptococcus bovis | [35] | ||
| Streptococcus gallolyticus | [37] | ||
| Schistosoma japonicum | [8,10] | ||
| Asbestos | [8] | ||
| Bladder | Schistosoma haematobium | [8,10,12,38] | |
| Cystitis | [10] | ||
| Urinary catheter-associated cystitis | [9,39] | ||
| Anus | HIV type 1 | [8] | |
| HPV type 16 | [8] | ||
| HPV types 18, 33 | [8] | ||
| Anal fistula | [40] | ||
| Testis | EBV | [41] | |
| Prostate | Prostatitis | [42] | |
| Proliferative inflammatory atrophy | [10] | ||
| Gonorrhea | [43] | ||
| Trichomonas vaginalis | [44] | ||
| Ovary | Asbestos | [8] | |
| Pelvic inflammatory disease | [9] | ||
| Endometriosis | [45] | ||
| Uterine cervix | HPV types 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59 | [8] | |
| HIV type 1 | [8] | ||
| HPV types 26, 53, 66, 67, 68, 70, 73, 82 | [8] | ||
| Herpes simplex virus | [10] | ||
| Penis | HPV type 16 | [8] | |
| HIV types 1 | [8] | ||
| HPV types 18 | [8] | ||
| Vulva | HPV type 16 | [8] | |
| HIV types 1 | [8] | ||
| HPV types 18, 33 | [8] | ||
| Lichen sclerosis | [9,46] | ||
| Vagina | HPV type 16 | [8] | |
| HIV types 1 | [8] | ||
| Skin | UV-associated skin inflammation | [8,10] | |
| Chronic osteomyelitis | [47] | ||
| HIV types 1 | [8] | ||
| HPV types 5, 8 | [8] | ||
| MCV | [48] | ||
| Melanoma | UV-associated skin inflammation | [9] | |
| Non-melanomatous skin cancer | Cutaneous HPV types | [48] | |
| Central nerve | JCV | [49] | |
| Endothelium (Kaposi’s sarcoma) | HIV type 1 | [8,10] | |
| KSHV | [8] | ||
| Vasculature | Bartonella | [50] | |
| Hodgkin’s lymphoma | EBV | [12] | |
| HIV type 1 | [51] | ||
| Non-Hodgkin lymphoma | EBV | [12] | |
| HBV | [52] | ||
| HCV | [12] | ||
| HTLV-1 | [12] | ||
| Lymphoma | EBV | [8,10] | |
| HCV | [8] | ||
| HIV type 1 | [8] | ||
| HTLV-1 | [8,10] | ||
| KSHV | [8] | ||
| HIV type 2 | [53] | ||
| Hashimoto’s thyroiditis | [9] | ||
| Sjögren’s syndrome | [9] | ||
| Childhood celiac disease | [54] | ||
| HBV | [55] | ||
| HTLV-1 | [56] | ||
| Malaria | [10] | ||
| Orbital lymphoma | Chlamydia psittaci | [57] | |
| Thyroid lymphoma | Hashimoto’s thyroiditis | [58] | |
| Lymphoma in the pleural cavity | EBV | [59] | |
| Pyothorax-associated lymphoma | EBV | [60] | |
| MALT lymphoma | Helicobacter pylori | [8,12] | |
| Small-bowel lymphoma | Campylobacter jejuni | [61] | |
| Cutaneous lymphoma | Borrelia burgdorferi | [62] | |
| DLBC lymphoma | Helicobacter pylori | [12] | |
| Adult T-cell leukemia | ATL (HTLV-1) | [63] | |
| T-cell lymphoma | EBV | [64] | |
| Burkitt’s lymphoma | EBV | [65] | |
| B-cell lymphoma | EBV | [66] | |
| Primary effusion lymphoma | KSHV | [67] | |
| Treatment | Carcinogen | Animal | Arising Tumor | Reference |
|---|---|---|---|---|
| Esophagojejunostomy | None | Rat | Esophageal adenocarcinoma | [84] |
| H. pylori infection | MNNG | Mongolian gerbil | Gastric adenocarcinoma | [85] |
| DSS | None | Mouse | Colorectal adenocarcinoma | [86] |
| DSS | AOM | Mouse | Colorectal adenocarcinoma | [86] |
| DSS | DMH | Mouse | Colorectal adenocarcinoma | [87] |
| DSS | PhIP | Mouse | Colorectal adenocarcinoma | [86] |
| DSS | None | ApcMin/+ mouse | Colorectal adenocarcinoma | [87] |
| None | None | HBV-transgenic mouse | Hepatocellular carcinoma | [88] |
| None | DEN | Rat | Hepatocellular carcinoma | [89] |
| CCl4 | DEN | Mouse | Hepatocellular carcinoma | [90] |
| O. viverrini infection | NDMA | Hamster | Cholangiocarcinoma | [91] |
| Choledochojejunostomy | N-nitrosobis(2-oxopropyl)amine | Hamster | Biliary carcinoma | [92] |
| Caerulein | None | K-ras mutated mouse | Pancreatic ductal adenocarcinoma | [93] |
| TPA | DMBA | Mouse | Squamous cell carcinoma | [94] |
| Prevention Strategy | Chemopreventive Agent [Reference] | Type of Agent |
|---|---|---|
| I. Inhibition of inflammatory cell infiltration | Apocynin [143], apple oligogalactan [144], FBRA [145], Ganoderma lucidum [146], MEFA [147], MELA [147], PAG [148], γ-TmT [149] | Food product |
| Auraptene [150], canolol [151], genistein-27 [152], geraniol [153], inotilone [154], micheliolide [155], nobiletin [156], tumerone [150], vitamin D [157] | Natural compound | |
| Hexaphosphate inositol [158], inositol [158], statin hydroxamate [159] | Low-molecular weight compound | |
| Melatonin [160] | Amino acid and its derivative | |
| Sulindac [161] | COX inhibitor | |
| Cholera-toxin [162] | Protein | |
| Oligonucleotides [163] | Oligonucleotides | |
| 13-HOA [164] | Fatty acid | |
| II. Inhibition of ROS | Juzen-taiho-to [165], oligonol [166], protandim [167] | Food product |
| Auraptene [150], benzyl isothiocyanate [168], caffeine [169], crocin [170], DBM [171], digitoflavone [172], geraniol [153], GOFA/β-CD [173], menthol [174], organomagnesium [175], oxykine [176], PEITC [171], PSK [177], silibinin [178], tumerone [150], vitamin E [179], 3,3-diindolylmethane [180] | Natural compound | |
| Bismuth subnitrate [165], 3-aroylmethylene-2,3,6,7-tetrahydro-1H-pyrazino[2,1-a]isoquinolin-4(11bH)-ones [181] | Low-molecular weight compound | |
| Melatonin [160], N-acetylcysteine [182], selenium [182] | Amino acid and its derivative | |
| III. Suppression of iNOS | EVOO [183], FBRA [184], MEFA [147], MELA [147], oligonol [166], PAG [148] | Food product |
| Astaxanthin [185], baicalein [186], betaine [187], canolol [151], crocin [170], curcumin [188], inotilone [154], nobiletin [156], organomagnesium [175], pterostilbene [189], silibinin [178], UDCA [190], 5-OH-HxMF [191] | Natural compound | |
| Aminoguanidine [131], bezafibrate [192], GOFA-L-NAME [193], omeprazole [194], ONO-1714 [195], troglitazone [192] | Low-molecular weight compound | |
| Aspirin [196], nimesulide [192] | COX inhibitor | |
| Glutamine [197] | Amino acid and its derivative | |
| IV. Induction of antioxidant enzymes | Juzen-taiho-to [165], oligonol [166], protandim [167] | Food product |
| Crocin [170], DBM [171], digitoflavone [172], geraniol [153], GOFA/β-CD [173], menthol [174], organomagnesium [175], PEITC [171], PSK [177], 3,3-diindolylmethane [180] | Natural compound | |
| Bismuth subnitrate [165], 3-aroylmethylene-2,3,6,7-tetrahydro-1H-pyrazino[2,1-a]isoquinolin-4(11bH)-ones [181] | Low-molecular weight compound | |
| Melatonin [160] | Amino acid and its derivative | |
| V. Antioxidants | Auraptene [150], benzyl isothiocyanate [168], caffeine [169], geraniol [153], oxykine [176], silibinin [178], tumerone [150], vitamin E [179] | Natural compound |
| N-acetylcysteine [182], selenium [182] | Amino acid and its derivative | |
| VI. Inactivation of NF-κB | Apple oligogalactan [144], EAPP [198], FBE [199], ME [199], oligonol [166], PAG [148], protandim [167] | Food product |
| Astaxanthin [185], baicalein [186], betaine [187], crocin [170], curcumin [188], genistein-27 [152], GOFA/β-CD [173], inotilone [154], menthol [174], micheliolide [155], pterostilbene [189], silibinin [178], tricin [200], vitamin D [157], 3,3-diindolylmethane [180], 5-OH-HxMF [191] | Natural compound | |
| Cerulenin [201] | Low-molecular weight compound | |
| Glutamine [197], melatonin [160] | Amino acid and its derivative | |
| MiR-214 chemical inhibitor [202] | Oligonucleotides | |
| VII. Downregulation of pro-inflammatory cytokines | Apple oligogalactan [144], EVOO [183], FBRA [145], Ganoderma lucidum [146], MEFA [147], MELA [147], oligonol [166] | Food product |
| Astaxanthin [185], betaine [187], canolol [151], crocin [170], curcumin [188], digitoflavone [172], genistein-27 [152], GOFA/β-CD [173], isoliquiritigenin [139], micheliolide [155], organomagnesium [175], oroxylin A [203], pterostilbene [189], silibinin [178], tricin [200], triptolide [204], resveratrol [205], UDCA [190], vitamin D [157] | Natural compound | |
| Cerulenin [201], GOFA-L-NAME [193], NT1014 [206], omeprazole [194], statin hydroxamate [159], 3-aroylmethylene-2,3,6,7-tetrahydro-1H-pyrazino[2,1-a]isoquinolin-4(11bH)-ones [181], 5-aza-dC [207] | Low-molecular weight compound | |
| Glutamine [197], melatonin [160] | Amino acid and its derivative | |
| Aspirin [196] | COX inhibitor | |
| Cholera-toxin [162], α-lactalbumin [208] | Protein | |
| Oligonucleotides [163] | Oligonucleotides | |
| Eicosapentaenoic acid-free fatty acid [209] | Fatty acid | |
| VIII. Upregulation of anti-inflammatory cytokines | PSK [177] | Natural compound |
| Cholera-toxin [162] | Protein | |
| IX. Downregulation of chemokines | FBRA [145] | Food product |
| Auraptene [150], tumerone [150], vitamin D [157] | Natural compound | |
| Statin hydroxamate [159] | Low-molecular weight compound | |
| Glutamine [197] | Amino acid and its derivative | |
| Oligonucleotides [163] | Oligonucleotides | |
| X. Inhibition of COX-2 | EVOO [183], FBRA [184], Ganoderma lucidum [146], MEFA [147], MELA [147], oligonol [166], PAG [148], γ-TmT [149] | Food product |
| Astaxanthin [185], betaine [187], canolol [151], crocin [170], curcumin [188], geraniol [153], inotilone [154], isoliquiritigenin [139], menthol [174], nobiletin [156], organomagnesium [175], pterostilbene [189], resveratrol [205], silibinin [178], 3,3-diindolylmethane [180], 5-OH-HxMF [191] | Natural compound | |
| Bezafibrate [192], cerulenin [201], GOFA-L-NAME [193], omeprazole [194], statin hydroxamate [159], troglitazone [192] | Low-molecular weight compound | |
| Glutamine [197], melatonin [160] | Amino acid and its derivative | |
| Aspirin [196], celecoxib [210], MF-tricyclic [211], nimesulide [192], sulindac [161] | COX inhibitor | |
| α-lactalbumin [208] | Protein | |
| Oligonucleotides [163] | Oligonucleotides | |
| Eicosapentaenoic acid-free fatty acid [209] | Fatty acid |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanda, Y.; Osaki, M.; Okada, F. Chemopreventive Strategies for Inflammation-Related Carcinogenesis: Current Status and Future Direction. Int. J. Mol. Sci. 2017, 18, 867. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040867
Kanda Y, Osaki M, Okada F. Chemopreventive Strategies for Inflammation-Related Carcinogenesis: Current Status and Future Direction. International Journal of Molecular Sciences. 2017; 18(4):867. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040867
Chicago/Turabian StyleKanda, Yusuke, Mitsuhiko Osaki, and Futoshi Okada. 2017. "Chemopreventive Strategies for Inflammation-Related Carcinogenesis: Current Status and Future Direction" International Journal of Molecular Sciences 18, no. 4: 867. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040867