
Aneurysm-Specific miR-221 and miR-146a Participates in Human Thoracic and Abdominal Aortic Aneurysms

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Patient Demographics
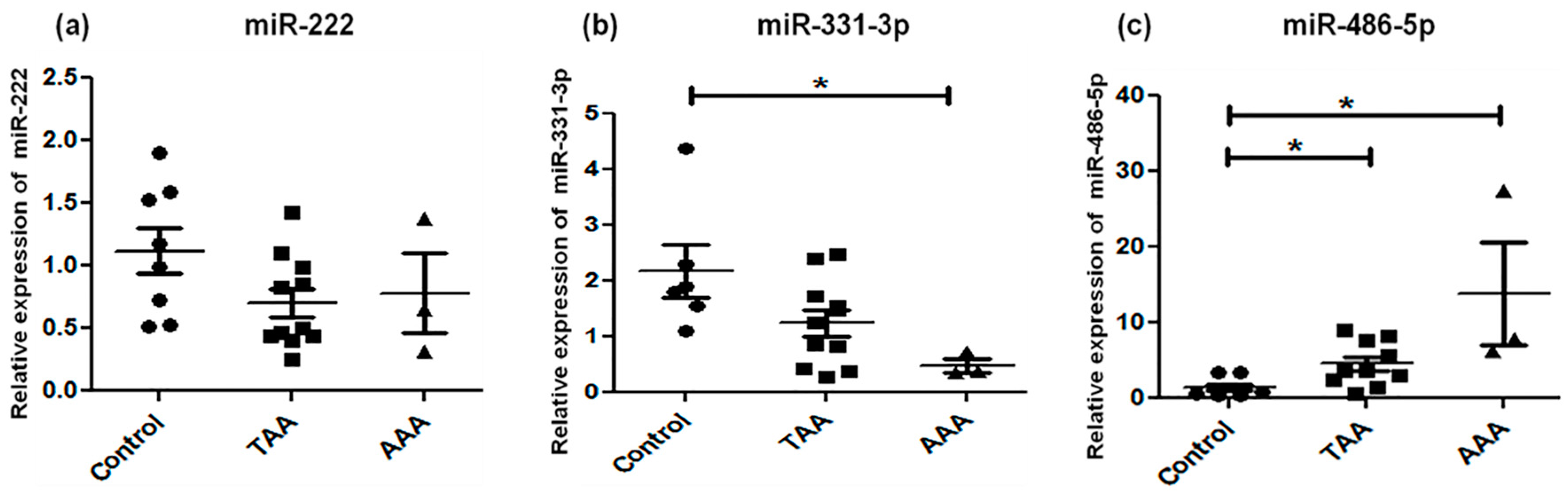
2.2. Analysis of Candidate miRNA Expression
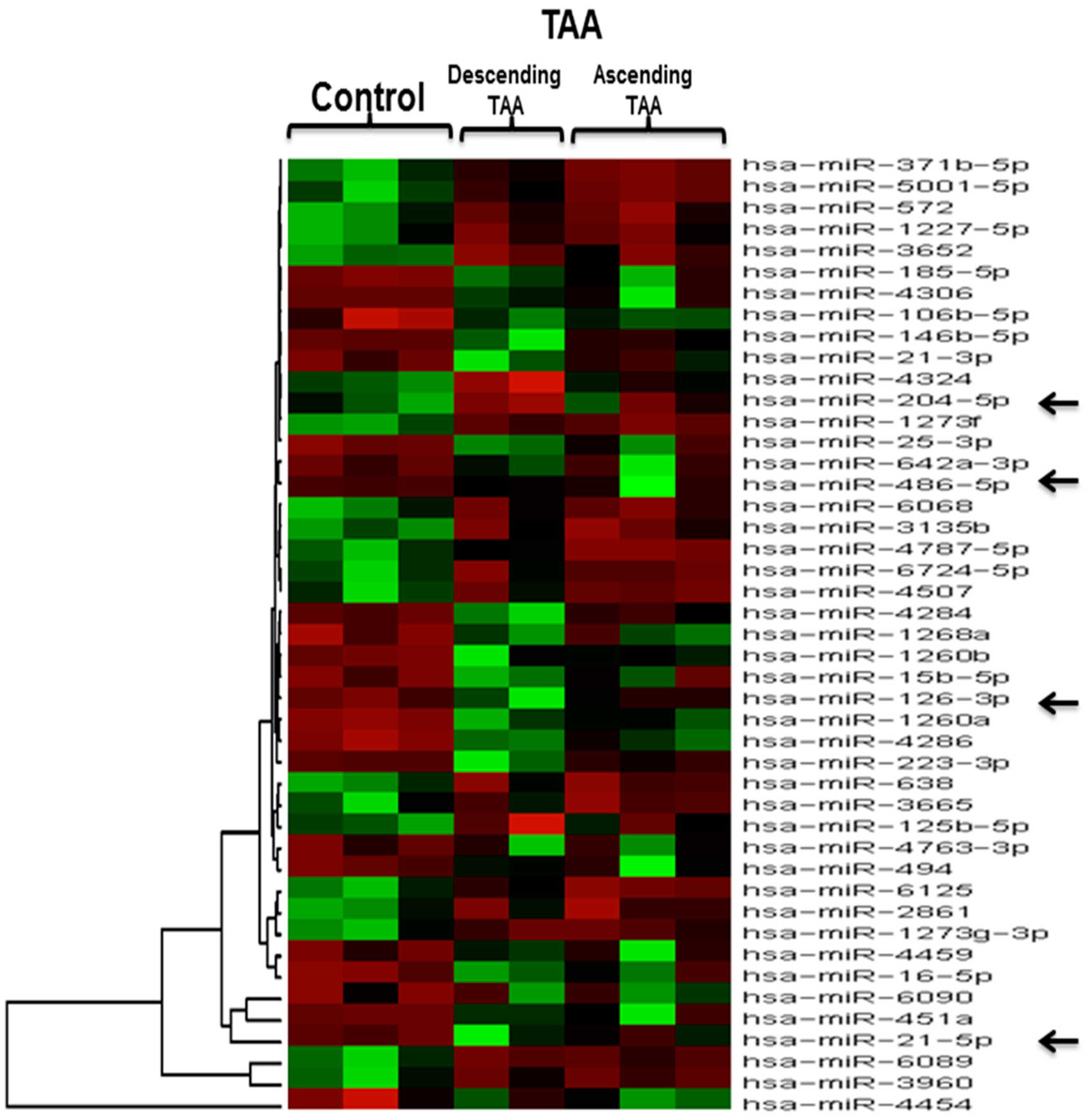
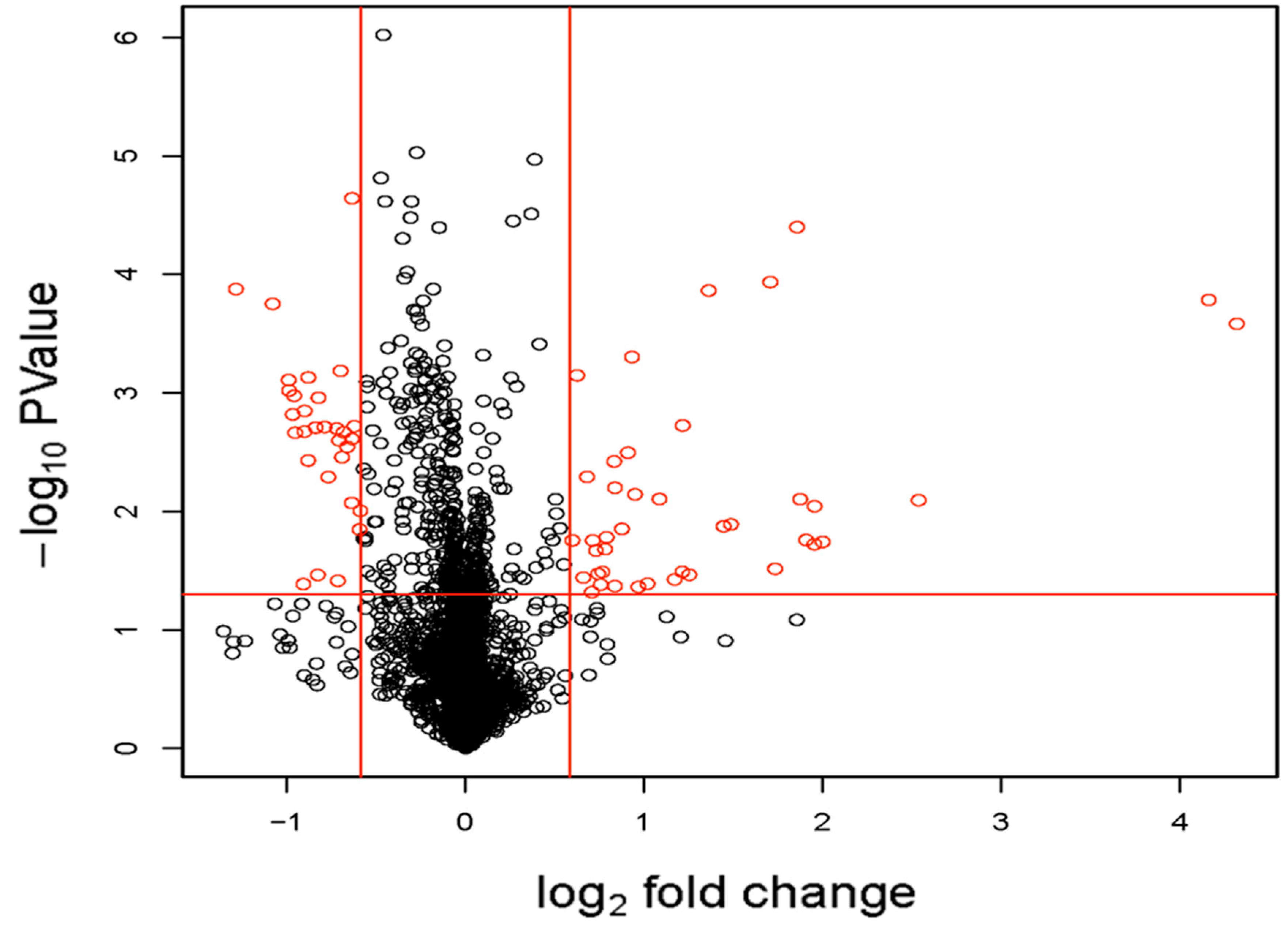
2.3. Microarray Expression Analysis
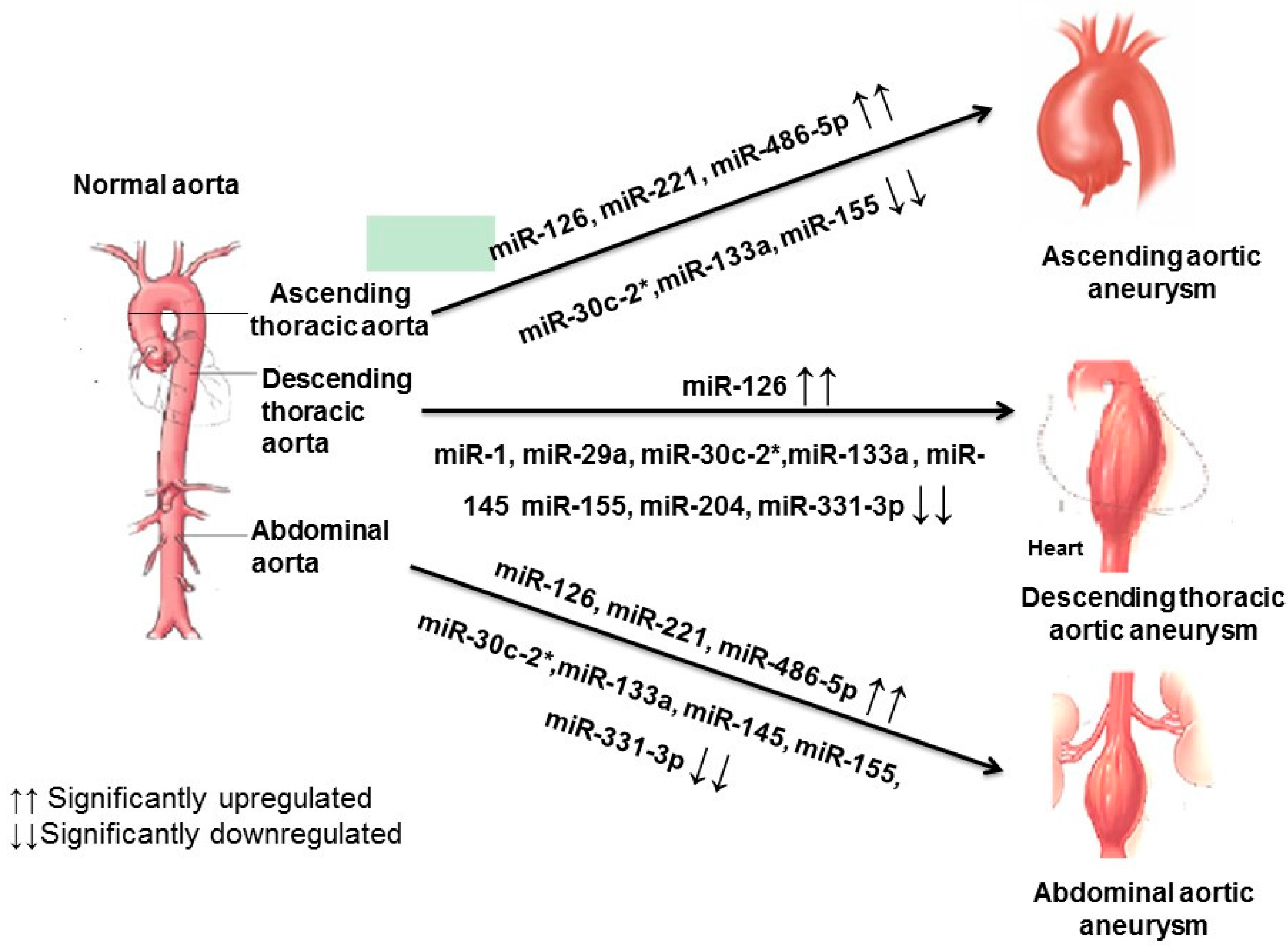
3. Discussion
4. Materials and Methods
4.1. Patient Demographics
4.2. Sample Preparation and miRNA Analysis
4.3. Microarray and Bioinformatics Analysis
4.4. Statistical Analysis
4.5. Study Limitations
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meszaros, I.; Morocz, J.; Szlavi, J.; Schmidt, J.; Tornoci, L.; Nagy, L.; Szep, L. Epidemiology and clinicopathology of aortic dissection. Chest 2000, 117, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Bozeman, M.C.; Ross, C.B. Intra-abdominal hypertension and abdominal compartment syndrome in association with ruptured abdominal aortic aneurysm in the endovascular era: Vigilance remains critical. Crit. Care Res. Pract. 2012, 2012, 151650. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Bailey, K.R.; Austin, E.; Kullo, I.J. Family history of atherosclerotic vascular disease is associated with the presence of abdominal aortic aneurysm. Vasc. Med. 2016, 21, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Peshkova, I.O.; Schaefer, G.; Koltsova, E.K. Atherosclerosis and aortic aneurysm: Is inflammation a common denominator? FEBS J. 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Abdulkareem, N.; Smelt, J.; Jahangiri, M. Bicuspid aortic valve aortopathy: Genetics, pathophysiology and medical therapy. Interact. Cardiovasc. Thorac. Surg. 2013, 17, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Cury, M.; Zeidan, F.; Lobato, A.C. Aortic disease in the young: Genetic aneurysm syndromes, connective tissue disorders, and familial aortic aneurysms and dissections. Int. J. Vasc. Med. 2013, 2013, 267215. [Google Scholar] [CrossRef] [PubMed]
- Elefteriades, J.A. Natural history of thoracic aortic aneurysms: Indications for surgery, and surgical versus nonsurgical risks. Ann. Thorac. Surg. 2002, 74, S1877–S1880. [Google Scholar] [CrossRef]
- Seeger, T.; Boon, R.A. MicroRNAs in cardiovascular ageing. J. Physiol. 2016, 594, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Murchison, E.P.; Tang, R.; Callis, T.E.; Tatsuguchi, M.; Deng, Z.; Rojas, M.; Hammond, S.M.; Schneider, M.D.; Selzman, C.H.; et al. Targeted deletion of Dicer in the heart leads to dilated cardiomyopathy and heart failure. Proc. Natl. Acad. Sci. USA 2008, 105, 2111–2116. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Tabin, C.J. Mirna-processing enzyme dicer is necessary for cardiac outflow tract alignment and chamber septation. Proc. Natl. Acad. Sci. USA 2010, 107, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Samal, E.; Srivastava, D. Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature 2005, 436, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Cordes, K.R.; Sheehy, N.T.; White, M.P.; Berry, E.C.; Morton, S.U.; Muth, A.N.; Lee, T.H.; Miano, J.M.; Ivey, K.N.; Srivastava, D. miR-145 and miR-143 regulate smooth muscle cell fate and plasticity. Nature 2009, 460, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Albinsson, S.; Suarez, Y.; Skoura, A.; Offermanns, S.; Miano, J.M.; Sessa, W.C. MicroRNAs are necessary for vascular smooth muscle growth, differentiation, and function. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Small, E.M.; Olson, E.N. Pervasive roles of microRNAs in cardiovascular biology. Nature 2011, 469, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Boon, R.A.; Seeger, T.; Heydt, S.; Fischer, A.; Hergenreider, E.; Horrevoets, A.J.; Vinciguerra, M.; Rosenthal, N.; Sciacca, S.; Pilato, M. MicroRNA-29 in aortic dilation: Implications for aneurysm formation. Circ. Res. 2011, 109, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Kin, K.; Miyagawa, S.; Fukushima, S.; Shirakawa, Y.; Torikai, K.; Shimamura, K.; Daimon, T.; Kawahara, Y.; Kuratani, T.; Sawa, Y. Tissue- and plasma-specific microRNA signatures for atherosclerotic abdominal aortic aneurysm. J. Am. Heart Assoc. 2012, 1, e000745. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Huang, A.; Ferruzzi, J.; Mecham, R.P.; Starcher, B.C.; Tellides, G.; Humphrey, J.D.; Giordano, F.J.; Niklason, L.E.; Sessa, W.C. Inhibition of microRNA-29 enhances elastin levels in cells haploinsufficient for elastin and in bioengineered vessels—Brief report. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Stroud, R.E.; O’Quinn, E.C.; Black, L.E.; Barth, J.L.; Elefteriades, J.A.; Bavaria, J.E.; Gorman, J.H.; Gorman, R.C.; Spinale, F.G. Selective microRNA suppression in human thoracic aneurysms relationship of miR-29a to aortic size and proteolytic induction. Circ. Cardiovasc. Genet. 2011, 4, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Zou, S.; Weng, J.; Hou, L.; Yang, L.; Zhao, Z.; Bao, J.; Jing, Z. A microRNA profile comparison between thoracic aortic dissection and normal thoracic aorta indicates the potential role of microRNAs in contributing to thoracic aortic dissection pathogenesis. J. Vasc. Surg. 2011, 53, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Pahl, M.C.; Derr, K.; Gabel, G.; Hinterseher, I.; Elmore, J.R.; Schworer, C.M.; Peeler, T.C.; Franklin, D.P.; Gray, J.L.; Carey, D.J.; et al. MicroRNA expression signature in human abdominal aortic aneurysms. BMC Med. Genom. 2012, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cheng, Y.; Zhang, S.; Lin, Y.; Yang, J.; Zhang, C. A necessary role of miR-221 and miR-222 in vascular smooth muscle cell proliferation and neointimal hyperplasia. Circ. Res. 2009, 104, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Merk, D.R.; Chin, J.T.; Dake, B.A.; Maegdefessel, L.; Miller, M.O.; Kimura, N.; Tsao, P.S.; Iosef, C.; Berry, G.J.; Mohr, F.W.; et al. miR-29b participates in early aneurysm development in marfan syndrome. Circ. Res. 2012, 110, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Fan, W.D.; Fang, R.; Wu, G.F. Regulation of microRNA-155 in endothelial inflammation by targeting nuclear factor (NF)-κB p65. J. Cell. Biochem. 2014, 115, 1928–1936. [Google Scholar] [CrossRef] [PubMed]
- Duisters, R.F.; Tijsen, A.J.; Schroen, B.; Leenders, J.J.; Lentink, V.; van der Made, I.; Herias, V.; van Leeuwen, R.E.; Schellings, M.W.; Barenbrug, P. MiR-133 and miR-30 regulate connective tissue growth factor implications for a role of microRNAs in myocardial matrix remodeling. Circ. Res. 2009, 104, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed]
- Boettger, T.; Beetz, N.; Kostin, S.; Schneider, J.; Kruger, M.; Hein, L.; Braun, T. Acquisition of the contractile phenotype by murine arterial smooth muscle cells depends on the miR143/145 gene cluster. J. Clin. Investig. 2009, 119, 2634–2647. [Google Scholar] [CrossRef] [PubMed]
- Elia, L.; Quintavalle, M.; Zhang, J.; Contu, R.; Cossu, L.; Latronico, M.V.; Peterson, K.L.; Indolfi, C.; Catalucci, D.; Chen, J.; et al. The knockout of miR-143 and -145 alters smooth muscle cell maintenance and vascular homeostasis in mice: Correlates with human disease. Cell Death Differ. 2009, 16, 1590–1598. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Small, E.M.; Sutherland, L.B.; Qi, X.; McAnally, J.; Plato, C.F.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. MicroRNAs miR-143 and miR-145 modulate cytoskeletal dynamics and responsiveness of smooth muscle cells to injury. Genes Dev. 2009, 23, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Guo, L.; Ji, J.; Zhang, J.; Chen, X.; Cai, Q.; Li, J.; Gu, Q.; Liu, B.; Zhu, Z.; et al. MiRNA-331-3p directly targets E2F1 and induces growth arrest in human gastric cancer. Biochem. Biophys. Res. Commun. 2010, 398, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Maegdefessel, L.; Azuma, J.; Toh, R.; Deng, A.; Merk, D.R.; Raiesdana, A.; Leeper, N.J.; Raaz, U.; Schoelmerich, A.M.; McConnell, M.V.; et al. MicroRNA-21 blocks abdominal aortic aneurysm development and nicotine-augmented expansion. Sci. Transl. Med. 2012, 4, 122ra122. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as biomarkers and diagnostics. J. Cell. Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.K.; Zhu, J.Q.; Zhang, J.T.; Li, Q.; Li, Y.; He, J.; Qin, Y.W.; Jing, Q. Circulating microRNA: A novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur. Heart J. 2010, 31, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Zhang, R.; Li, Y.; Pu, J.; Lu, Y.; Jiao, J.; Li, K.; Yu, B.; Li, Z.; Wang, R.; et al. Circulating microRNA-1 as a potential novel biomarker for acute myocardial infarction. Biochem. Biophys. Res. Commun. 2010, 391, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Albinsson, S.; Della Corte, A.; Alajbegovic, A.; Krawczyk, K.K.; Bancone, C.; Galderisi, U.; Cipollaro, M.; de Feo, M.; Forte, A. Patients with bicuspid and tricuspid aortic valve exhibit distinct regional microRNA signatures in mildly dilated ascending aorta. Heart Vessels 2017. [Google Scholar] [CrossRef] [PubMed]
- Biros, E.; Moran, C.S.; Rush, C.M.; Gabel, G.; Schreurs, C.; Lindeman, J.H.; Walker, P.J.; Nataatmadja, M.; West, M.; Holdt, L.M.; et al. Differential gene expression in the proximal neck of human abdominal aortic aneurysm. Atherosclerosis 2014, 233, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kandic, I.; Kutryk, M.J. Dysregulation of angiogenesis-related microRNAs in endothelial progenitor cells from patients with coronary artery disease. Biochem. Biophys. Res. Commun. 2011, 405, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, P.; Sun, T.; Li, D.; Xu, X.; Rui, Y.; Li, C.; Chong, M.; Ibrahim, T.; Mercatali, L.; et al. miR-126 and miR-126* repress recruitment of mesenchymal stem cells and inflammatory monocytes to inhibit breast cancer metastasis. Nat. Cell. Biol. 2013, 15, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, L.; Wang, H.; Zhu, S.; Dong, C.; Webster, K.A.; Wei, J. Synergistic induction of miR-126 by hypoxia and hdac inhibitors in cardiac myocytes. Biochem. Biophys. Res. Commun. 2013, 430, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Satoh, M.; Minami, Y.; Tabuchi, T.; Itoh, T.; Nakamura, M. Expression of miR-146a/b is associated with the toll-like receptor 4 signal in coronary artery disease: Effect of renin-angiotensin system blockade and statins on miRNA-146a/b and toll-like receptor 4 levels. Clin. Sci. 2010, 119, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Phillips, R.A.; Zhang, Y.; Khan, D.; Crasta, O.; Ahmed, S.A. Suppression of LPS-induced interferon-γ and nitric oxide in splenic lymphocytes by select estrogen-regulated microRNAs: A novel mechanism of immune modulation. Blood 2008, 112, 4591–4597. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cai, J.; Ma, F.; Lu, P.; Huang, H.; Zhou, J. miR-155 mediates suppressive effect of progesterone on TLR3, TLR4-triggered immune response. Immunol. Lett. 2012, 146, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Aalaei-Andabili, S.H.; Fabbri, M.; Rezaei, N. Reciprocal effects of toll-like receptors and miRNAs on biological processes in human health and disease: A systematic review. Immunotherapy 2013, 5, 1127–1142. [Google Scholar] [CrossRef] [PubMed]
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.; Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances microRNA miR-146a in ApoE−/− mice during experimental periodontal disease. Infect. Immun. 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- TEAM RC. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 18 March 2017).
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Site | Ascending/Descending | Surgical Diagnosis/Tissue Location | Hypertension/Smoking | |||
|---|---|---|---|---|---|---|
| Abdominal | ||||||
| 1 | AAA | AAA, for repair, aneurysm tissue, vessel wall | Y/Y | |||
| 2 | AAA | juxtarenal ascending aortic aneurysm, aortic wall | Y/Y | |||
| 3 | AAA | AAA, for repair | Y/Y | |||
| Thoracic | ||||||
| 1 | TAA | ascending | aortic arch repair, aorta, aneurysm wall | UK/N | ||
| 2 | TAA | ascending | ascending arch aneurysm, aorta | Y/Y | ||
| 3 | TAA | ascending | ascending/arch aneurysm, aorta | Y/Y | ||
| 4 | TAA | ascending | ascending aneurysm repair, aorta | N/Y | ||
| 5 | TAA | descending | thoracic aneurysm repair, aorta | N/N | ||
| 6 | TAA | ascending | ascending TAA, aorta | Y/N | ||
| 7 | TAA | descending | aortic aneurysm above a previous EVAR, aorta | Y/Y | ||
| 8 | DTAA | descending | descending TAA | |||
| 9 | TAA | ascending | ascending TAA | |||
| 10 | TAA | ascending | ascending TAA | |||
| 11 | TAA | ascending | TAA | |||
| Aorta | Operation | Aortic Diameter | Diagnosis | |||
| 1 | Control | Heart transplant recipient | <32 mm | Ischemic cardio myopathy | ||
| 2 | Control | Heart transplant recipient | <32 mm | Uhl’s anomaly | ||
| 3 | Control | Ascending aorta, total arch, descending aorta replacement | 34 mm | Kommerell’s diverticulum aneurysm | ||
| 4 | Control | Heart transplant recipient | <32 mm | Ischemic cardiomyopathy | ||
| 5 | Control | Redo root/asc/hemi arch replacement | 42 mm | Bicuspid aortic valve | ||
| 6 | Control | Heart transplant recipient | <32 mm | Ischemic cardiomyopathy | ||
| 7 | Control | Aortic valve replacement/root and ascending aorta replacement | 40 mm | Severe aortic insufficiency | ||
| 8 | Control | Heart transplant recipient | <32 mm | Congenital tricuspid atresia |
| miRNA | TAA | AAA | |
|---|---|---|---|
| 1 | miR-1 | Downregulated | No alteration |
| 2 | miR-21 | No alteration | No alteration |
| 3 | miR-29a | Downregulated | No alteration |
| 4 | miR-30c-2* | Downregulated | Downregulated |
| 5 | miR-124 | No alteration | No alteration |
| 6 | miR-126 | Upregulated | Upregulated |
| 7 | miR-133a | Downregulated | No alteration |
| 8 | miR-145 | No alteration | Downregulated |
| 9 | miR-146 | No alteration | Upregulated |
| 10 | miR-155 | Downregulated | Downregulated |
| 11 | miR-204 | Downregulated | Downregulated |
| 12 | miR-221 | Upregulated | No alteration |
| 13 | miR-222 | No alteration | No alteration |
| 14 | miR-331–3p | No alteration | Downregulated |
| 15 | miR-486–5p | Upregulated | Upregulated |
| Serial Number | miRNA | Fold Change | p Value |
|---|---|---|---|
| 1 | hsa-miR-15b-5p | 1.66 | 0.021 |
| 2 | hsa-miR-16-5p | 1.93 | 0.0072 |
| 3 | hsa-miR-21-3p | 1.58 | 0.036 |
| 4 | hsa-miR-21-5p | 3.67 | 0.0079 |
| 5 | hsa-miR-25-3p | 2.12 | 0.0079 |
| 6 | hsa-miR-106b-5p | 1.54 | 0.00071 |
| 7 | hsa-miR-126-3p | 3.88 | 0.0091 |
| 8 | hsa-miR-146b-5p | 2.72 | 0.013 |
| 9 | hsa-miR-185-5p | 1.88 | 0.0032 |
| 10 | hsa-miR-223-3p | 5.81 | 0.0081 |
| 11 | hsa-miR-451a | 17.91 | 0.00016 |
| 12 | hsa-miR-486-5p | 4.00 | 0.018 |
| 13 | hsa-miR-494 | 1.72 | 0.021 |
| 14 | hsa-miR-642a-3p | 2.32 | 0.032 |
| 15 | hsa-miR-1260b | 2.32 | 0.0019 |
| 16 | hsa-miR-1260a | 3.62 | 0.00004 |
| 17 | hsa-miR-1268a | 1.60 | 0.0051 |
| 18 | hsa-miR-4284 | 1.63 | 0.048 |
| 19 | hsa-miR-4286 | 1.91 | 0.00050 |
| 20 | hsa-miR-4306 | 1.73 | 0.017 |
| 21 | hsa-miR-4454 | 1.83 | 0.014 |
| 22 | hsa-miR-4459 | 1.69 | 0.42 |
| 23 | hsa-miR-4763-3p | 1.96 | 0.044 |
| 24 | hsa-miR-6090 | 1.79 | 0.043 |
| Serial Number | miRNA | Fold Change | p Value |
|---|---|---|---|
| 1 | hsa-miR-125b-5p | 0.41 | 0.041 |
| 2 | hsa-miR-204-5p | 0.47 | 0.035 |
| 3 | hsa-miR-371b-5p | 0.50 | 0.00078 |
| 4 | hsa-miR-572 | 0.50 | 0.0020 |
| 5 | hsa-miR-638 | 0.52 | 0.0035 |
| 6 | hsa-miR-1227-5p | 0.53 | 0.0025 |
| 7 | hsa-miR-1273g-3p | 0.54 | 0.00018 |
| 8 | hsa-miR-1273f | 0.54 | 0.00002 |
| 9 | hsa-miR-2861 | 0.56 | 0.0085 |
| 10 | hsa-miR-3135b | 0.56 | 0.0014 |
| 11 | hsa-miR-3652 | 0.59 | 0.00074 |
| 12 | hsa-miR-3665 | 0.61 | 0.014 |
| 13 | hsa-miR-3960 | 0.61 | 0.0019 |
| 14 | hsa-miR-4324 | 0.62 | 0.039 |
| 15 | hsa-miR-4507 | 0.62 | 0.0099 |
| 16 | hsa-miR-4787-5p | 0.63 | 0.0051 |
| 17 | hsa-miR-5001-5p | 0.64 | 0.00065 |
| 18 | hsa-miR-6068 | 0.64 | 0.0022 |
| 19 | hsa-miR-6089 | 0.65 | 0.00013 |
| 20 | hsa-miR-6125 | 0.66 | 0.00096 |
| 21 | hsa-miR-6724-5p | 0.66 | 0.0029 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venkatesh, P.; Phillippi, J.; Chukkapalli, S.; Rivera-Kweh, M.; Velsko, I.; Gleason, T.; VanRyzin, P.; Aalaei-Andabili, S.H.; Ghanta, R.K.; Beaver, T.; et al. Aneurysm-Specific miR-221 and miR-146a Participates in Human Thoracic and Abdominal Aortic Aneurysms. Int. J. Mol. Sci. 2017, 18, 875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040875
Venkatesh P, Phillippi J, Chukkapalli S, Rivera-Kweh M, Velsko I, Gleason T, VanRyzin P, Aalaei-Andabili SH, Ghanta RK, Beaver T, et al. Aneurysm-Specific miR-221 and miR-146a Participates in Human Thoracic and Abdominal Aortic Aneurysms. International Journal of Molecular Sciences. 2017; 18(4):875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040875
Chicago/Turabian StyleVenkatesh, Premakumari, Julie Phillippi, Sasanka Chukkapalli, Mercedes Rivera-Kweh, Irina Velsko, Thomas Gleason, Paul VanRyzin, Seyed Hossein Aalaei-Andabili, Ravi Kiran Ghanta, Thomas Beaver, and et al. 2017. "Aneurysm-Specific miR-221 and miR-146a Participates in Human Thoracic and Abdominal Aortic Aneurysms" International Journal of Molecular Sciences 18, no. 4: 875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18040875