
Inhibitory Effects of Dimethyllirioresinol, Epimagnolin A, Eudesmin, Fargesin, and Magnolin on Cytochrome P450 Enzyme Activities in Human Liver Microsomes



Abstract
:1. Introduction
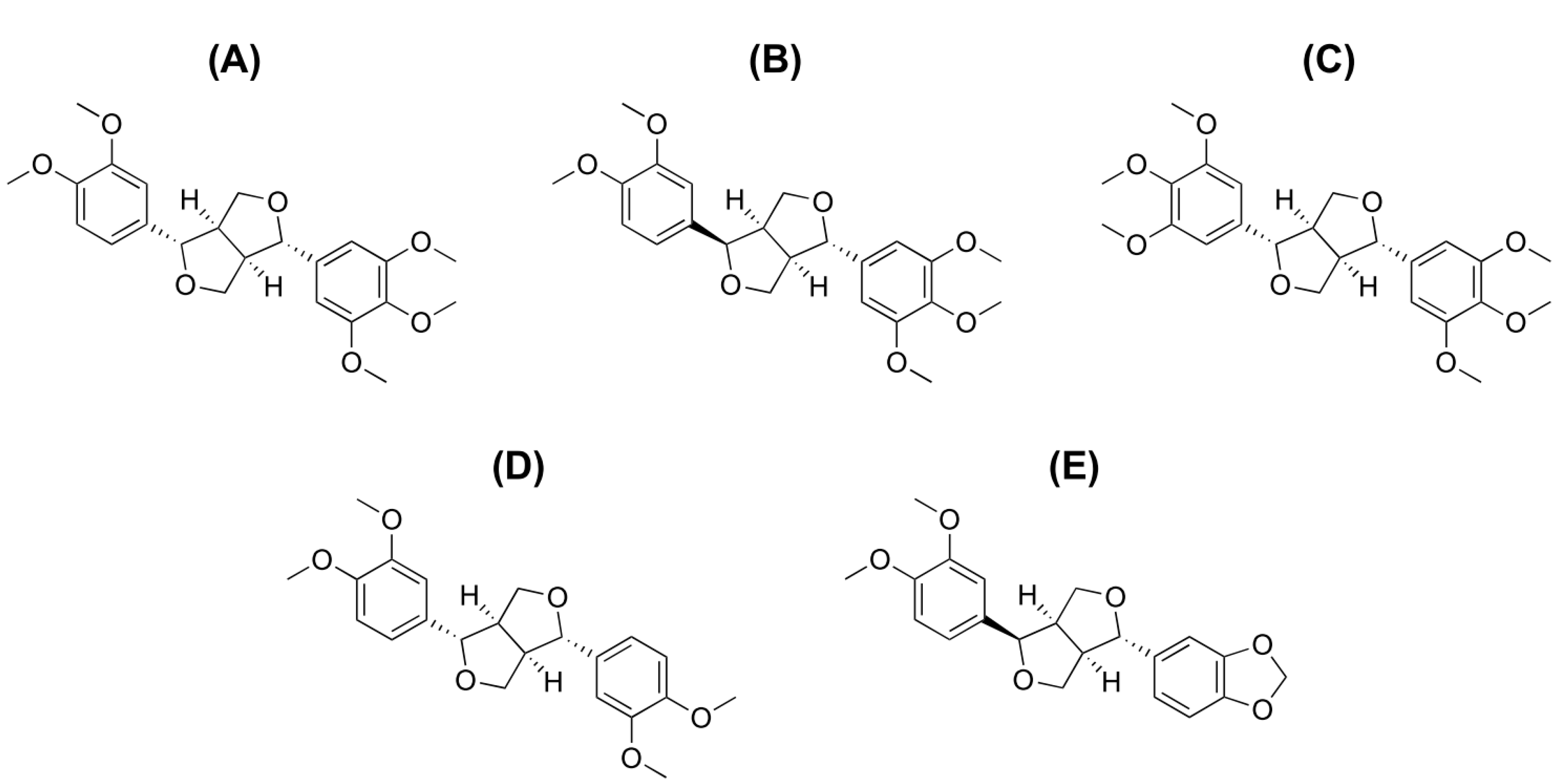
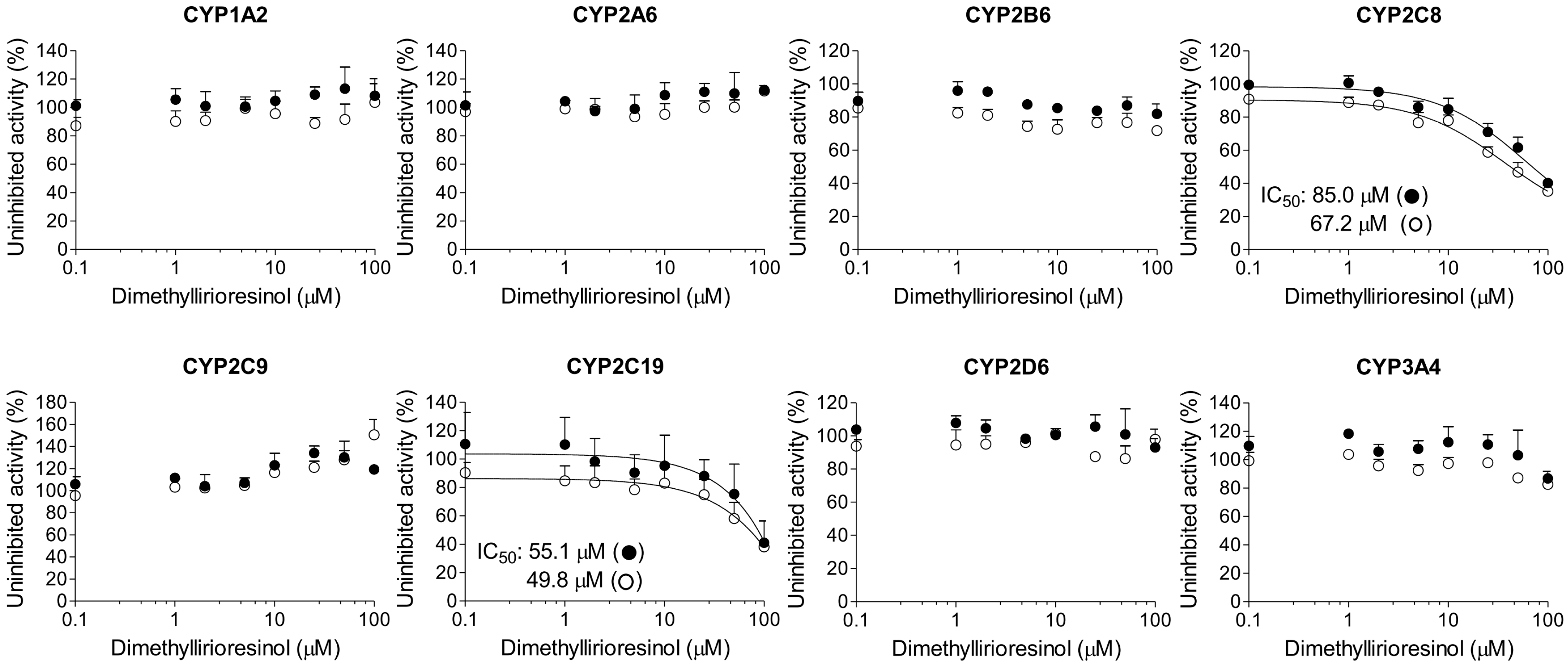
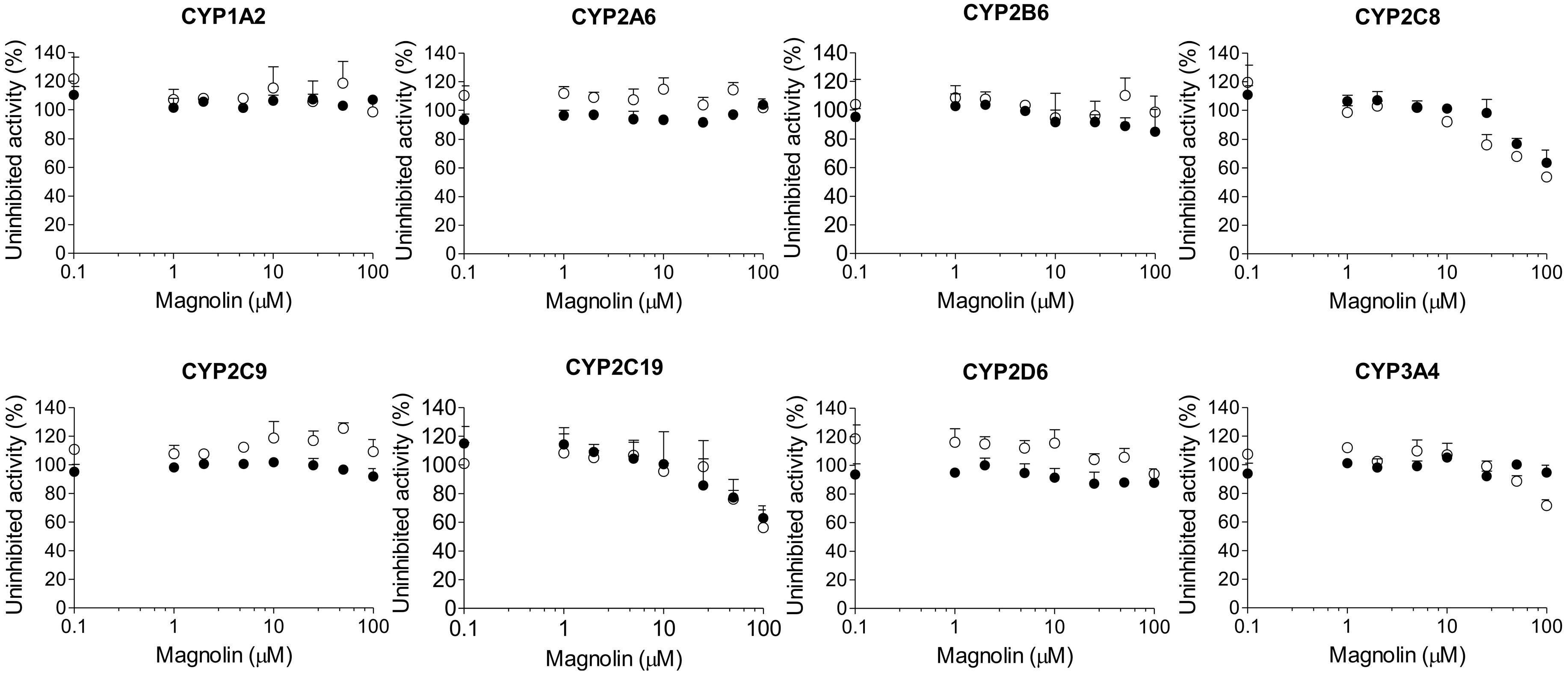
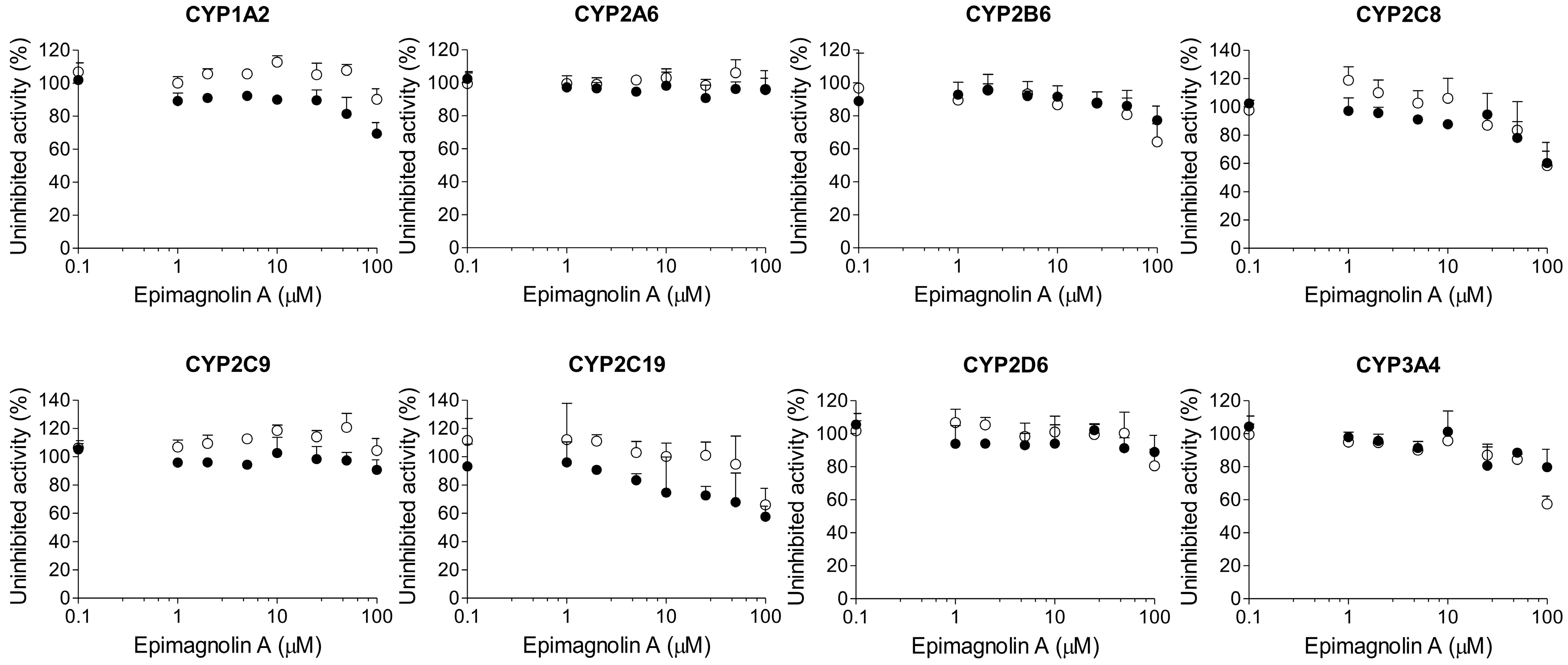
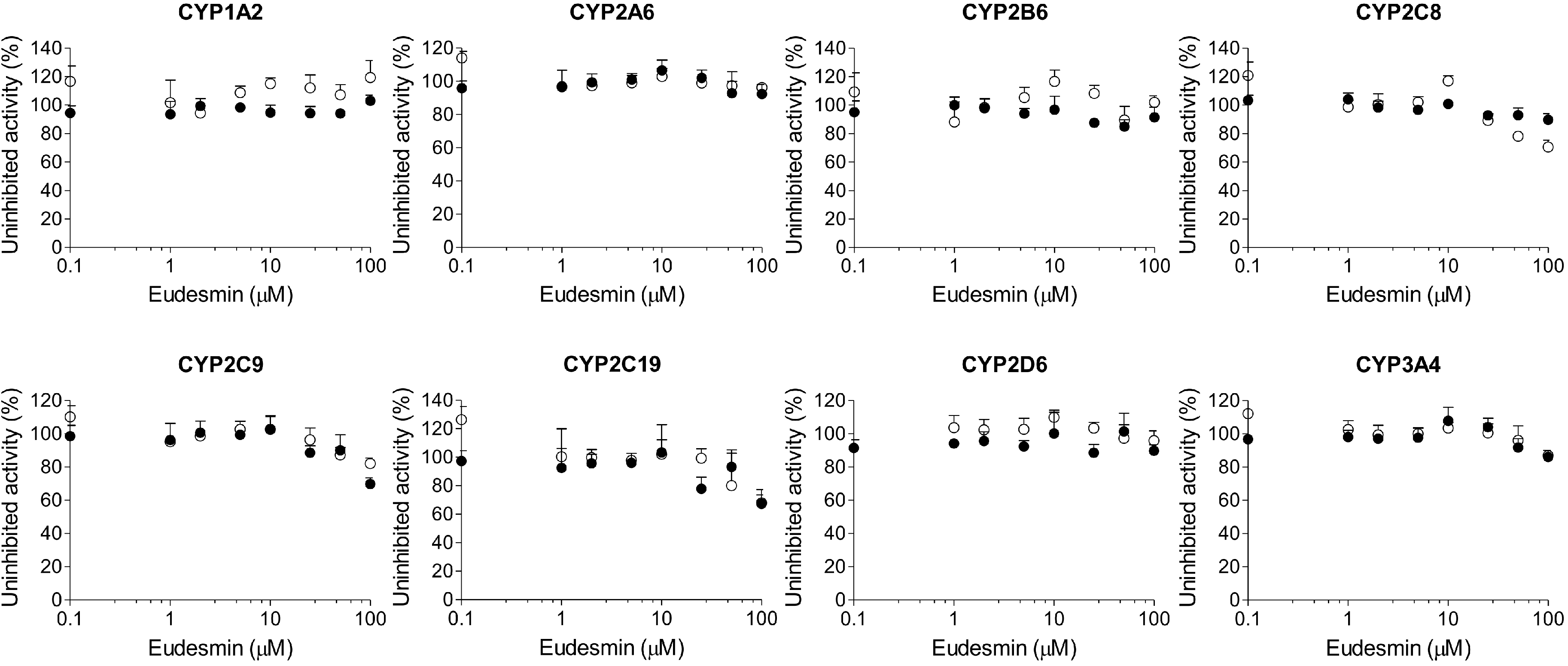
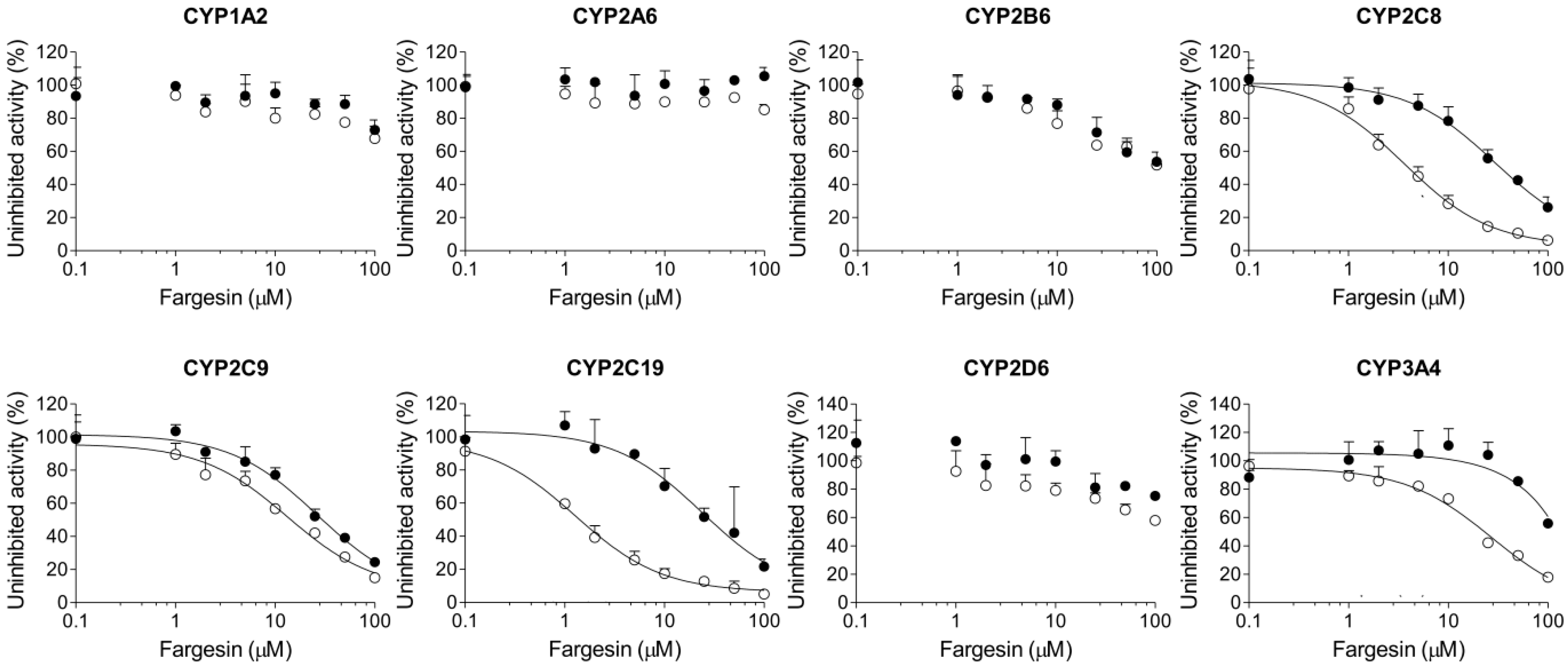
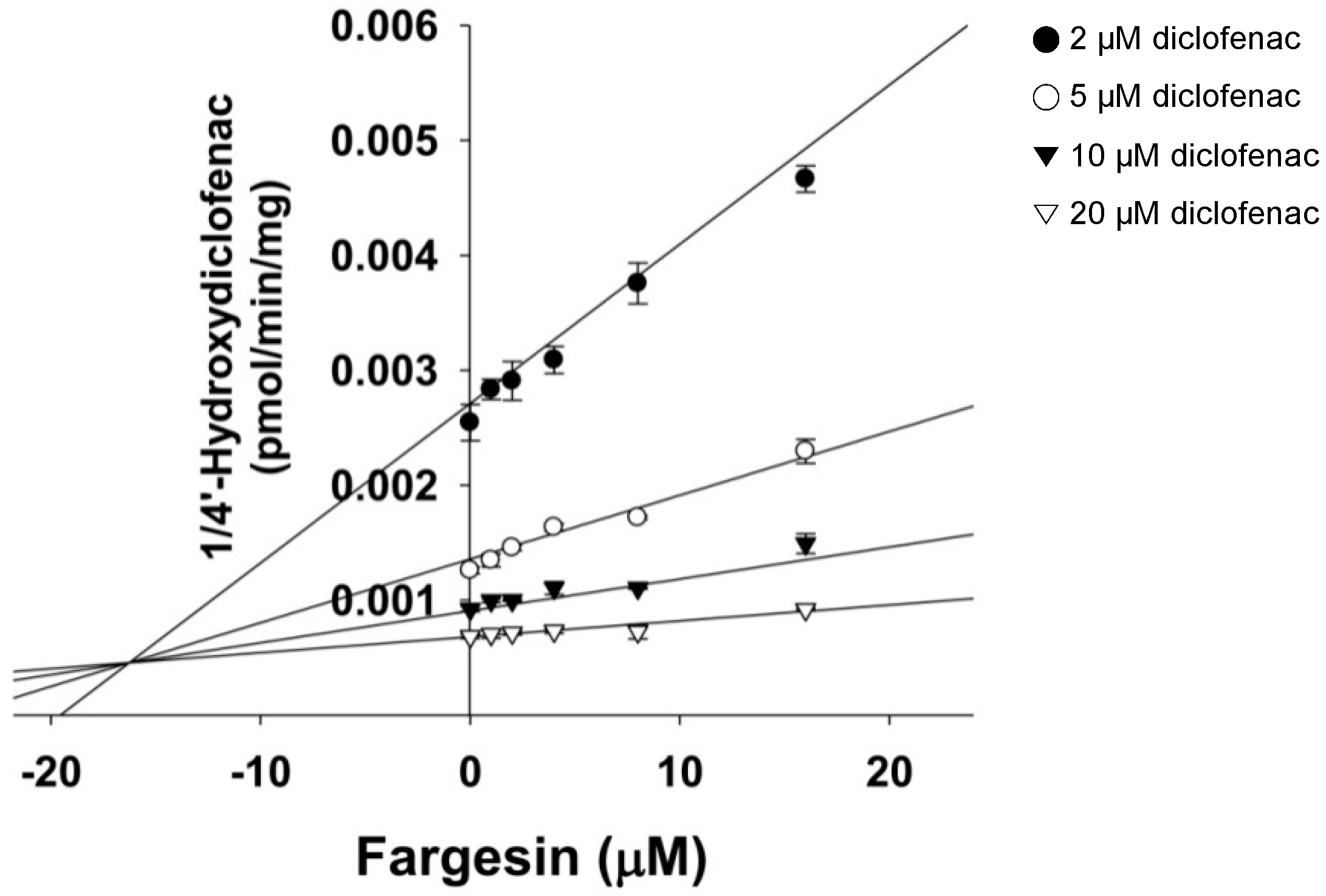
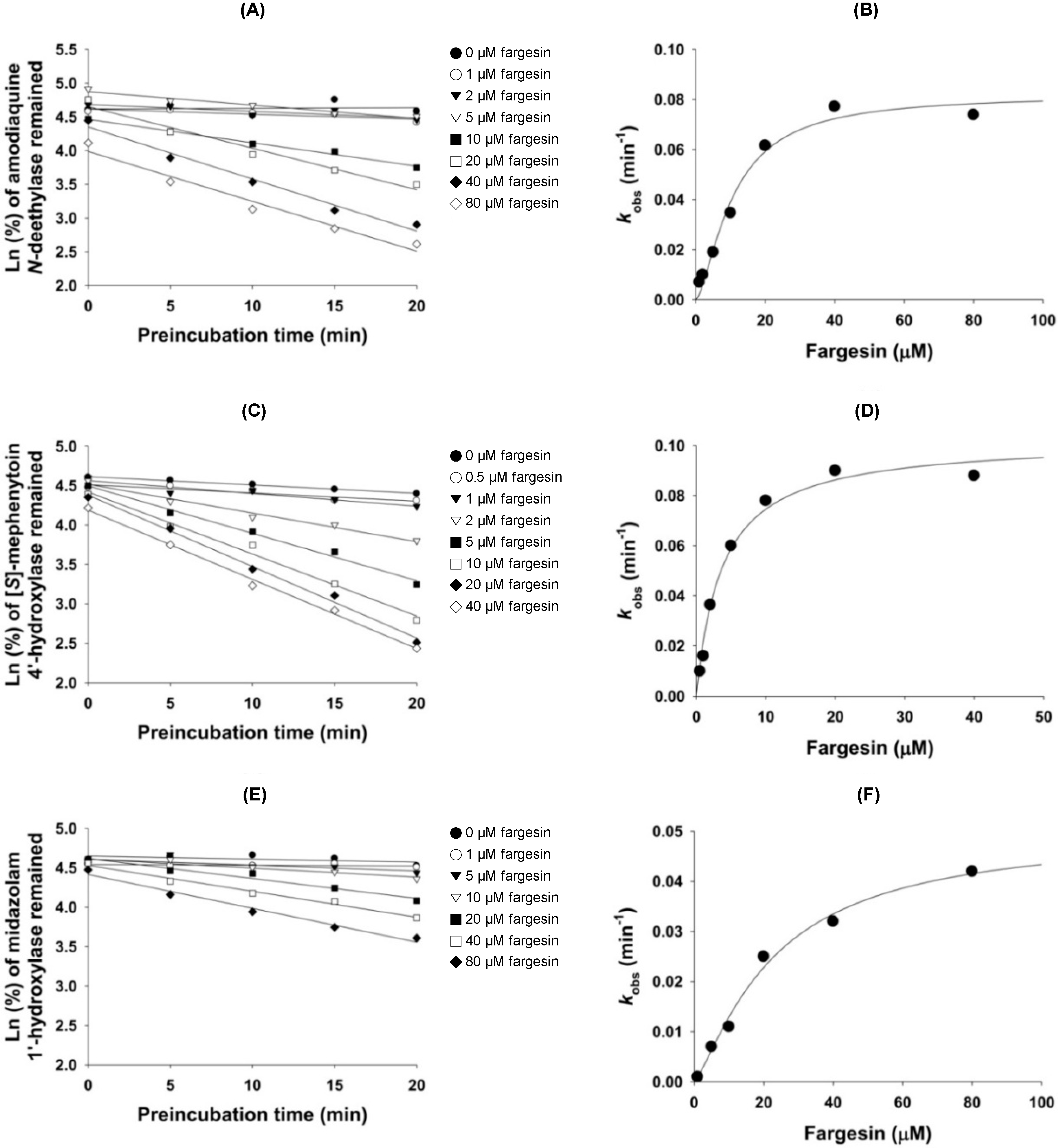
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Inhibitory Effects of Dimethyllirioresinol, Epimagnolin A, Eudesmin, Fargesin, and Magnolin on 8 Major CYP Activities in Human Liver Microsomes
4.3. Kinetic Analysis of CYP2C9 Inhibition by Fargesin
4.4. Mechanism-Based Inhibition of CYP2C8, CYP2C19, and CYP3A4 Activities by Fargesin
4.5. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhou, X.; Chen, C.; Ye, X.; Song, F.; Fan, G.; Wu, F. Analysis of lignans in Magnoliae flos by turbulent flow chromatography with online solid-phase extraction and high-performance liquid chromatography with tandem mass spectrometry. J. Sep. Sci. 2016, 39, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, V.; Sharma, S.; Kumar, N.; Sharma, U.; Singh, B. Simultaneous quantification and identification of flavonoids, lignans, coumarin and amides in leaves of Zanthoxylum armatum using UPLC-DAD-ESI-QTOF-MS/MS. J. Pharm. Biomed. Anal. 2017, 132, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar, S.; Singh, B.; Kumar, N. Quantitative and structural analysis of amides and lignans in Zanthoxylum armatum by UPLC-DAD-ESI-QTOF-MS/MS. J. Pharm. Biomed. Anal. 2014, 94, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Su, D.; Huang, Y.; Wang, Y.; Li, Y.-H. Ultrasound-assisted aqueous two-phase system for extraction and enrichment of Zanthoxylum armatum lignans. Molecules 2015, 20, 15273–15286. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Deng, Y.X.; Xie, H.; Yao, C.Y.; Cai, C.C.; Pan, S.L.; Wang, Y.L. Antinociceptive and anti-inflammatory activities of ethyl acetate fraction from Zanthoxylum armatum in mice. Fitoterapia 2011, 82, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Son, K.H.; Bae, K.H.; Hung, T.M.; Kim, Y.S.; Kim, H.P. 5-Lipoxygenase-inhibitory constituents from Schizandra fructus and Magnolia flos. Phytother. Res. 2009, 23, 1489–1492. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.A.; Lee, Y.D.; Lee, C.B.; Go, H.K.; Kim, J.P.; Seo, J.J.; Rhee, Y.K.; Kim, A.M.; Na, D.J. Extracts of Magnoliae flos inhibit inducible nitric oxide synthase via ERK in human respiratory epithelial cells. Nitric Oxide 2009, 20, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, J.Y.; Lee, H.J.; Lim, H.J.; Lee, D.Y.; Kim, D.H.; Ryu, J.H. Suppression of inducible nitric oxide synthase expression by furfuran lignans from flower buds of Magnolia fargesii in BV-2 microglial cells. Phytother. Res. 2010, 24, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, C.G.; Zhou, S.F.; Pang, E.C.; Story, D.F.; Xue, C.C. Chemistry and bioactivity of Flos Magnoliae, a Chinese herb for rhinitis and sinusitis. Curr. Med. Chem. 2008, 15, 1616–1627. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Kim, M.S.; Le, M.Q.; Song, Y.S.; Bak, Y.; Ryu, H.W.; Oh, S.R.; Yoon, D.Y. Fargesin exerts anti-inflammatory effects in THP-1 monocytes by suppressing PKC-dependent AP-1 and NF-κB signaling. Phytomedicine 2017, 24, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Arellanes, A.; Leon-Diaz, R.; Meckes, M.; Tapia, A.; Molina-Salinas, G.M.; Luna-Herrera, J.; Yepez-Mulia, L. Antiprotozoal and antimycobacterial activities of pure compounds from Aristolochia elegans rhizomes. Evid.-Based Complement. Altern. Med. 2012, 2012, 593403. [Google Scholar] [CrossRef] [PubMed]
- Jun, A.Y.; Kim, H.J.; Park, K.K.; Son, K.H.; Lee, D.H.; Woo, M.H.; Chung, W.Y. Tetrahydrofurofuran-type lignans inhibit breast cancer-mediated bone destruction by blocking the vicious cycle between cancer cells, osteoblasts and osteoclasts. Investig. New Drugs 2014, 32, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cheng, Y.; Xue, H.; Yue, Y.; Zhang, W.; Li, X. Fargesin as a potential β1 adrenergic receptor antagonist protects the hearts against ischemia/reperfusion injury in rats via attenuating oxidative stress and apoptosis. Fitoterapia 2015, 105, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.S.; Cha, B.Y.; Choi, B.K.; Lee, Y.S.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Fargesin, a component of Flos Magnoliae, stimulates glucose uptake in L6 myotubes. J. Nat. Med. 2013, 67, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Cha, B.Y.; Choi, S.S.; Harada, Y.; Choi, B.K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Fargesin improves lipid and glucose metabolism in 3T3-L1 adipocytes and high-fat diet-induced obese mice. Biofactors 2012, 38, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Sha, S.; Xu, D.; Wang, Y.; Zhao, W.; Li, X. Antihypertensive effects of fargesin in vitro and in vivo via attenuating oxidative stress and promoting nitric oxide release. Can. J. Physiol. Pharmacol. 2016, 94, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-J.; Lee, M.-H.; Yoo, S.-M.; Choi, K.-I.; Song, J.-H.; Jang, J.-H.; Oh, S.-R.; Ryu, H.-W.; Lee, H.-S.; Surh, Y.-J.; et al. Magnolin inhibits cell migration and invasion by targeting the ERKs/RSK2 signaling pathway. BMC Cancer 2015, 15, 576. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.J.; Lee, H.S.; Ryu, H.W.; Lee, M.H.; Lee, J.Y.; Li, Y.; Dong, Z.; Lee, H.K.; Oh, S.R.; Cho, Y.Y. Targeting of magnolin on ERKs inhibits Ras/ERKs/RSK2-signaling-mediated neoplastic cell transformation. Carcinogenesis 2014, 35, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zou, X.; Zhang, X.; Wang, F.; Zhu, W.; Zhang, G.; Xiao, J.; Chen, M. Magnolin inhibits prostate cancer cell growth in vitro and in vivo. Biomed. Pharmacother. 2017, 87, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, G.; Zhou, Y.; Gui, D.; Li, J.; Xing, T.; Wang, N. Magnolin protects against contrast-induced nephropathy in rats via antioxidation and antiapoptosis. Oxid. Med. Cell. Longev. 2014, 2014, 203458. [Google Scholar] [CrossRef] [PubMed]
- Na, D.H.; Ji, H.Y.; Park, E.J.; Kim, M.S.; Liu, K.H.; Lee, H.S. Evaluation of metabolism-mediated herb-drug interactions. Arch. Pharm. Res. 2011, 34, 1829–1842. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Z.; Zhang, Y.L.; Zeng, M.Z.; He, F.Z.; Luo, Z.Y.; Luo, J.Q.; Wen, J.G.; Chen, X.P.; Zhou, H.H.; Zhang, W. Pharmacogenomics and herb-drug interactions: Merge of future and tradition. Evid.-Based Complement. Altern. Med. 2015, 2015, 321091. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.L.; Ma, Y.M. Pharmacokinetic herb-drug interactions with traditional Chinese medicine: Progress, causes of conflicting results and suggestions for future research. Drug Metab. Rev. 2016, 48, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Liu, K. Pharmacokinetic interactions between herbal medicines and prescribed drugs: Focus on drug metabolic enzymes and transporters. Curr. Drug Metab. 2014, 15, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Brantley, S.J.; Argikar, A.A.; Lin, Y.S.; Nagar, S.; Paine, M.F. Herb–drug interactions: Challenges and opportunities for improved predictions. Drug Metab. Dispos. 2014, 42, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Roe, A.L.; Paine, M.F.; Gurley, B.J.; Brouwer, K.R.; Jordan, S.; Griffiths, J.C. Assessing natural product–drug interactions: An end-to-end safety framework. Regul. Toxicol. Pharmacol. 2016, 76, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Huang, M.; Kanfer, I.; Chow, M.S.; Cho, W.C. Herb-drug interactions: Systematic review, mechanisms, and therapies. Evid.-Based Complement. Altern. Med. 2015, 2015, 239150. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Scicchitano, F.; Whalley, B.J.; Mazzitello, C.; Ciriaco, M.; Esposito, S.; Patane, M.; Upton, R.; Pugliese, M.; Chimirri, S.; et al. Hypericum perforatum: Pharmacokinetic, mechanism of action, tolerability, and clinical drug-drug interactions. Phytother. Res. 2014, 28, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Unger, M. Pharmacokinetic drug interactions involving Ginkgo biloba. Drug Metab. Rev. 2013, 45, 353–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Chen, L.; Shan, L.; Fan, G.; Gao, X. Liquorice, a unique “guide drug” of traditional Chinese medicine: A review of its role in drug interactions. J. Ethnopharmacol. 2013, 150, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.U.; Lee, J.Y.; Kwon, S.S.; Kim, J.H.; Kim, Y.M.; Hong, S.W.; Yeon, S.H.; Lee, S.M.; Cho, Y.Y.; Lee, H.S. Metabolism-mediated drug interaction potential of HS-23, a new herbal drug for the treatment of sepsis in human hepatocytes and liver microsomes. Arch. Pharm. Res. 2015, 38, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.S.; Kim, J.H.; Jeong, H.U.; Cho, Y.Y.; Oh, S.R.; Lee, H.S. Inhibitory effects of aschantin on cytochrome P450 and uridine 5′-diphospho-glucuronosyltransferase enzyme activities in human liver microsomes. Molecules 2016, 21, E554. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.U.; Kong, T.Y.; Kwon, S.S.; Hong, S.W.; Yeon, S.H.; Choi, J.H.; Lee, J.Y.; Cho, Y.Y.; Lee, H.S. Effect of honokiol on cytochrome P450 and UDP-glucuronosyltransferase enzyme activities in human liver microsomes. Molecules 2013, 18, 10681–10693. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; You, J.; Choi, H.G.; Kim, J.A.; Jee, J.G.; Lee, S. Selective inhibitory effects of machilin A isolated from Machilus thunbergii on human cytochrome P450 1A and 2B6. Phytomedicine 2015, 22, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Taesotikul, T.; Dumrongsakulchai, W.; Wattanachai, N.; Navinpipat, V.; Somanabandhu, A.; Tassaneeyakul, W.; Tassaneeyakul, W. Inhibitory effects of Phyllanthus amarus and its major lignans on human microsomal cytochrome P450 activities: Evidence for CYP3A4 mechanism-based inhibition. Drug Metab. Pharmacokinet. 2011, 26, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Sun, D.X.; Chen, B.; Ji, D.H.; Pu, J.; Xu, J.; Tian, F.D.; Guo, L. Inhibition of CYP3A4 and CYP2C9 by podophyllotoxin: Implication for clinical drug-drug interactions. J. Biosci. 2011, 36, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.-H.; He, X.; Chen, J.-X.; He, L.-N.; Jin, C.-H.; Wang, L.-L.; Zhang, F.-L.; An, L.-J. Identification and characterization of reactive metabolites in myristicin-mediated mechanism-based inhibition of CYP1A2. Chem. Biol. Interact. 2015, 237, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, Y.; Jin, C.; Lee, S.H.; Kang, M.J.; Jeong, T.C.; Jeong, S.Y.; Kim, D.H.; Yoo, H.H. Inhibitory effects of deoxypodophyllotoxin from Anthriscus sylvestris on human CYP2C9 and CYP3A4. Planta Med. 2010, 76, 701–704. [Google Scholar] [CrossRef] [PubMed]
- Polasek, T.M.; Elliot, D.J.; Lewis, B.C.; Miners, J.O. Mechanism-based inactivation of human cytochrome P4502C8 by drugs in vitro. J. Pharmacol. Exp. Ther. 2004, 311, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Sakamoto, M.; Itoh, T.; Fujiwara, R. Underlying mechanism of drug–drug interaction between pioglitazone and gemfibrozil: Gemfibrozil acyl-glucuronide is a mechanism-based inhibitor of CYP2C8. Drug Metab. Pharmacokinet. 2015, 30, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Nishiya, Y.; Hagihara, K.; Kurihara, A.; Okudaira, N.; Farid, N.A.; Okazaki, O.; Ikeda, T. Comparison of mechanism-based inhibition of human cytochrome P450 2C19 by ticlopidine, clopidogrel, and prasugrel. Xenobiotica 2009, 39, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Tassaneeyakul, W.; Guo, L.Q.; Fukuda, K.; Ohta, T.; Yamazoe, Y. Inhibition selectivity of grapefruit juice components on human cytochromes P450. Arch. Biochem. Biophys. 2000, 378, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Tezuka, Y.; Kadota, S.; Hiratsuka, A.; Watabe, T. Mechanism-based inactivation of human liver microsomal CYP3A4 by rutaecarpine and limonin from Evodia fruit extract. Drug Metab. Pharmacokinet. 2005, 20, 34–45. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CYP | Enzyme Activities | IC50 (µM) | Ki (µM) | |
|---|---|---|---|---|
| No Pre-Incubation | With Pre-Incubation * | (kinact, min−1 or Inhibition Mode) | ||
| 1A2 | Phenacetin O-deethylase | >100 | >100 | - |
| 2A6 | Coumarin 7-hydroxylase | >100 | >100 | - |
| 2B6 | Bupropion hydroxylase | >100 | >100 | - |
| 2C8 | Amodiaquine N-deethylase | 34.9 | 4.0 | 10.7 (kinact: 0.082) |
| 2C9 | Diclofenac 4′-hydroxylase | 30.8 | 16.4 | 16.3 (competitive) |
| 2C19 | (S)-Mephenytoin 4′-hydroxylase | 30.2 | 1.6 | 3.7 (kinact: 0.102) |
| 2D6 | Bufuralol 1′-hydroxylase | >100 | >100 | - |
| 3A4 | Midazolam 1′-hydroxylase | >100 | 17.9 | 23.0 (kinact: 0.050) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Kwon, S.-S.; Jeong, H.-U.; Lee, H.S. Inhibitory Effects of Dimethyllirioresinol, Epimagnolin A, Eudesmin, Fargesin, and Magnolin on Cytochrome P450 Enzyme Activities in Human Liver Microsomes. Int. J. Mol. Sci. 2017, 18, 952. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18050952
Kim J-H, Kwon S-S, Jeong H-U, Lee HS. Inhibitory Effects of Dimethyllirioresinol, Epimagnolin A, Eudesmin, Fargesin, and Magnolin on Cytochrome P450 Enzyme Activities in Human Liver Microsomes. International Journal of Molecular Sciences. 2017; 18(5):952. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18050952
Chicago/Turabian StyleKim, Ju-Hyun, Soon-Sang Kwon, Hyeon-Uk Jeong, and Hye Suk Lee. 2017. "Inhibitory Effects of Dimethyllirioresinol, Epimagnolin A, Eudesmin, Fargesin, and Magnolin on Cytochrome P450 Enzyme Activities in Human Liver Microsomes" International Journal of Molecular Sciences 18, no. 5: 952. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18050952