
Comparative Proteomics of Rubber Latex Revealed Multiple Protein Species of REF/SRPP Family Respond Diversely to Ethylene Stimulation among Different Rubber Tree Clones

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
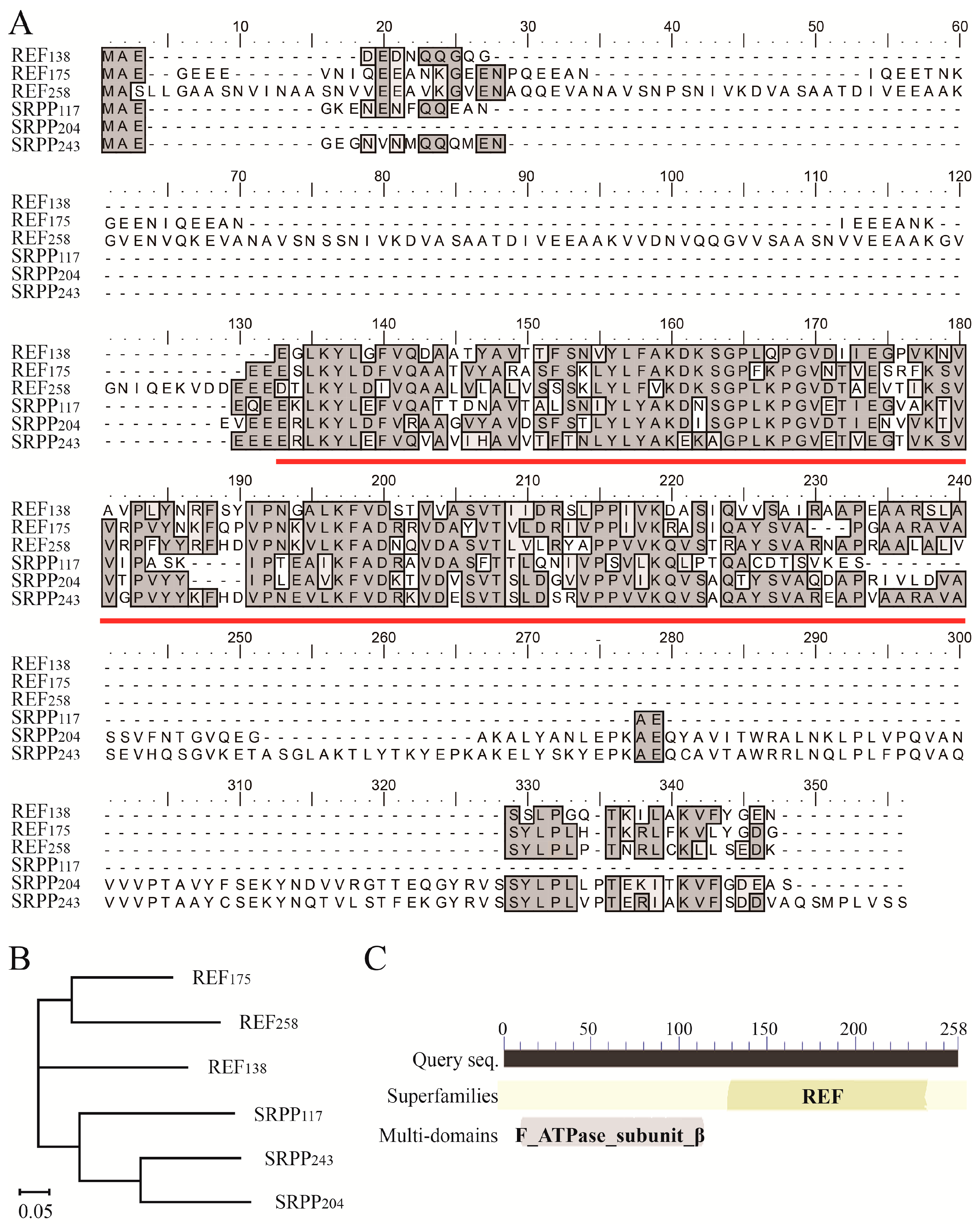
2.1. REF and SRPP Subfamily Members in H. brasiliensis
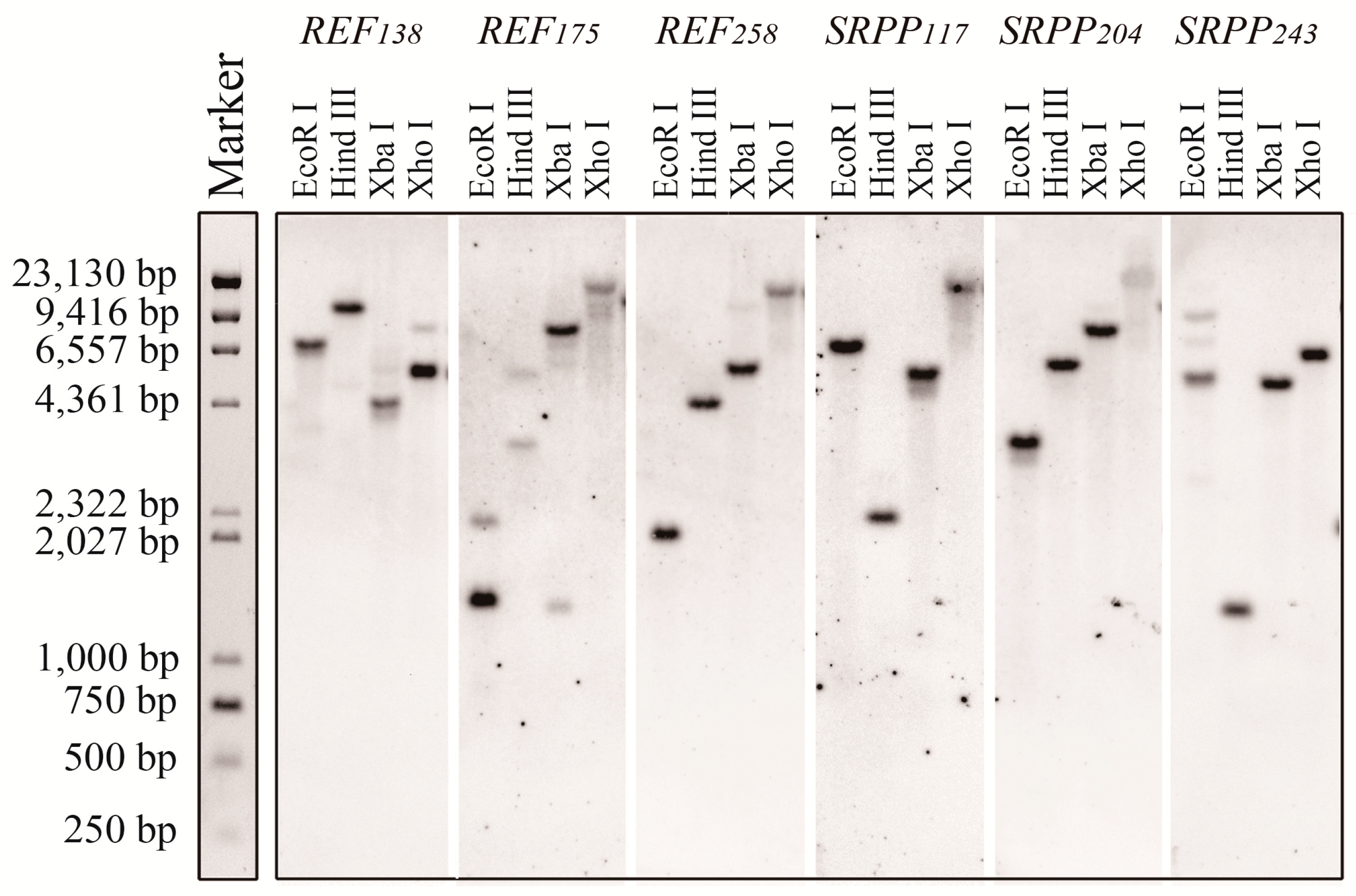
2.2. Detection of the Six REF and SRPP Genes in H. brasiliensis Genomic DNA
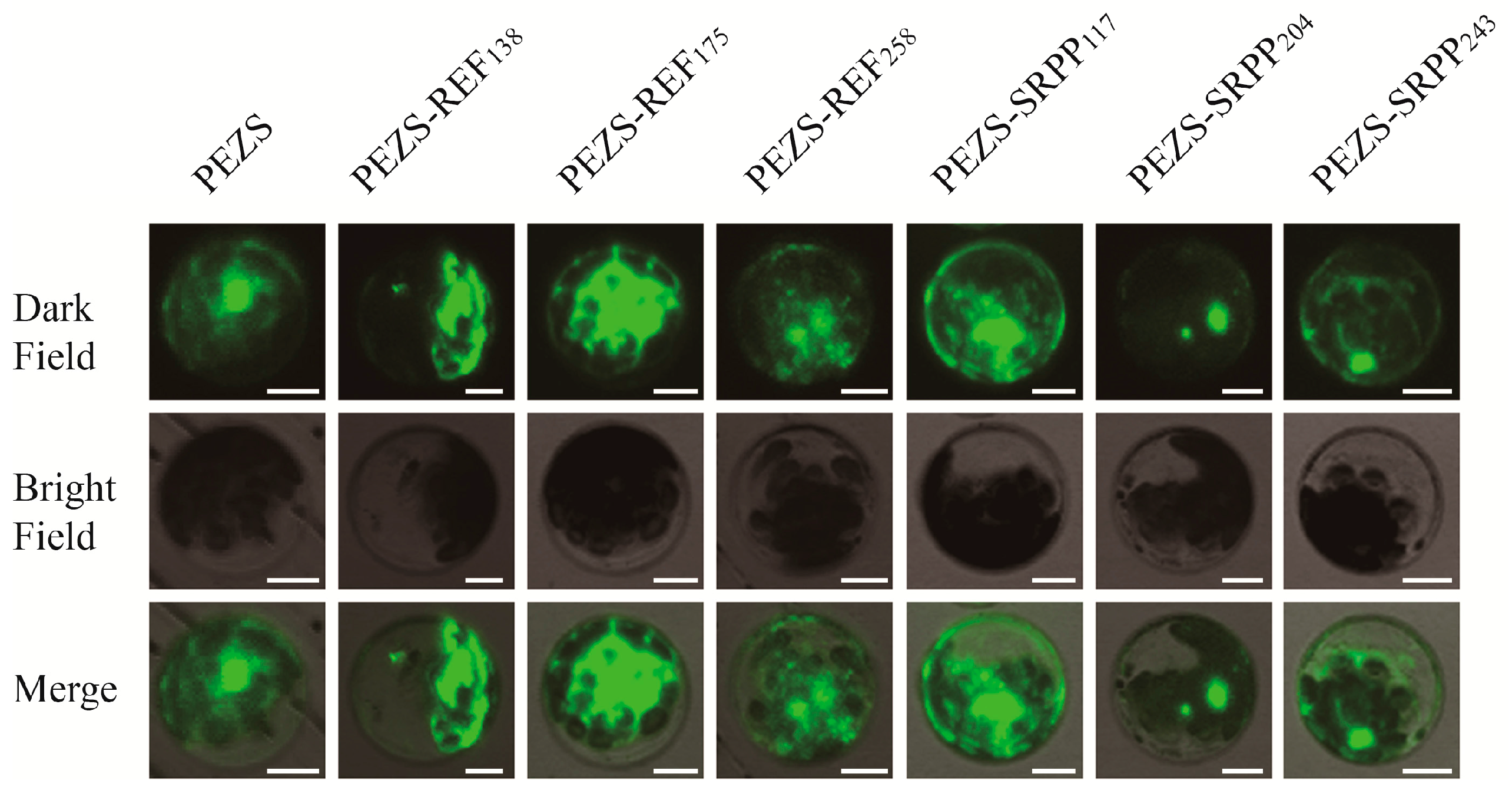
2.3. Subcellular Localization of REF and SRPP in Arabidopsis Protoplasts
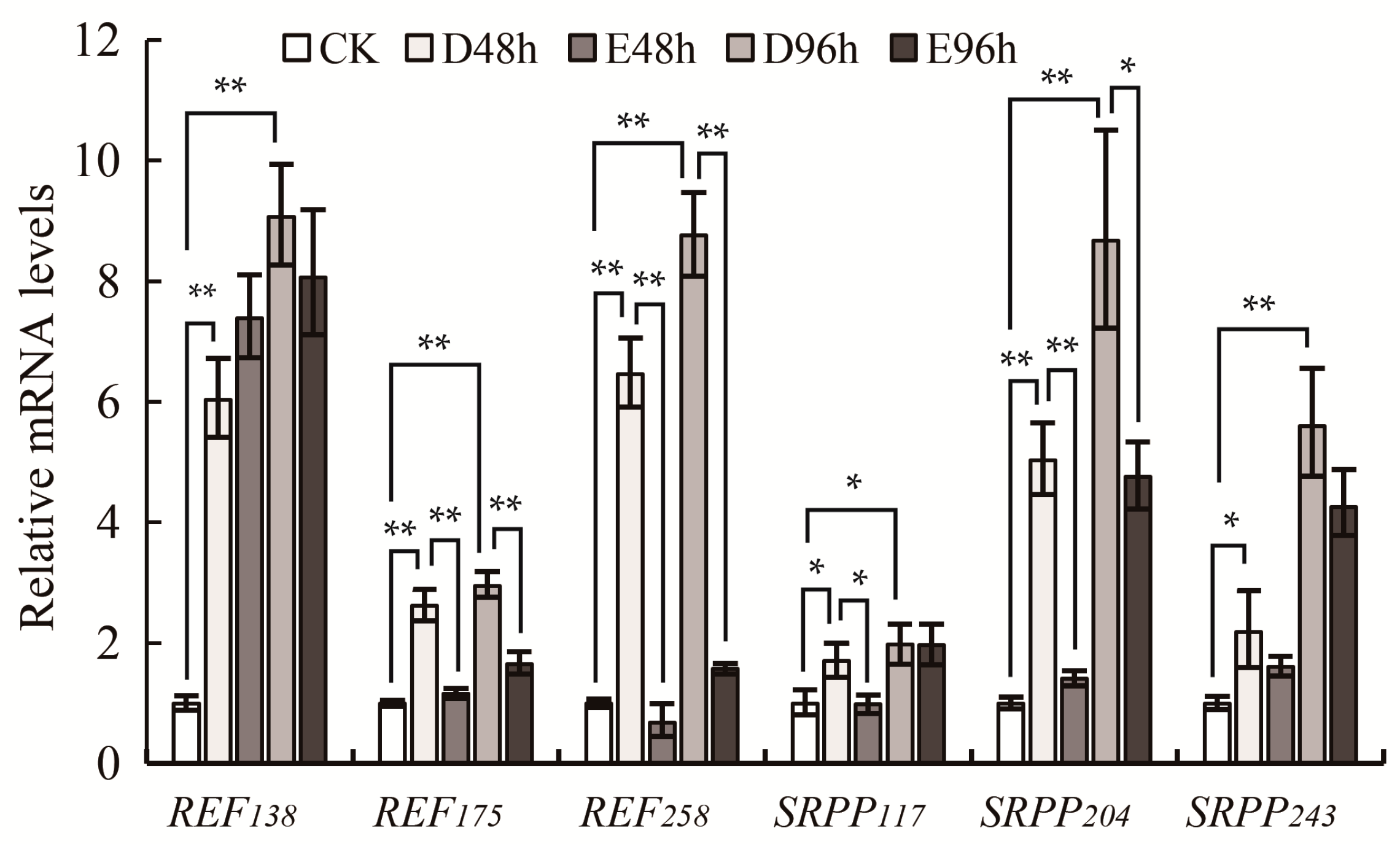
2.4. Gene Expression Patterns of REF and SRPP in Rubber Latex under Ethylene Treatment
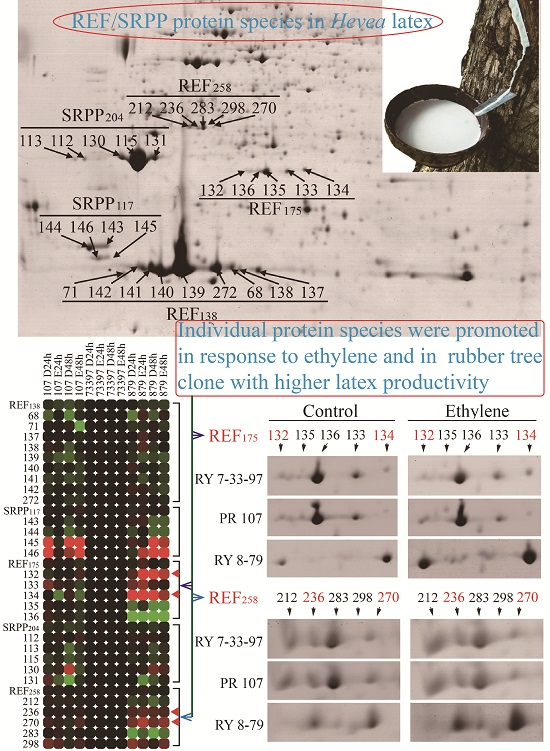
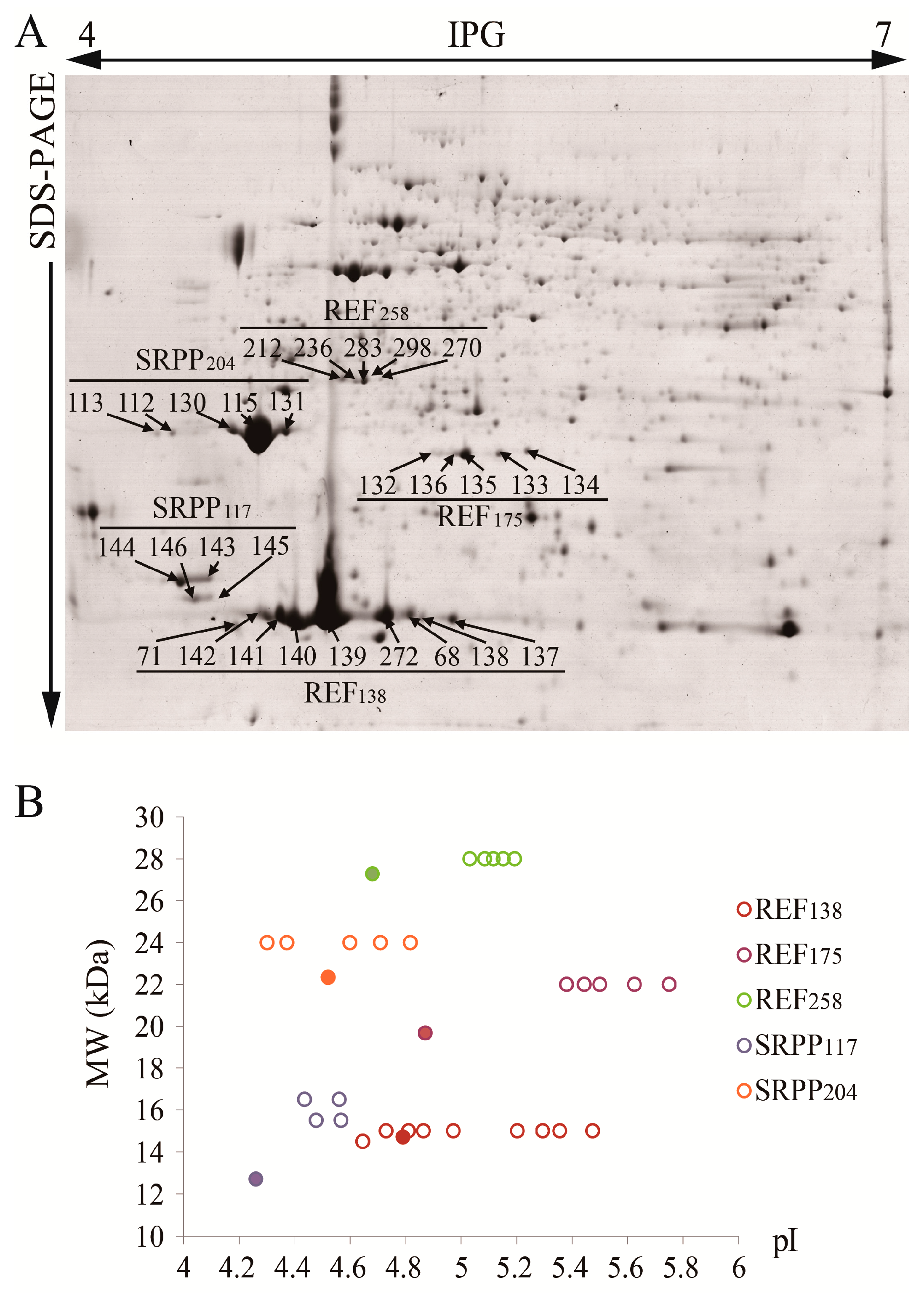
2.5. Identification of Protein Species of Each REF/SRPP Protein
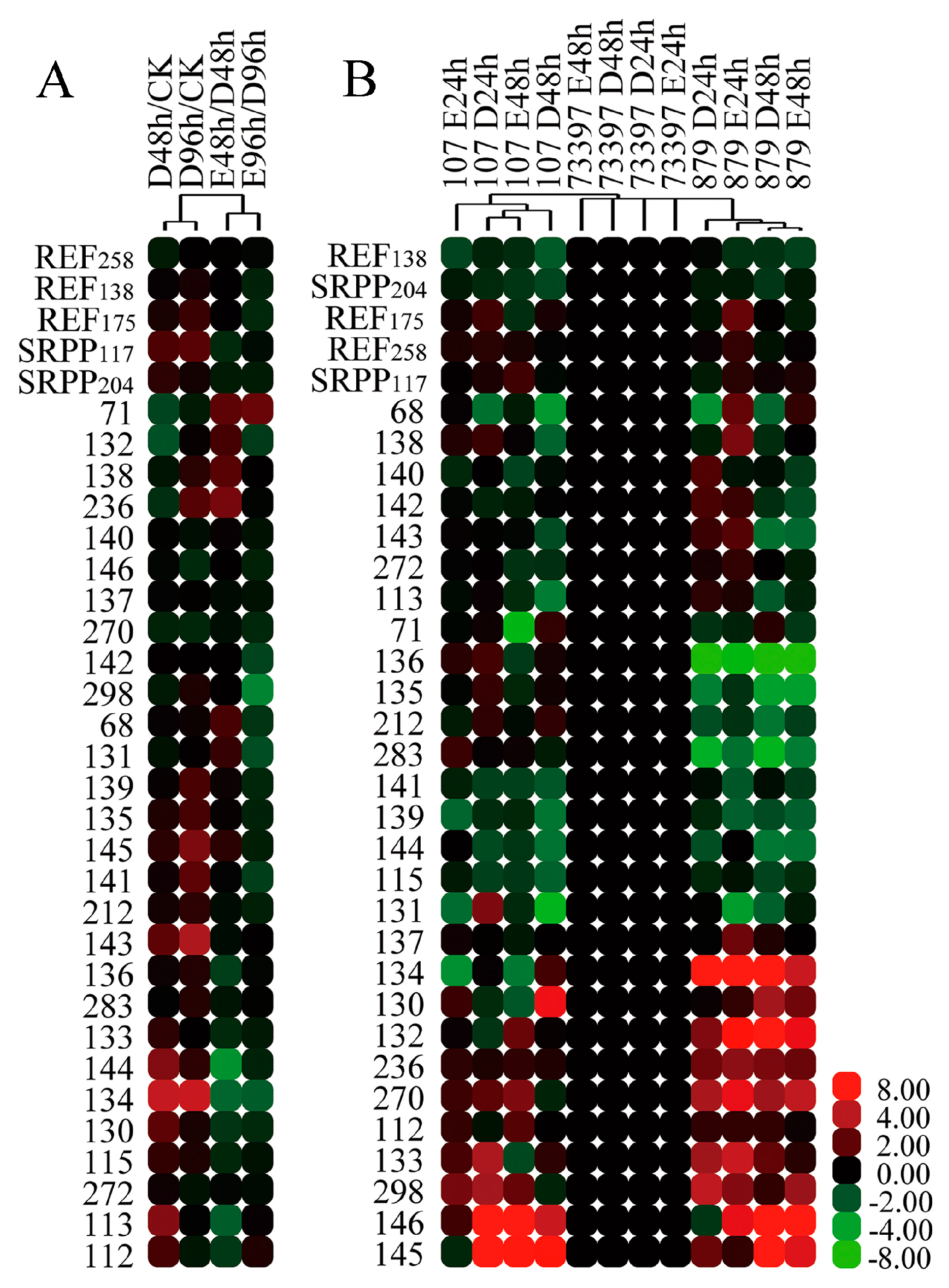
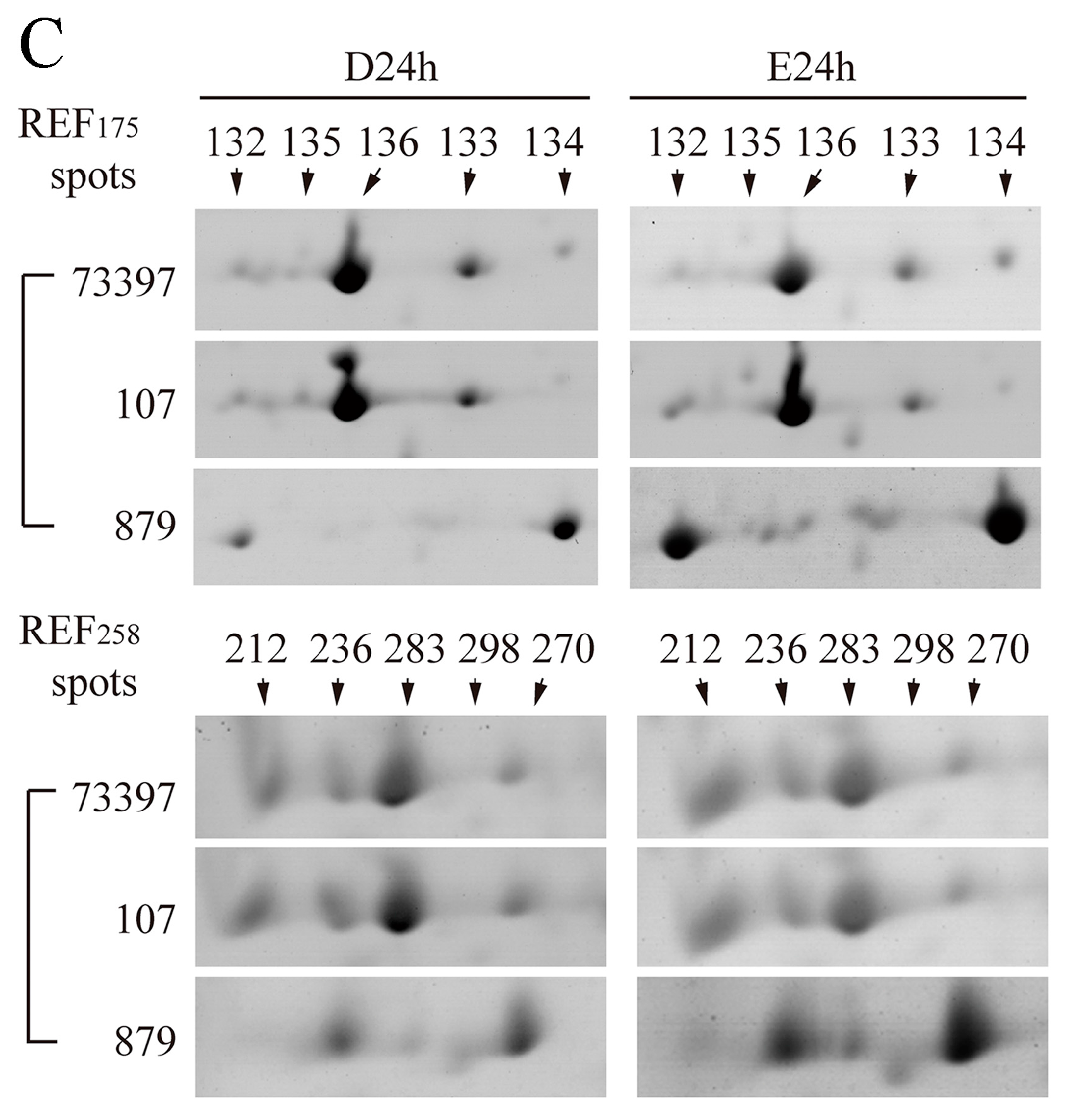
2.6. Characterization of REF/SRPP Protein Species in Different Rubber Tree Clones in Response to Tapping or Ethylene Treatment
3. Discussion
3.1. The REF/SRPP Family in H. brasiliensis
3.2. REF258 Is a Multiple-Domain Protein in REF/SRPP Family
3.3. The Multiple Protein Species of REF/SRPP in H. brasiliensis
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Gene Clone
4.3. Southern Blot Analysis
4.4. REF/SRPP-GFP Fusion Transient Expression
4.5. Quantitative PCR Analysis of REF and SRPP
4.6. Protein Extraction and 2-DE
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bonner, J. The history of rubber. In Guayule Natural Rubber: A Technical Publication with Emphasis on Recent Findings; Whitworth, J.W., Whitehead, E.E., Eds.; Guayule Administrative Management Committee and US Department of Agriculture Cooperative State Research Service, Office of Arid Lands Studies, University of Arizona: Tucson, AZ, USA, 1991; pp. 1–16. [Google Scholar]
- Hagel, J.M.; Yeung, E.C.; Facchini, P.J. Got milk? The secret life of laticifers. Trends Plant Sci. 2008, 13, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Nor, H.M.; Ebdon, J.R. Telechelic liquid natural rubber: A review. Prog. Polym. Sci. 1998, 23, 143–177. [Google Scholar] [CrossRef]
- Sakdapipanich, J.T. Structural characterization of natural rubber based on recent evidence from selective enzymatic treatments. J. Biosci. Bioeng. 2007, 103, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Cornish, K.; Backhaus, R. Rubber transferase activity in rubber particles of guayule. Phytochemistry 1990, 29, 3809–3813. [Google Scholar] [CrossRef]
- Cornish, K.; Wood, D.F.; Windle, J.J. Rubber particles from four different species, examined by transmission electron microscopy and electron-paramagnetic-resonance spin labeling, are found to consist of a homogeneous rubber core enclosed by a contiguous, monolayer biomembrane. Planta 1999, 210, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Archer, B.L.; Audley, B.G.; Cockbain, E.G.; McSweeney, G.P. The biosynthesis of rubber. Incorporation of mevalonate and isopentenyl pyrophosphate into rubber by Hevea brasiliensis-latex fractions. Biochem. J. 1963, 89, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.R.; Madhavan, S.; Greenblatt, G.A.; Venkatachalam, K.V.; Foster, M.A. The enzymatic synthesis of rubber polymer in Parthenium argentatum Gray. Plant Physiol. 1990, 92, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Yamaguchi, H.; Waki, T.; Aoki, Y.; Mizuno, M.; Yanbe, F.; Ishii, T.; Funaki, A.; Tozawa, Y.; Miyagi-Inoue, Y.; et al. Identification and reconstitution of the rubber biosynthetic machinery on rubber particles from Hevea brasiliensis. eLife 2016, 5, e19022. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.S.; Light, D.R. Rubber elongation factor from Hevea brasiliensis. Identification, characterization, and role in rubber biosynthesis. J. Biol. Chem. 1989, 264, 18608–18613. [Google Scholar] [PubMed]
- Oh, S.K.; Kang, H.; Shin, D.H.; Yang, J.; Chow, K.S.; Yeang, H.Y.; Wagner, B.; Breiteneder, H.; Han, K.H. Isolation, characterization and functional analysis of a novel cDNA clone encoding a small rubber particle protein from Hevea brasiliensis. J. Biol. Chem. 1999, 274, 17132–17138. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.Z.; Wu, J.L. Laticifer Differentiation in Hevea brasiliensis: Induction by exogenous jasmonic acid and linolenic acid. Ann. Bot. 2000, 85, 37–43. [Google Scholar] [CrossRef]
- Sando, T.; Hayashi, T.; Takeda, T.; Akiyama, Y.; Nakazawa, Y.; Fukusaki, E.; Kobayashi, A. Histochemical study of detailed laticifer structure and rubber biosynthesis-related protein localization in Hevea brasiliensis using spectral confocal laser scanning microscopy. Planta 2009, 230, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, K.; Lecomte, S.; Estevez, Y.; Coulary-Salin, B.; Bentaleb, A.; Cullin, C.; Deffieux, A.; Peruch, F. Rubber elongation factor (REF), a major allergen component in Hevea brasiliensis latex has amyloid properties. PLoS ONE 2012, 7, e48065. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, K.; Lecomte, S.; Estevez, Y.; Zhendre, V.; Henry, S.; Thevenot, J.; Dufourc, E.J.; Alves, I.D.; Peruch, F. Rubber particle proteins, HbREF and HbSRPP, show different interactions with model membranes. Biochim. Biophys. Acta 2014, 1838, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.R.; Yang, M.; Fang, Y.J.; Luo, Y.F.; Gao, S.H.; Xiao, X.H.; An, Z.; Zhou, B.; Zhang, B.; Tan, X.; et al. The rubber tree genome reveals new insights into rubber production and species adaptation. Nat. Plants 2016, 2, 16073. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.Q.; Tong, Z.; Jin, X.; Wang, D.; Sun, Y.; Meng, X.R.; Wang, X.C. Cloning and Expression of REF/SRPP Gene Family in Rubber Tree (Hevea brasiliensis). Mol. Plant Breed. 2016, 14, 3024–3032. [Google Scholar]
- Sookmark, U.; Pujade-Renaud, V.; Chrestin, H.; Lacote, R.; Naiyanetr, C.; Seguin, M.; Romruensukharom, P.; Narangajavana, J. Characterization of polypeptides accumulated in the latex cytosol of rubber trees affected by the tapping panel dryness syndrome. Plant Cell Physiol. 2002, 43, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Nie, Z.; Kang, G.; Li, Y.; Zeng, R. Identification and subcellular localization analysis of two rubber elongation factor isoforms on Hevea brasiliensis rubber particles. Plant Physiol. Biochem. 2017, 111, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Park, K.Y.; Seo, Y.S.; Kim, W.T. Arabidopsis small rubber particle protein homolog SRPs play dual roles as positive factors for tissue growth and development and in drought stress responses. Plant Physiol. 2016, 170, 2494–2510. [Google Scholar] [CrossRef] [PubMed]
- Gidda, S.K.; Park, S.J.; Pyc, M.; Yurchenko, O.; Cai, Y.Q.; Wu, P.; Andrews, D.W.; Chapman, K.D.; Dyer, J.M.; Mullen, R.T. Lipid droplet-associated proteins (LDAPs) are required for the 19 he dynamic regulation 20 of neutral lipid compartmentation in plant cells. Plant Physiol. 2016, 170, 2052–2071. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Wang, D.; Sun, Y.; Yang, Q.; Chang, L.L.; Wang, L.M.; Meng, X.; Huang, Q.; Jin, X.; Tong, Z. Comprehensive proteomics analysis of laticifer latex reveals new insights into ethylene stimulation of natural rubber production. Sci. Rep. 2015, 5, 13778. [Google Scholar] [CrossRef] [PubMed]
- Priya, P.; Venkatachalam, P.; Thulaseedharan, A. Differential expression pattern of rubber elongation factor (REF) mRNA transcripts from high and low yielding clones of rubber tree (Hevea brasiliensis Muell. Arg.). Plant Cell Rep. 2007, 26, 1833–1838. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.B.; Qin, Y.X.; Tang, C.R. Physiological characters of latex from three Hevea clones. Reyan: 8-79, Reyan 7-33-97 and PR107. J. Trop. Subtrop. Bot. 2010, 18, 170–175. [Google Scholar]
- An, F.; Zou, Z.; Cai, X.; Wang, J.; Rookes, J.; Lin, W.; Cahill, D.; Kong, L. Regulation of HbPIP2;3, a latex-abundant water transporter, is associated with latex dilution and yield in the rubber tree (Hevea brasiliensis Muell. Arg.). PLoS ONE 2015, 10, e0125595. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.Q.; Chen, Y.Y.; Wu, S.H.; Tian, W.M. Comparative transcriptome analysis of latex from rubber tree clone CATAS8–79 and PR107 reveals new cues for the regulation of latex regeneration and duration of latex flow. BMC Plant Biol. 2015, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Laibach, N.; Hillebrand, A.; Twyman, R.M.; Prüfer, D.; Gronover, C.S. Identification of a Taraxacum brevicorniculatum rubber elongation factor protein that is localized on rubber particles and promotes rubber biosynthesis. Plant J. 2015, 82, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Abdin, M.Z.; Kiran, U.; Alam, A. Analysis of osmotin, a PR protein as metabolic modulator in plants. Bioinformation 2011, 5, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, K.; Lecomte, S.; Estevez, Y.; Peruch, F. Hevea brasiliensis REF (Hev b 1) and SRPP (Hev b 3): An overview on rubber particle proteins. Biochimie 2014, 106, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wadeesirisak, K.; Castano, S.; Berthelot, K.; Vaysse, L.; Bonfils, F.; Peruch, F.; Rattanaporn, K.; Liengprayoon, S.; Lecomte, S.; Bottier, C. Rubber particle proteins REF1 and SRPP1 interact differently with native lipids extracted from Hevea brasiliensis latex. Biochim. Biophys. Acta 2017, 1859, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Horn, P.J.; James, C.N.; Gidda, S.K.; Kilaru, A.; Dyer, J.M.; Mullen, R.T.; Ohlrogge, J.B.; Chapman, K.D. Identification of a new class of lipid droplet-associated proteins in plants. Plant Physiol. 2013, 162, 1926–1936. [Google Scholar] [CrossRef] [PubMed]
- D’Auzac, J.; Ribaillier, D. Ethylene, a new agent stimulating latex production in Hevea brasiliensis. CR Hebd. Seances Acad. Sci. D 1969, 268, 3046–30499. [Google Scholar]
- Liu, J.P.; Zhuang, Y.F.; Guo, X.L.; Li, Y.J. Molecular mechanism of ethylene stimulation of latex yield in rubber tree (Hevea brasiliensis) revealed by de novo sequencing and transcriptome analysis. BMC Genom. 2016, 17, 257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Chen, J.Y. A method for identifying discriminative isoform-specific peptides for clinical proteomics application. BMC Genom. 2016, 17, 522. [Google Scholar] [CrossRef] [PubMed]
- Cooke, A.R.; Randall, D.I. 2-Haloethanephosphonic acids as ethylene releasing agents for the induction of flowering in pineapples. Nature 1968, 218, 974–975. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.D.; Wycherley, P.R.; Pakianathan, S.W. Stimulation of latex flow in Hevea brasiliensis by 4-Amino—3,5,6-Trichloropicolinic acid and 2-chloroethanephosphonic acid. Rubber Chem. Technol. 1972, 45, 883–899. [Google Scholar] [CrossRef]
- Audley, B.G.; Archer, B.L.; Carruthers, I.B. Metabolism of ethephon (2-chloroethylphosphonic acid) and related compounds in Hevea brasiliensis. Arch. Environ. Contam. Toxicol. 1976, 4, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Shi, M.J.; Wang, D.; Chen, Y.Y.; Cai, F.G.; Zhang, S.X.; Wang, L.; Tong, Z.; Tian, W.M. Comparative proteomics of primary and secondary lutoids reveals that chitinase and glucanase play a crucial combined role in rubber particle aggregation in Hevea brasiliensis. J. Proteome Res. 2013, 12, 5146–5159. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(t) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Shi, M.J.; Lu, X.L.; Ma, R.F.; Wu, C.G.; Guo, A.P.; Peng, M.; Tian, W. A method for protein extraction from different subcellular fractions of laticifer latex in Hevea brasiliensis compatible with 2-DE and MS. Proteome Sci. 2010, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, Z.; Wang, D.; Sun, Y.; Yang, Q.; Meng, X.; Wang, L.; Feng, W.; Li, L.; Wurtele, E.S.; Wang, X. Comparative Proteomics of Rubber Latex Revealed Multiple Protein Species of REF/SRPP Family Respond Diversely to Ethylene Stimulation among Different Rubber Tree Clones. Int. J. Mol. Sci. 2017, 18, 958. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18050958
Tong Z, Wang D, Sun Y, Yang Q, Meng X, Wang L, Feng W, Li L, Wurtele ES, Wang X. Comparative Proteomics of Rubber Latex Revealed Multiple Protein Species of REF/SRPP Family Respond Diversely to Ethylene Stimulation among Different Rubber Tree Clones. International Journal of Molecular Sciences. 2017; 18(5):958. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18050958
Chicago/Turabian StyleTong, Zheng, Dan Wang, Yong Sun, Qian Yang, Xueru Meng, Limin Wang, Weiqiang Feng, Ling Li, Eve Syrkin Wurtele, and Xuchu Wang. 2017. "Comparative Proteomics of Rubber Latex Revealed Multiple Protein Species of REF/SRPP Family Respond Diversely to Ethylene Stimulation among Different Rubber Tree Clones" International Journal of Molecular Sciences 18, no. 5: 958. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18050958