
Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals

 ,
,
Abstract
:
1. Introduction
1.1. Free Radicals on Reproduction (ROS/RNS)
1.2. Oxidative Stress
1.3. Nitrosative Stress
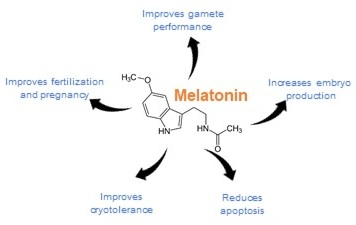
1.4. Melatonin and ROS/RNS
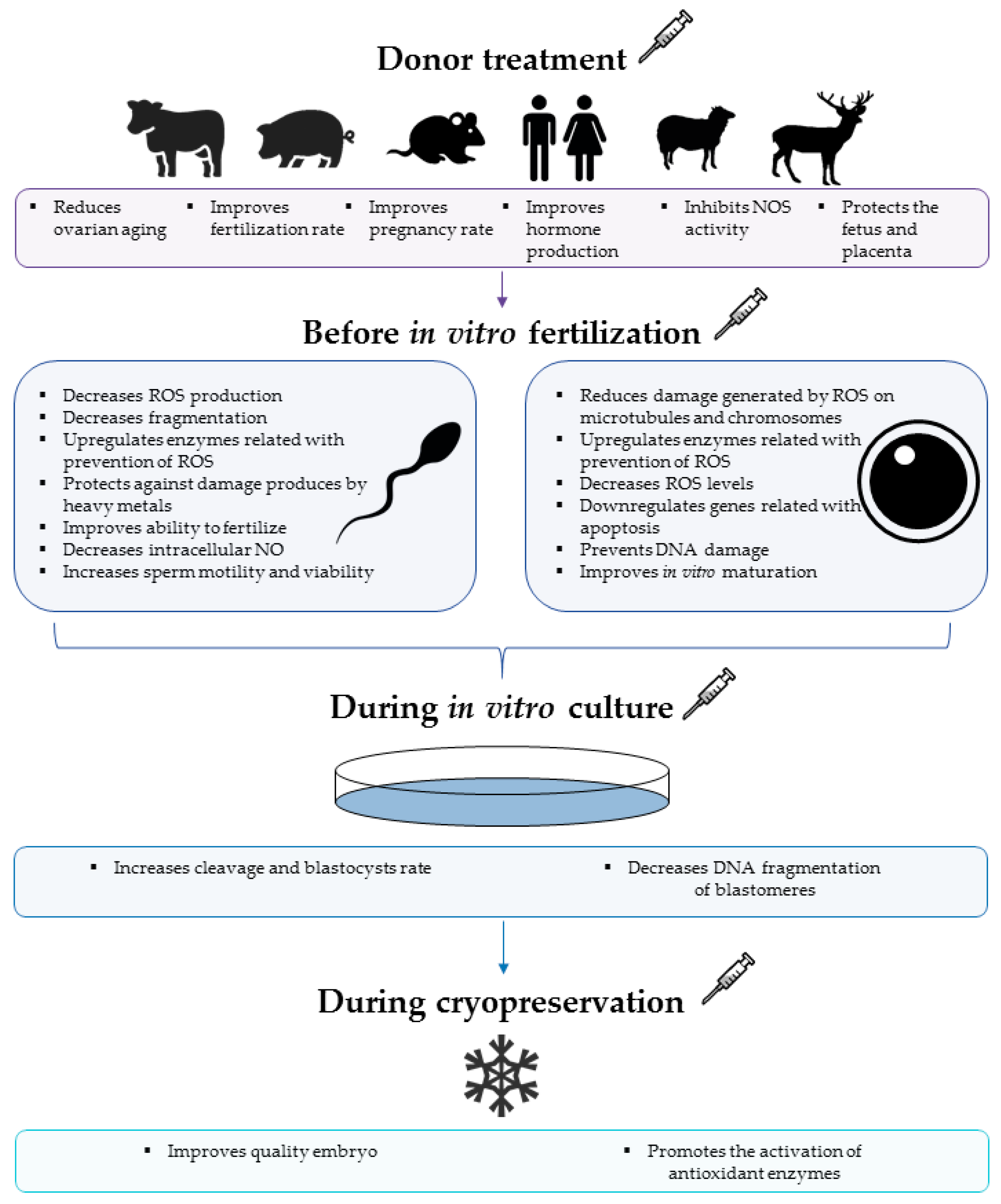
2. Melatonin Modulates Oxidative Stress on Gametes and In Vitro Embryo Production (IVP)
3. Potential Use of Melatonin against Nitrosative Stress during ART
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative stress in an assisted reproductive techniques setting. Fertil. Steril. 2006, 86, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fiskum, G.; Schubert, D. Generation of reactive oxygen species by the mitochondrial electron transport chain. J. Neurochem. 2002, 80, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Pasqualotto, E.B.; Agarwal, A.; Sharma, R.K.; Izzo, V.M.; Pinotti, J.A.; Joshi, N.J.; Rose, B.I. Effect of oxidative stress in follicular fluid on the outcome of assisted reproductive procedures. Fertil. Steril. 2004, 81, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Attaran, M.; Pasqualotto, E.; Falcone, T.; Goldberg, J.M.; Miller, K.F.; Agarwal, A.; Sharma, R.K. The effect of follicular fluid reactive oxygen species on the outcome of in vitro fertilization. Int. J. Fertil. Womens Med. 2000, 45, 314–320. [Google Scholar] [PubMed]
- Morado, S.A.; Cetica, P.D.; Beconi, M.T.; Dalvit, G.C. Reactive oxygen species in bovine oocyte maturation in vitro. Reprod. Fertil. Dev. 2009, 21, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Khatun, S.; Pandey, A.N.; Mishra, S.K.; Chaube, R.; Shrivastav, T.G.; Chaube, S.K. Intracellular levels of hydrogen peroxide and nitric oxide in oocytes at various stages of meiotic cell cycle and apoptosis. Free Radic. Res. 2009, 43, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Chaube, S.K.; Khatun, S.; Misra, S.K.; Shrivastav, T.G. Calcium ionophore-induced egg activation and apoptosis are associated with the generation of intracellular hydrogen peroxide. Free Radic. Res. 2008, 42, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Shkolnik, K.; Tadmor, A.; Ben-Dor, S.; Nevo, N.; Galiani, D.; Dekel, N. Reactive oxygen species are indispensable in ovulation. Proc. Natl. Acad. Sci. USA 2011, 108, 1462–1467. [Google Scholar] [CrossRef] [PubMed]
- De Lamirande, E.; Gagnon, C. A positive role for the superoxide anion in triggering hyperactivation and capacitation of human spermatozoa. Int. J. Androl. 1993, 16, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Bize, I.; Santander, G.; Cabello, P.; Driscoll, D.; Sharpe, C. Hydrogen peroxide is involved in hamster sperm capacitation in vitro. Biol. Reprod. 1991, 44, 398–403. [Google Scholar] [CrossRef] [PubMed]
- De Lamirande, E.; Gagnon, C. Human sperm hyperactivation and capacitation as parts of an oxidative process. Free Radic. Biol. Med. 1993, 14, 157–166. [Google Scholar] [CrossRef]
- Leclerc, P.; de Lamirande, E.; Gagnon, C. Interaction between Ca2+, cyclic 3′,5′ adenosine monophosphate, the superoxide anion, and tyrosine phosphorylation pathways in the regulation of human sperm capacitation. J. Androl. 1998, 19, 434–443. [Google Scholar] [PubMed]
- Leclerc, P.; de Lamirande, E.; Gagnon, C. Regulation of protein-tyrosine phosphorylation and human sperm capacitation by reactive oxygen derivatives. Free Radic. Biol. Med. 1997, 22, 643–656. [Google Scholar] [CrossRef]
- Roy, S.C.; Atreja, S.K. Effect of reactive oxygen species on capacitation and associated protein tyrosine phosphorylation in buffalo (Bubalus bubalis) spermatozoa. Anim. Reprod. Sci. 2008, 107, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Dona, G.; Fiore, C.; Tibaldi, E.; Frezzato, F.; Andrisani, A.; Ambrosini, G.; Fiorentin, D.; Armanini, D.; Bordin, L.; Clari, G. Endogenous reactive oxygen species content and modulation of tyrosine phosphorylation during sperm capacitation. Int. J. Androl. 2011, 34, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Rivlin, J.; Mendel, J.; Rubinstein, S.; Etkovitz, N.; Breitbart, H. Role of hydrogen peroxide in sperm capacitation and acrosome reaction. Biol. Reprod. 2004, 70, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Sulieman, F.J.; Li, J.; Wei, Q.; Xu, M.; Shi, F. Nitric oxide and thyroid hormone receptor alpha 1 contribute to ovarian follicular development in immature hyper- and hypo-thyroid rats. Reprod. Biol. 2015, 15, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Dubey, P.K.; Tripathi, V.; Singh, R.P.; Saikumar, G.; Nath, A.; Pratheesh, M.D.; Gade, N.; Sharma, G.T. Expression of nitric oxide synthase isoforms in different stages of buffalo (Bubalus bubalis) ovarian follicles: Effect of nitric oxide on in vitro development of preantral follicle. Theriogenology 2012, 77, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Bu, S.; Xia, G.; Tao, Y.; Lei, L.; Zhou, B. Dual effects of nitric oxide on meiotic maturation of mouse cumulus cell-enclosed oocytes in vitro. Mol. Cell. Endocrinol. 2003, 207, 21–30. [Google Scholar] [CrossRef]
- Viana, K.S.; Caldas-Bussiere, M.C.; Matta, S.G.; Faes, M.R.; de Carvalho, C.S.; Quirino, C.R. Effect of sodium nitroprusside, a nitric oxide donor, on the in vitro maturation of bovine oocytes. Anim. Reprod. Sci. 2007, 102, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Jablonka-Shariff, A.; Olson, L.M. The role of nitric oxide in oocyte meiotic maturation and ovulation: Meiotic abnormalities of endothelial nitric oxide synthase knock-out mouse oocytes. Endocrinology 1998, 139, 2944–2954. [Google Scholar] [CrossRef] [PubMed]
- Pires, P.R.L.; Santos, N.P.; Adona, P.R.; Natori, M.M.; Schwarz, K.R.L.; de Bem, T.H.C.; Leal, C.L.V. Endothelial and inducible nitric oxide synthases in oocytes of cattle. Anim. Reprod. Sci. 2009, 116, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Tranguch, S.; Steuerwald, N.; Huet-Hudson, Y.M. Nitric oxide synthase production and nitric oxide regulation of preimplantation embryo development. Biol. Reprod. 2003, 68, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, D.; Kadanga, A.; Rings, F.; Bauch, K.; Jennen, D.; Nganvongpanit, K.; Holker, M.; Tholen, E.; Ponsuksili, S.; Wimmers, K.; et al. The effect of nitric oxide inhibition and temporal expression patterns of the mRNA and protein products of nitric oxide synthase genes during in vitro development of bovine pre-implantation embryos. Reprod. Domest. Anim. 2006, 41, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Roessner, C.; Paasch, U.; Glander, H.J.; Grunewald, S. Activity of nitric oxide synthase in mature and immature human spermatozoa. Andrologia 2010, 42, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.C.; Valdez, L.B.; Zaobornyj, T.; Boveris, A.; Beconi, M.T. Nitric oxide and superoxide anion production during heparin-induced capacitation in cryopreserved bovine spermatozoa. Reprod. Domest. Anim. 2011, 46, 74–81. [Google Scholar] [CrossRef] [PubMed]
- De Lamirande, E.; Lamothe, G. Reactive oxygen-induced reactive oxygen formation during human sperm capacitation. Free Radic. Biol. Med. 2009, 46, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.B.; de Lamirande, E.; Gagnon, C. Nitric oxide is a signaling molecule in spermatozoa. Curr. Pharm. Des. 2003, 9, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Transcriptional responses to oxidative stress: Pathological and toxicological implications. Pharmacol. Ther. 2010, 125, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Devlin, T.M. Bioquímica: Libro de Texto con Aplicaciones Clínicas, 3rd ed.; Revert: Barcelona, Spain, 2004; Volume 1, pp. 217–266. [Google Scholar]
- Pourova, J.; Kottova, M.; Voprsalova, M.; Pour, M. Reactive oxygen and nitrogen species in normal physiological processes. Acta Physiol. 2010, 198, 15–35. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M. Oxidative Stress and Redox Regulation on In Vitro development of mammalian embryos. J. Reprod. Dev. 2012, 58, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; He, L.; Liu, Y.; Shi, Y.; Du, H. The expression and role of oxidative stress markers in the serum and follicular fluid of patients with endometriosis. Clin. Exp. Obstet. Gynecol. 2013, 40, 372–376. [Google Scholar] [PubMed]
- Lee, T.H.; Lee, M.S.; Liu, C.H.; Tsao, H.M.; Huang, C.C.; Yang, Y.S. The association between microenvironmental reactive oxygen species and embryo development in assisted reproduction technology cycles. Reprod. Sci. 2012, 19, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Bedaiwy, M.A.; Elnashar, S.A.; Goldberg, J.M.; Sharma, R.; Mascha, E.J.; Arrigain, S.; Agarwal, A.; Falcone, T. Effect of follicular fluid oxidative stress parameters on intracytoplasmic sperm injection outcome. Gynecol. Endocrinol. 2012, 28, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Rajani, S.; Chattopadhyay, R.; Goswami, S.K.; Ghosh, S.; Sharma, S.; Chakravarty, B. Assessment of oocyte quality in polycystic ovarian syndrome and endometriosis by spindle imaging and reactive oxygen species levels in follicular fluid and its relationship with IVF-ET outcome. J. Hum. Reprod. Sci. 2012, 5, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Loren, P.; Cheuquemán, C.; Risopatrón, J.; Felmer, R.; Arias, M.E.; Sánchez, R. Modulación del Estado de Óxido-Reducción por Peróxido de Hidrógeno en la Etapa de Maduración Ovocitaria: Efecto sobre el Desarrollo Embrionario en Bovinos. Int. J. Morphol. 2016, 34, 431–435. [Google Scholar] [CrossRef]
- Loren, P.; Sanchez, E.; Risopatrón, J.; Arias, M.; Felmer, R.; Sánchez, R. Efecto de estrés oxidativo in vitro en complejos ovocito-cumulus: Evaluación del potencial de desarrollo embrionario, calidad y perfil de expresión génica en bovinos. In Proceedings of the VII Congreso de la Asociación Iberoamericana de Sociedades de Andrología, Lisboa, Portugal, 3–5 December 2016. [Google Scholar]
- Pandey, A.N.; Tripathi, A.; PremKumar, K.V.; Shrivastav, T.G.; Chaube, S.K. Reactive oxygen and nitrogen species during meiotic resumption from diplotene arrest in mammalian oocytes. J. Cell. Biochem. 2010, 111, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Aitken, R.J. The importance of redox regulated pathways in sperm cell biology. Mol. Cell. Endocrinol. 2004, 216, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Blanco, S.; Hernández, R.; Franchelli, G.; Ramos-Álvarez, M.M.; Peinado, M.Á. Melatonin influences NO/NOS pathway and reduces oxidative and nitrosative stress in a model of hypoxic-ischemic brain damage. Nitric Oxide 2017, 62, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, S.S.; Hagenaar, K.; Lampiao, F. The in vitro effects of melatonin on human sperm function and its scavenging activities on NO and ROS. Andrologia 2010, 42, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Loren, P.; Cheuqueman, C.; Sanchez, E.; Risopatron, J.; Arias, M.E.; Felmer, R.; Sanchez, R. Effect of short-term exposure of cumulus-oocyte complex to 3-morpholinosydnonimine on in vitro embryo development and gene expression in cattle. Reprod. Domest. Anim. 2016, 51, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Cheuqueman, C.; Loren, P.; Arias, M.; Risopatron, J.; Felmer, R.; Alvarez, J.; Mogas, T.; Sanchez, R. Effects of short-term exposure of mature oocytes to sodium nitroprusside on in vitro embryo production and gene expression in bovine. Theriogenology 2015, 84, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [PubMed]
- Gonzalez-Arto, M.; Hamilton, T.R.D.S.; Gallego, M.; Gaspar-Torrubia, E.; Aguilar, D.; Serrano-Blesa, E.; Abecia, J.A.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; et al. Evidence of melatonin synthesis in the ram reproductive tract. Andrology 2016, 4, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.-X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Venegas, C.; Garcia, J.A.; Escames, G.; Ortiz, F.; Lopez, A.; Doerrier, C.; Garcia-Corzo, L.; Lopez, L.C.; Reiter, R.J.; Acuna-Castroviejo, D. Extrapineal melatonin: Analysis of its subcellular distribution and daily fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.D.; Mora-Santos, M.; Naji, L.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Calvo, J.R. Evidence of melatonin synthesis and release by mast cells. Possible modulatory role on inflammation. Pharmacol. Res. 2010, 62, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Barrett, P.; Bolborea, M. Molecular pathways involved in seasonal body weight and reproductive responses governed by melatonin. J. Pineal Res. 2012, 52, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Zarazaga, L.A.; Celi, I.; Guzman, J.L.; Malpaux, B. Melatonin concentrations in the two jugular veins, and relationship with the seasonal reproductive activity in goats. Theriogenology 2010, 74, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and reproduction revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Eryilmaz, O.G.; Devran, A.; Sarikaya, E.; Aksakal, F.N.; Mollamahmutoğlu, L.; Cicek, N. Melatonin improves the oocyte and the embryo in IVF patients with sleep disturbances, but does not improve the sleeping problems. J. Assist. Reprod. Genet. 2011, 28, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Dahl, G.E.; Buchanan, B.A.; Tucker, H.A. Photoperiodic effects on dairy cattle: A review. J. Dairy Sci. 2000, 83, 885–893. [Google Scholar] [CrossRef]
- Voiculescu, S.E.; Zygouropoulos, N.; Zahiu, C.D.; Zagrean, A.M. Role of melatonin in embryo fetal development. J. Med. Life 2014, 7, 488–492. [Google Scholar] [PubMed]
- Refsdal, A.O. Reproductive performance of Norwegian cattle from 1985 to 2005: Trends and seasonality. Acta Vet. Scand. 2007, 49, 5. [Google Scholar] [CrossRef] [PubMed]
- Cheuquemán, C.; Loren, P.; Arias, M.; Risopatrón, J.; Felmer, R.; Álvarez, J.; Mogas, T.; Sánchez, R. Decrease in bovine in vitro embryo production efficiency during winter season in a warm-summer Mediterranean climate. Andrologia 2016. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Tokunaga, K.; Kakegawa, R.; Yanagisawa, A.; Takemoto, A.; Tatemizo, A.; Watanabe, T.; Hatanaka, Y.; Matsushita, A.; Kishi, M.; et al. Expression analysis of circadian genes in oocytes and preimplantation embryos of cattle and rabbits. Anim. Reprod. Sci. 2010, 121, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Matsushita, A.; Hatanaka, Y.; Watanabe, T.; Oishi, K.; Ishida, N.; Anzai, M.; Mitani, T.; Kato, H.; Kishigami, S.; et al. Expression and functional analyses of circadian genes in mouse oocytes and preimplantation embryos: Cry1 is involved in the meiotic process independently of circadian clock regulation. Biol. Reprod. 2009, 80, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Qi, W. Biochemical reactivity of melatonin with reactive oxygen and nitrogen species: A review of the evidence. Cell Biochem. Biophys. 2001, 34, 237–256. [Google Scholar] [CrossRef]
- Antolin, I.; Rodriguez, C.; Sainz, R.M.; Mayo, J.C.; Uria, H.; Kotler, M.L.; Rodriguez-Colunga, M.J.; Tolivia, D.; Menendez-Pelaez, A. Neurohormone melatonin prevents cell damage: Effect on gene expression for antioxidant enzymes. FASEB J. 1996, 10, 882–890. [Google Scholar] [PubMed]
- Poeggeler, B.; Reiter, R.J.; Hardeland, R.; Tan, D.X.; Barlow-Walden, L.R. Melatonin and structurally-related, endogenous indoles act as potent electron donors and radical scavengers in vitro. Redox Rep. Commun. Free Radic. Res. 1996, 2, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Squadrito, G.L.; Pryor, W.A. The reaction of melatonin with peroxynitrite: Formation of melatonin radical cation and absence of stable nitrated products. Biochem. Biophys. Res. Commun. 1998, 251, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Barlow-Walden, L.R.; Reiter, R.J.; Abe, M.; Pablos, M.; Menendez-Pelaez, A.; Chen, L.D.; Poeggeler, B. Melatonin stimulates brain glutathione peroxidase activity. Neurochem. Int. 1995, 26, 497–502. [Google Scholar] [CrossRef]
- Mayo, J.C.; Sainz, R.M.; Antoli, I.; Herrera, F.; Martin, V.; Rodriguez, C. Melatonin regulation of antioxidant enzyme gene expression. Cell. Mol. Life Sci. 2002, 59, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nagai, R.; Watanabe, K.; Wakatsuki, A.; Hamada, F.; Shinohara, K.; Hayashi, Y.; Imamura, R.; Fukaya, T. Melatonin preserves fetal growth in rats by protecting against ischemia/reperfusion-induced oxidative/nitrosative mitochondrial damage in the placenta. J. Pineal Res. 2008, 45, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Ressmeyer, A.R.; Mayo, J.C.; Zelosko, V.; Sainz, R.M.; Tan, D.X.; Poeggeler, B.; Antolin, I.; Zsizsik, B.K.; Reiter, R.J.; Hardeland, R. Antioxidant properties of the melatonin metabolite N1-acetyl-5-methoxykynuramine (AMK): Scavenging of free radicals and prevention of protein destruction. Redox Rep. Commun. Free Radic. Res. 2003, 8, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Terron, M.P.; Flores, L.J.; Czarnocki, Z. Melatonin and its metabolites: New findings regarding their production and their radical scavenging actions. Acta Biochim. Pol. 2007, 54, 1–9. [Google Scholar] [PubMed]
- Reiter, R.J.; Paredes, S.D.; Korkmaz, A.; Jou, M.-J.; Tan, D.-X. Melatonin combats molecular terrorism at the mitochondrial level. Interdiscip. Toxicol. 2008, 1, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Thakor, A.S.; Herrera, E.A.; Seron-Ferre, M.; Giussani, D.A. Melatonin and vitamin C increase umbilical blood flow via nitric oxide-dependent mechanisms. J. Pineal Res. 2010, 49, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.N.; Sullivan, P.G.; Hall, E.D. Peroxynitrite-mediated oxidative damage to brain mitochondria: Protective effects of peroxynitrite scavengers. J. Neurosci. Res. 2007, 85, 2216–2223. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, B.N.; Sadeghi, S.; Alipour, S.; Parsanezhad, M.E.; Alamdarloo, S.M. Effect of melatonin on the outcome of assisted reproductive technique cycles in women with diminished ovarian reserve: A double-blinded randomized clinical trial. Iran. J. Med. Sci. 2017, 42, 73–78. [Google Scholar] [PubMed]
- Arjmand, F.; Khanmohammadi, M.; Arasteh, S.; Mohammadzadeh, A.; Kazemnejad, S.; Akhondi, M.-M. Extended culture of encapsulated human blastocysts in alginate hydrogel containing decidualized endometrial stromal cells in the presence of melatonin. Mol. Biotechnol. 2016, 58, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Pacchiarotti, A.; Carlomagno, G.; Antonini, G.; Pacchiarotti, A. Effect of myo-inositol and melatonin versus myo-inositol, in a randomized controlled trial, for improving in vitro fertilization of patients with polycystic ovarian syndrome. Gynecol. Endocrinol. 2016, 32, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Karimfar, M.H.; Niazvand, F.; Haghani, K.; Ghafourian, S.; Shirazi, R.; Bakhtiyari, S. The protective effects of melatonin against cryopreservation-induced oxidative stress in human sperm. Int. J. Immunopathol. Pharmacol. 2015, 28, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, I.; Monllor, F.; Marchena, A.M.; Ortiz, A.; Lozano, G.; Jimenez, M.I.; Gaspar, P.; Garcia, J.F.; Pariente, J.A.; Rodriguez, A.B.; et al. Exogenous melatonin supplementation prevents oxidative stress-evoked DNA damage in human spermatozoa. J. Pineal Res. 2014, 57, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, T.; Hashimoto, S.; Ito, K.; Nakaoka, Y.; Matsumoto, K.; Hosoi, Y.; Morimoto, Y. Oral melatonin supplementation improves oocyte and embryo quality in women undergoing in vitro fertilization-embryo transfer. Gynecol. Endocrinol. 2014, 30, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.X.; Lee, S.; Taweechaipaisankul, A.; Kim, G.A.; Lee, B.C. Melatonin regulates lipid metabolism in porcine oocytes. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef] [PubMed]
- Majrashi, K.A.; Barakat, I.A.; Al-Himaidi, A.R.; Adham, K.A. Effect of exogenous melatonin treatment on the reproductive characteristics and progeny of male rats exposed to different periods from light and darkness. Physiol. Res. 2017, 62, e12388. [Google Scholar]
- Li, B.; He, X.; Zhuang, M.; Niu, B.; Wu, C.; Mu, H.; Tang, F.; Cui, Y.; Liu, W.; Zhao, B.; et al. Melatonin ameliorates busulfan-induced spermatogonial stem cell oxidative apoptosis in mouse testes. Antioxid. Redox Signal. 2017. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Lu, Y.; Zhang, M.; Miao, Y.; Zhou, C.; Cui, Z.; Xiong, B. Melatonin improves the fertilization ability of post-ovulatory aged mouse oocytes by stabilizing ovastacin and Juno to promote sperm binding and fusion. Hum. Reprod. 2017, 32, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.S.; Rocha-Frigoni, N.A.; Mingoti, G.Z.; Roth, Z.; Hansen, P.J. Modification of embryonic resistance to heat shock in cattle by melatonin and genetic variation in HSPA1L. J. Dairy Sci. 2016, 99, 9152–9164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, W.; Ma, Y.; Wang, D.; Zhao, X.; Zeng, C.; Zhang, M.; Zeng, X.; Meng, Q.; Zhou, G. Improved development by melatonin treatment after vitrification of mouse metaphase II oocytes. Cryobiology 2016, 73, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.W.; Sun, Y.Q.; Jiang, X.L.; Huang, Z.Q.; Zhao, S.J.; Du, W.H.; Hao, H.S.; Zhao, X.M.; Zhu, H.B. Protective effects of melatonin on bovine sperm characteristics and subsequent in vitro embryo development. Mol. Reprod. Dev. 2016, 83, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Cunha, M.C.; Mesquita, L.G.; Bressan, F.; Collado, M.D.; Balieiro, J.C.; Schwarz, K.R.; de Castro, F.C.; Watanabe, O.Y.; Watanabe, Y.F.; de Alencar Coelho, L.; et al. Effects of melatonin during IVM in defined medium on oocyte meiosis, oxidative stress, and subsequent embryo development. Theriogenology 2016, 86, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.W.; Sun, Y.Q.; Sun, W.J.; Du, W.H.; Hao, H.S.; Zhao, S.J.; Zhu, H.B. Melatonin inhibits paraquat-induced cell death in bovine preimplantation embryos. J. Pineal Res. 2016, 60, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Park, H.J.; Kim, J.W.; Park, S.Y.; Yang, S.G.; Jung, J.M.; Kim, M.J.; Koo, D.B. 173 Melatonin alleviates the endoplasmic reticulum stress through the regulating of unfolding protein response signaling during porcine oocyte maturation in vitro. Reprod. Fertil. Dev. 2016, 29, 195. [Google Scholar] [CrossRef] [PubMed]
- Remiao, M.H.; Lucas, C.G.; Domingues, W.B.; Silveira, T.; Barther, N.N.; Komninou, E.R.; Basso, A.C.; Jornada, D.S.; Beck, R.C.; Pohlmann, A.R.; et al. Melatonin delivery by nanocapsules during in vitro bovine oocyte maturation decreased the reactive oxygen species of oocytes and embryos. Reprod. Toxicol. 2016, 63, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Komninou, E.R.; Remiao, M.H.; Lucas, C.G.; Domingues, W.B.; Basso, A.C.; Jornada, D.S.; Deschamps, J.C.; Beck, R.C.; Pohlmann, A.R.; Bordignon, V.; et al. Effects of two types of melatonin-loaded nanocapsules with distinct supramolecular structures: Polymeric (nc) and lipid-core nanocapsules (lnc) on bovine embryo culture model. PLoS ONE 2016, 11, e0157561. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, J.; Zhang, Z.; Yang, M.; Li, Y.; Tian, X.; Ma, T.; Tao, J.; Zhu, K.; Song, Y.; et al. Mitochondria synthesize melatonin to ameliorate its function and improve mice oocyte's quality under in vitro conditions. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Luo, X.; Li, L.; Peng, Q.; Yang, Y.; Zhao, L.; Ma, M.; Hou, Z. The protective effects of melatonin against oxidative stress and inflammation induced by acute cadmium exposure in mice testis. Biol. Trace Element Res. 2016, 170, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Zhang, Y.; Jia, G.X.; Meng, Q.G.; Bunch, T.D.; Liu, G.S.; Zhu, S.E.; Xhou, G.B. Effect of melatonin supplementation on cryopreserved sperm quality in mouse. Cryo Lett. 2016, 37, 115–122. [Google Scholar]
- Nagina, G.; Asima, A.; Nemat, U.; Shamim, A. Effect of melatonin on maturation capacity and fertilization of Nili-Ravi buffalo (Bubalus bubalis) oocytes. Open Vet. J. 2016, 6, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Guo, J.; Choi, J.W.; Kim, N.H.; Cui, X.S. Effect and possible mechanisms of melatonin treatment on the quality and developmental potential of aged bovine oocytes. Reprod. Fertil. Dev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Mehaisen, G.M.; Saeed, A.M.; Gad, A.; Abass, A.O.; Arafa, M.; El-Sayed, A. Antioxidant capacity of melatonin on preimplantation development of fresh and vitrified rabbit embryos: Morphological and molecular aspects. PLoS ONE 2015, 10, e0139814. [Google Scholar] [CrossRef] [PubMed]
- Ganji, R.; Nabiuni, M.; Faraji, R. Development of mouse preantral follicle after in vitro culture in a medium containing melatonin. Cell J. 2015, 16, 546–553. [Google Scholar] [PubMed]
- Su, J.; Wang, Y.; Xing, X.; Zhang, L.; Sun, H.; Zhang, Y. Melatonin significantly improves the developmental competence of bovine somatic cell nuclear transfer embryos. J. Pineal Res. 2015, 59, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; He, C.; Zhu, K.; Xu, Z.; Ma, T.; Tao, J.; Liu, G. Melatonin protects porcine oocyte in vitro maturation from heat stress. J. Pineal Res. 2015, 59, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Do, L.T.; Shibata, Y.; Taniguchi, M.; Nii, M.; Nguyen, T.V.; Tanihara, F.; Takagi, M.; Otoi, T. Melatonin supplementation during in vitro maturation and development supports the development of porcine embryos. Reprod. Domest. Anim. 2015, 50, 1054–1058. [Google Scholar] [CrossRef] [PubMed]
- Cheuqueman, C.; Arias, M.E.; Risopatron, J.; Felmer, R.; Alvarez, J.; Mogas, T.; Sanchez, R. Supplementation of IVF medium with melatonin: Effect on sperm functionality and in vitro produced bovine embryos. Andrologia 2015, 47, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Mehaisen, G.M.; Saeed, A.M. In vitro development rate of preimplantation rabbit embryos cultured with different levels of melatonin. Zygote 2015, 23, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tian, X.; Zhou, Y.; Tan, D.; Zhu, S.; Dai, Y.; Liu, G. Melatonin improves the quality of in vitro produced (IVP) bovine embryos: Implications for blastocyst development, cryotolerance, and modifications of relevant gene expression. PLoS ONE 2014, 9, e93641. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tian, X.; Zhang, L.; Gao, C.; He, C.; Fu, Y.; Ji, P.; Li, Y.; Li, N.; Liu, G. Beneficial effects of melatonin on in vitro bovine embryonic development are mediated by melatonin receptor 1. J. Pineal Res. 2014, 56, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, F.; He, C.; Zhang, L.; Tan, D.; Reiter, R.J.; Xu, J.; Ji, P.; Liu, G. Beneficial effects of melatonin on bovine oocytes maturation: A mechanistic approach. J. Pineal Res. 2014, 57, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Niknafs, B.; Mehdipour, A.; Mohammadi Roushandeh, A. Melatonin improves development of early mouse embryos impaired by actinomycin-D and TNF-alpha. Iran. J. Reprod. Med. 2014, 12, 799–804. [Google Scholar] [PubMed]
- Minaii, B.; Moayeri, A.; Shokri, S.; Habibi Roudkenar, M.; Golmohammadi, T.; Malek, F.; Barbarestani, M. Melatonin improve the sperm quality in forced swimming test induced oxidative stress in nandrolone treated Wistar rats. Acta Med. Iran. 2014, 52, 496–504. [Google Scholar] [PubMed]
- Salehi, M.; Kato, Y.; Tsunoda, Y. Effect of melatonin treatment on developmental potential of somatic cell nuclear-transferred mouse oocytes in vitro. Zygote 2014, 22, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Pasciu, V.; Manca, M.E.; Chelucci, S.; Torres-Rovira, L.; Leoni, G.G.; Zinellu, A.; Carru, C.; Naitana, S.; Berlinguer, F. Dose-dependent effect of melatonin on postwarming development of vitrified ovine embryos. Theriogenology 2014, 81, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhuo, Z.Y.; Shi, W.Q.; Tan, D.X.; Gao, C.; Tian, X.Z.; Zhang, L.; Zhou, G.B.; Zhu, S.E.; Yun, P.; et al. Melatonin promotes superovulation in sika deer (Cervus nippon). Int. J. Mol. Sci. 2014, 15, 12107–12118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chai, M.; Tian, X.; Wang, F.; Fu, Y.; He, C.; Deng, S.; Lian, Z.; Feng, J.; Tan, D.X.; et al. Effects of melatonin on superovulation and transgenic embryo transplantation in small-tailed han sheep (Ovis aries). Neuro Endocrinol. Lett. 2013, 34, 294–301. [Google Scholar] [PubMed]
- Pang, Y.W.; An, L.; Wang, P.; Yu, Y.; Yin, Q.D.; Wang, X.H.; Xin, Z.; Qian, Z.; Yang, M.L.; Min, G.; et al. Treatment of porcine donor cells and reconstructed embryos with the antioxidant melatonin enhances cloning efficiency. J. Pineal Res. 2013, 54, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Bahadori, M.H.; Ghasemian, F.; Ramezani, M.; Asgari, Z. Melatonin effect during different maturation stages of oocyte and subsequent embryo development in mice. Iran. J. Reprod. Med. 2013, 11, 11–18. [Google Scholar] [PubMed]
- Cebrian-Serrano, A.; Salvador, I.; Raga, E.; Dinnyes, A.; Silvestre, M.A. Beneficial effect of melatonin on blastocyst in vitro production from heat-stressed bovine oocytes. Reprod. Domest. Anim. 2013, 48, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tian, X.; Zhang, L.; Tan, D.; Reiter, R.J.; Liu, G. Melatonin promotes the in vitro development of pronuclear embryos and increases the efficiency of blastocyst implantation in murine. J. Pineal Res. 2013, 55, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.H.; Leal, C.L.; Cruz, J.F.; Tan, D.X.; Reiter, R.J. Essential actions of melatonin in protecting the ovary from oxidative damage. Theriogenology 2014, 82, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, J.; Maitra, D.; Diamond, M.P.; Abu-Soud, H.M. Melatonin prevents hypochlorous acid-induced alterations in microtubule and chromosomal structure in metaphase-II mouse oocytes. J. Pineal Res. 2012, 53, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Peng, W.; Yin, S.; Zhao, J.; Fu, B.; Zhang, J.; Mao, T.; Wu, H.; Zhang, Y. Melatonin improves age-induced fertility decline and attenuates ovarian mitochondrial oxidative stress in mice. Sci. Rep. 2016, 6, 35165. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Taketani, T.; Matsuoka, A.; Yamagata, Y.; Shimamura, K.; et al. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Yin, C.; Gong, Y.; Liu, J.; Guo, H.; Zhao, R. Melatonin-induced increase of lipid droplets accumulation and in vitro maturation in porcine oocytes is mediated by mitochondrial quiescence. J. Cell Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.; Qin, L.; Reiter, R. Melatonin: A Mitochondrial Targeting Molecule Involving Mitochondrial Protection and Dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Sheng, S.; Sun, Y.; Li, H.; Li, W.P.; Zhang, C.; Chen, Z.J. Melatonin levels in follicular fluid as markers for IVF outcomes and predicting ovarian reserve. Reproduction 2017, 153, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; et al. Melatonin as a free radical scavenger in the ovarian follicle. Endocr. J. 2013, 60, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Espino, J.; Bejarano, I.; Ortiz, A.; Lozano, G.M.; Garcia, J.F.; Pariente, J.A.; Rodriguez, A.B. Melatonin as a potential tool against oxidative damage and apoptosis in ejaculated human spermatozoa. Fertil. Steril. 2010, 94, 1915–1917. [Google Scholar] [CrossRef] [PubMed]
- Espino, J.; Ortiz, A.; Bejarano, I.; Lozano, G.M.; Monllor, F.; Garcia, J.F.; Rodriguez, A.B.; Pariente, J.A. Melatonin protects human spermatozoa from apoptosis via melatonin receptor- and extracellular signal-regulated kinase-mediated pathways. Fertil. Steril. 2011, 95, 2290–2296. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.Y.; Kim, Y.H.; Kim, B.W.; Park, I.C.; Cheong, H.T.; Kim, J.T.; Park, C.K.; Kong, H.S.; Lee, H.K.; Yang, B.K. Ameliorative effects of melatonin against hydrogen peroxide-induced oxidative stress on boar sperm characteristics and subsequent in vitro embryo development. Reprod. Domest. Anim. 2010, 45, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Wakatsuki, A.; Kaneda, C. Melatonin increases activities of glutathione peroxidase and superoxide dismutase in fetal rat brain. J. Pineal Res. 2000, 28, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Bhatla, S.C. Melatonin and nitric oxide modulate glutathione content and glutathione reductase activity in sunflower seedling cotyledons accompanying salt stress. Nitric Oxide 2016, 59, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Yang, X.G.; Lu, Y.Q.; Lu, S.S.; Zhang, M.; Yao, H.I.; Meng, L.J.; Lu, K.H. Protective effects of melatonin against oxidative stress in flow cytometry-sorted buffalo sperm. Reprod. Domest. Anim. 2012, 47, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Dehghani-Mohammadabadi, M.; Salehi, M.; Farifteh, F.; Nematollahi, S.; Arefian, E.; Hajjarizadeh, A.; Parivar, K.; Nourmohammadi, Z. Melatonin modulates the expression of BCL-xl and improve the development of vitrified embryos obtained by IVF in mice. J. Assist. Reprod. Genet. 2014, 31, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Korkmaz, A.; Paredes, S.D.; Manchester, L.C.; Tan, D.X. Melatonin reduces oxidative/nitrosative stress due to drugs, toxins, metals, and herbicides. Neuro Endocrinol. Lett. 2008, 29, 609–613. [Google Scholar] [PubMed]
- Ersoz, N.; Guven, A.; Cayci, T.; Uysal, B.; Turk, E.; Oztas, E.; Akgul, E.O.; Korkmaz, A.; Cetiner, S. Comparison of the efficacy of melatonin and 1400W on renal ischemia/reperfusion injury: A role for inhibiting iNOS. Ren. Fail. 2009, 31, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.A.; Ortiz, F.; Miana, J.; Doerrier, C.; Fernandez-Ortiz, M.; Rusanova, I.; Escames, G.; Garcia, J.J.; Acuna-Castroviejo, D. Contribution of inducible and neuronal nitric oxide synthases to mitochondrial damage and melatonin rescue in LPS-treated mice. J. Physiol. Biochem. 2017, 73, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Tapias, V.; Escames, G.; Lopez, L.C.; Lopez, A.; Camacho, E.; Carrion, M.D.; Entrena, A.; Gallo, M.A.; Espinosa, A.; Acuna-Castroviejo, D. Melatonin and its brain metabolite N(1)-acetyl-5-methoxykynuramine prevent mitochondrial nitric oxide synthase induction in parkinsonian mice. J. Neurosci. Res. 2009, 87, 3002–3010. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Kawamoto, M.; Sato, S.; Tamura, I.; Maekawa, R.; Taketani, T.; Aasada, H.; Takaki, E.; Nakai, A.; Reiter, R.J.; et al. Long-term melatonin treatment delays ovarian aging. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef] [PubMed]
- Scala, G.; Maruccio, L. Nitric oxide (NO) expression during annual reproductive activity in buffalo epididymis: A histochemical and immunocytochemical study. Theriogenology 2012, 78, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Rexhaj, E.; Pireva, A.; Paoloni-Giacobino, A.; Allemann, Y.; Cerny, D.; Dessen, P.; Sartori, C.; Scherrer, U.; Rimoldi, S.F. Prevention of vascular dysfunction and arterial hypertension in mice generated by assisted reproductive technologies by addition of melatonin to culture media. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1151–H1156. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Specie | Tissue | Treatment | Results | Reference |
|---|---|---|---|---|
| Human | Patient | 3 mg per day from the fifth day onwards of one cycle in women with diminished ovarian reserve | Increases the mean number of M-II oocytes, top-quality embryos with grade 1 and 2 | [77] |
| Human | Blastocyst | 10−7 M in culture system in 3D (Encapsulation) | Increases the survival time of encapsulated embryos | [78] |
| Human | Patient | 3 mg for 14 days in patients with polycystic ovarian syndrome | Enhances the oocyte and embryo quality | [79] |
| Human | Sperm | 0.01 mM in freezing extender before cryopreservation of sperm from infertile men | Increases motility and viability, decreases ROS and MDA levels | [80] |
| Human | Patient | 6 mg for 45 days | Increases the antioxidant capacity in seminal plasma, reduces the oxidative damage caused in sperm DNA, increases the quality of embryos | [81] |
| Human | Patient | 3 mg/day for 2 weeks | Increases the fertilization rate I the second cycle, improves the fertilization and embryos quality rate | [82] |
| Specie | Tissue | Treatment | Results | Reference |
|---|---|---|---|---|
| Porcine | Oocytes | 10−9 M during in vitro maturation | Increases cleavage and blastocyst rate and the total cell number of blastocyst; promotes lipid metabolis, providing energy for oocyte maturation and embryo development | [83] |
| Rat | Animal | Intraperitoneal injection of 20 mg/kg for 4 weeks | Increases testosterone hormone in blood serum and body weigh | [84] |
| Mouse | Spermatogonial stem cells | 10 mg/kg for 2 weeks after busulfan treatment | Relieves the loss and apoptosis in mouse testes; upregulates MnSOD | [85] |
| Mouse | Oocytes | 10−9 to 10−3 M after in vitro maturation | Increases in vitro fertilization rate, reduces ROS and inhibits apoptosis | [86] |
| Bovine | Zygotes | 1 µM for 3 h after insemination and at 40 °C | Reduces ROS levels in embryos | [87] |
| Mouse | Oocyte M-II | 10−9 mol/L during vitrification/warming and PA | Increases blastocyst rate after warming compared with control group | [88] |
| Bovine | Sperm | 10−3 M for 3 h before in vitro fertilization | Improves plasma membrane and acrosome integrity, mitochondrial activity; decreases intracellular ROS levels; increases the blastocyst rate and it decreases apoptosis rate | [89] |
| Bovine | Oocytes | 10−6 or 10−9 M for 24 h during in vitro maturation | Up-regulates MnSOD and Cu-ZnSOD in cumulus cells; decreases fragmentation. Decreases ROS levels In oocytes | [90] |
| Bovine | Embryos | 10−7 M melatonin for 24 h prior to exposure to 250 µM Paraquat (herbicide) | Decreases the incidence of apoptotic nuclei induced by Paraquat | [91] |
| Porcine | Oocytes | 0.1 μM for 22–44 h after endoplasmic reticulum stress during in vitro maturation | Improves oocyte maturation and cumulus cells expansion induced by endoplasmic reticulum stress | [92] |
| Bovine | Oocytes | Melatonin-loaded lipid-core nanocapsules at 10−6 M, 10−9 M and 10−12 M during in vitro maturation | Enhances in vitro embryo production, decreases ROS levels and the apoptotic nuclei, upregulates GPX1 and SOD2 and downregulates CASP3 and BAX | [93] |
| Bovine | Zygotes | Melatonin-loaded lipid-core nanocapsules at 10−9 M during in vitro culture | Increases hatching rate and embryo cell number, decreases cell apoptosis and ROS levels; downregulates BAX, CASP3, and SHC1 genes, and upregulates CAT and SOD2 genes | [94] |
| Mouse | Oocytes | 10−7 M during in vitro maturation | Improves blastocyst rate and cell number of blastocysts | [95] |
| Mouse | Sperm | 10 mg/kg body weight for 7 days during cadmium exposure | Reduces oxidative stress and inflammation induced by cadmium in male reproductive system | [96] |
| Mouse | Sperm | 0.125 mg/mL in freezing extender during cryopreservation | Increases progressive motility, decreases ROS levels and upregulates BCL-XL | [97] |
| Buffalo | Oocytes | 250 µM during in vitro maturation | Improves fertilization rate | [98] |
| Bovine | Oocytes | 1 µM during in vitro maturation of aged oocytes | Decreased aberrant spindle organization, increases ATP production, increases the development of bovine oocytes and reduces apoptotic rat; downregulates BAX and CASP3 and increases BCL2 | [99] |
| Rabbit | Morula | 10−3 M prior in vitro culture, prior vitrification | Promotes blastocyst rate, increases SOD activity and decreases LPO and NO levels | [100] |
| Mouse | Preantral follicles | 10 pM after vitrification, during culture | Increases diameter of follicles and their survival | [101] |
| Bovine | Embryos produced by SCNT | 10−11 to 10−2 M during in vitro culture | Increases total cell number, ICM and the development of bovine SCNT embryos; suppresses the expression of p53 and Bax, and upregulates SOD1, Gpx4, BCL2L1 and SOX2 | [102] |
| Porcine | Oocyte | 10−7 M during in vitro maturation under heat stress | Improves polar body and blastocyst rate impaired by heat stress.; preserves normal levels of steroid hormone, reduces ROS, enhances GSH production and inhibits apoptosis | [103] |
| Porcine | Oocyte and embryos | 25 ng/mL during in vitro maturation and culture | Increases blastocyst rate and decrease apoptotic nuclei in embryos | [104] |
| Bovine | Sperm | 1000 nM | Increases higher wobbler coefficient, decreases sperm with intact acrosome and viable spermatozoa with ROS | [105] |
| Rabbit | Embryos | 10−9 to 10−3 M during in vitro culture | Increases in vitro development and improves hatching rate | [106] |
| Bovine | Zygotes | 10−7 M during in vitro culture | Promotes the cleavage and blastocyst rate, accelerates the development of in vitro embryos and improves the quality of blastocysts | [107] |
| Bovine | Zygotes | 10−7 M for 2 days at the beginning of in vitro culture | Increases the blastocysts and hatched blastocyst rate | [108] |
| Bovine | Zygotes | 10−9 M for after 2 days of pre-culture and for the remaining 6 days of culture | Increases the blastocysts and hatched blastocysts rate | [108] |
| Bovine | GV oocytes | 10−9 or 10−7 M during in vitro maturation | Improves embryo development and the total cell number after in vitro fertilization; upregulates genes associated during in vitro maturation: GDF9, MARF1 and DNMT1α | [109] |
| Mouse | 2-cell embryos | 10 µM during in vitro culture | Improves quality and developmental rate of embryos; can prevent cell death | [110] |
| Rat | Sperm | 10 mg/kg weekly for 8 weeks | Improves sperm motility | [111] |
| Mouse | Embryos | 10−12 M during in vitro culture of embryos produced by SCNT | Increases embryo development | [112] |
| Ovine | Blastocysts | 10−9 M during thawing after cryopreservation | Improves embryo development after post warming culture | [113] |
| Deer | Animal | Subcutaneous implantation of 40 mg | Elevates serum FSH and LH levels, increases number of corpora luteal and the number of embryos | [114] |
| Sheep | Animal | Subcutaneous implantation of 40 or 80 mg | Increases corpus lutea, the number of recovered embryos, pregnancy and birth rates, and the number of lambs born per embryo | [115] |
| Porcine | Donor cell and embryos | 10−10 M in the medium for donor cell and 10−9 M during in vitro culture of embryos produced by SCNT | Increases proliferation of fetal fibroblasts and the blastocysts rate,; reduces the apoptotic nuclei.; upregulates BCL2L1 and downregulates BAX and p53 | [116] |
| Mouse | Oocytes | 10 to 100 nM during in vitro maturation | Increases expansion, maturation, fertilization and blastocyst rate in a dose dependent manner | [117] |
| Bovine | Oocytes | 10−12 to 10−3 M during in vitro maturation under heat stress | Increases blastocyst rate of embryos submitted to heat stress | [118] |
| Murine | Pronuclear embryos | 10−7 M during in vitro culture | Promotes embryo development, blastocyst rate, hatching rate and blastocyst cell number; upregulates SOD and BCL2 and downregulates CAS3 and p53 | [119] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loren, P.; Sánchez, R.; Arias, M.-E.; Felmer, R.; Risopatrón, J.; Cheuquemán, C. Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals. Int. J. Mol. Sci. 2017, 18, 1119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061119
Loren P, Sánchez R, Arias M-E, Felmer R, Risopatrón J, Cheuquemán C. Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals. International Journal of Molecular Sciences. 2017; 18(6):1119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061119
Chicago/Turabian StyleLoren, Pía, Raúl Sánchez, María-Elena Arias, Ricardo Felmer, Jennie Risopatrón, and Carolina Cheuquemán. 2017. "Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals" International Journal of Molecular Sciences 18, no. 6: 1119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061119